4.1.3 Скелет парных конечностей и их поясов
Ногообразные конечности наземных позвоночных произошли из парных плавникообразных конечностей предковых форм, живших в водной среде. В примитиве за исходную форму конечностей берется кожно-мышечная складка, которая проходила вдоль тела животного и выполняла роль рулей глубины и сохранения определенного положения тела в водном пространстве. Впоследствии средняя часть боковой складки редуцируется и образуются парные грудные и брюшные плавники, распологающиеся в менее подвижных участках осевого отдела туловища. Плавникообразные конечности, присущие всем рыбам, сохраняют первоначальную основную функцию складок, но приобретают большие возможности для адаптивных изменений, связанных как с совершенствованием локомоторной функции, так и с выполнением каких-либо дополнительных приспособлений.
У хрящевых рыб плавники располагаются в горизонтальной плоскости, выполняют роль рулей глубины и незначительную опорную функцию. Увеличение размеров плавников приводит к усилению в них хрящевой основы, которая первоначально представлена метамерно расположенными лучами. Хрящевые лучи своими основаниями, образуют общую хрящевую пластинку, которая затем преобразуется в скелет пояса конечности (рисунок 24).
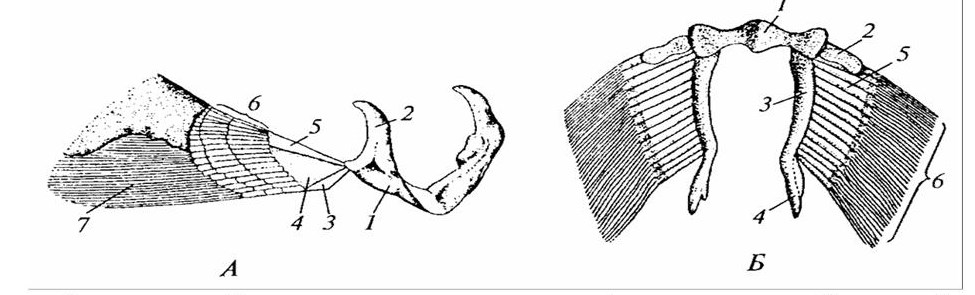
А – плечевой пояс и грудные плавники: 1 – коракоидный отдел; 2 – лопаточный отдел; 3, 4, 5 – базалии; 6 – радиалий; 7 – эластоидиновые нити. Б – пояс задних конечностей и брюшные плавники: 1 – пояс; 2, 3, 4 – базалии; 5 – радиалий; 6 – эластоидиновые нити.
Рисунок 24 – Скелет поясов и парных конечностей самца акулы
У костистых рыб, с их более разнообразными приспособлениями к условиям существования, плавники стали выполнять функцию изменения направления движения, роль органов опоры (особенно у двоякодышащих), и даже в осуществлении планирующего полета (летающие рыбы). Такие адаптации сопровождались усилением хрящевого пояса, охватывающего тела со всех сторон, и образованием на его боковой поверхности выступа для соединения с главным, или осевым, лучом плавника. Плавник по отношению к осевому отделу туловища стал занимать не горизонтальное, а наклонное или даже поперечное положение, в результате чего бывшая дорсальная поверхность становится краниолатеральной. Усиление функциональной нагрузки на плавникообразные конечности привело к дополнительному усилению их хрящевого остова костными образованиями кожного происхождения, которые затем постепенно замещают хрящевой скелет. Одновременно происходит и усложнение костной основы плавника, где наряду с расчленением конечностей на звенья происходит и их дифференциация. Брюшные плавники, имеющие меньшую функциональную загруженность, длительное время сохраняют и более примитивное строение (рисунок 25).
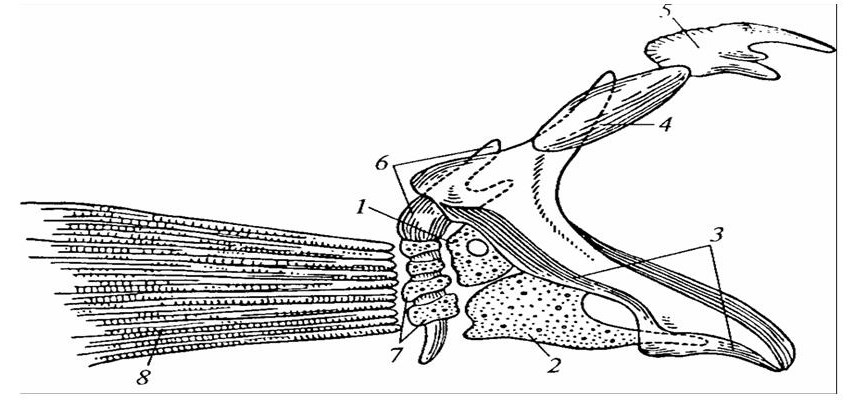
Первичный пояс: 1 – лопатка; 2 – коракоид. Вторичный пояс: 3 – клейтрум; 4 – супраклейтрум; 5 – заднетеменная кость; 6 – подклейтрум. Скелет плавника: 7 – радиалий; 8 – лепидотрихии (кожные костные лучи).
Рисунок 25 – Грудной пояс костистой рыбы судака
У наземных позвоночных за счет соединения вентральных концов нижниx ребер образуются парные хрящевые пластинки, которые, срастаясь между собой, образуют тело грудины. У амфибий в связи с вторичной редукцией нижних ребер грудина развивается независимо от ребер и имеет непосредственное отношение к плечевому поясу, так как служит местом прикрепления коракоидных костей. В связи со сменой среды обитания на воздушную плавникообразные конечности преобразуются в короткие ногообразные конечности с характерным подразделением их остова на скелет пояса и скелет свободного отдела, как и у последующих классов наземных позвоночных, подразделенного на три основных звена – плечо – бедро, предплечье – голень, кисть – стопа (рисунок 26). Несмотря на наличие коротких ногообразных конечностей, основной принцип движения для хвостатых амфибий, точно так же и для ящерищеобразных, сохраняется прежний – боковые изгибы осевого отдела туловища и хвоста.
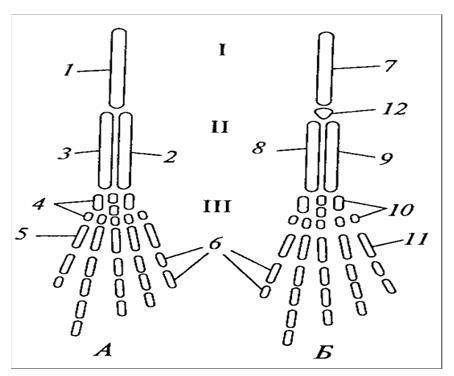
А– передняя конечность; Б – задняя ко ценность; I – плечо – бедро; II – предпечье – голень; III – кисть – стопа; 1 – плечевая кость; 2 – локтевая кость; 3 – лучевая кость; 4 – запястье; 5 – пясть; б – фаланги пальцев; 7 – бедренная кость; 8 – большая берцовая кость; 9– малая берцовая кость; 10 – предплюсна; 11 – плюсна 12 – коленная чашечка.
Рисунок 26 – Схема строения парных конечностей наземных позвоночных
Скелет плечевого пояса у низших наземных позвоночных сохраняет многие черты такового костистых рыб. Однако в связи с большим значением при опоре и передвижении животного его отделы приобретают большую дифферецированность и усиление за счет развития аналогичных костных образований кожного происхождения, которые в конечном итоге преобразуются в дорсальный – лопатку и вентральный отделы – коракоид и ключицу (рисунок 27). Наибольшее развитие приобретают коракоидиая кость, как опорное звено, и лопатка, обеспечивающая прочное соединение конечности с туловищем.
Тазовый пояс, как и плечевой, имеет те же отделы, из которых большее развитие получил вентральный, представленный; седалищной и лонной костями, в то время как подвздошная, относящаяся к дорсальному отделу развита значительно слабее (рисунок 27). Она имеет вытянутую заостренную форму, направлена дорсокаудально и соединяется с боковой поверхностью ребер и одним из позвонков.
У рептилий, в частности у ящериц и крокодилов, плечевой пояс включает в основном те же элементы, что и у амфибии (рисунок 28).
Брюшная его часть представлена коракоидами, несущими сочленовные ямки для причленения плечевых костей. Спинная часть пояса состоит из лопатки и надлопаточного хряща. Коракоид связан с грудиной и обладает подвижностью. Впереди от него расположена ключица. Характерно наличие крестообразного надгруднаника, лежащего перед грудиной и связывающего ее с ключицами. В целом плечевой пояс рептилий более прочен, чем у амфибий, и эта его черта отражает их лучшую приспособленность к передвижению по суше.
В тазовом поясе парные кости: подвздошная, седалищная и лобковая соединены в единый замкнутый таз. Спереди и сзади, на стыке костных элементов расположены передний и задний хрящевые отростки. Подвздошными костями пояс причленяется к поперечным отросткам двух крестцовых позвонков. На стыке трех костей таза расположена вертлужная впадина: в нее входит круглая головка бедренной кости.
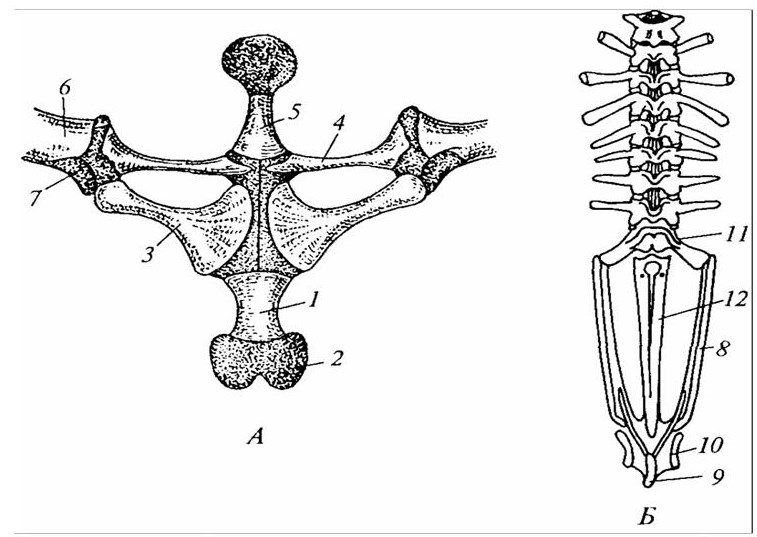
1 – грудина: 2 – хрящевая задняя часть грудины; 3 – коракоид; 4 – ключица лежит на прокоракоиде; 5 – предгрудина; 6 – лопатка; 7 – сочленовная впадина на лопатке для плеча (хрящ покрыт точками); 8 – подвздошная кость; 9 – седалищная кость; 10 – вертлужная впадина; 11 – крестцовый позвонок, 12 —уростиль.
Рисунок 27 – Плечевой (А) и тазовый (Б) пояс лягушки (вид спереди)
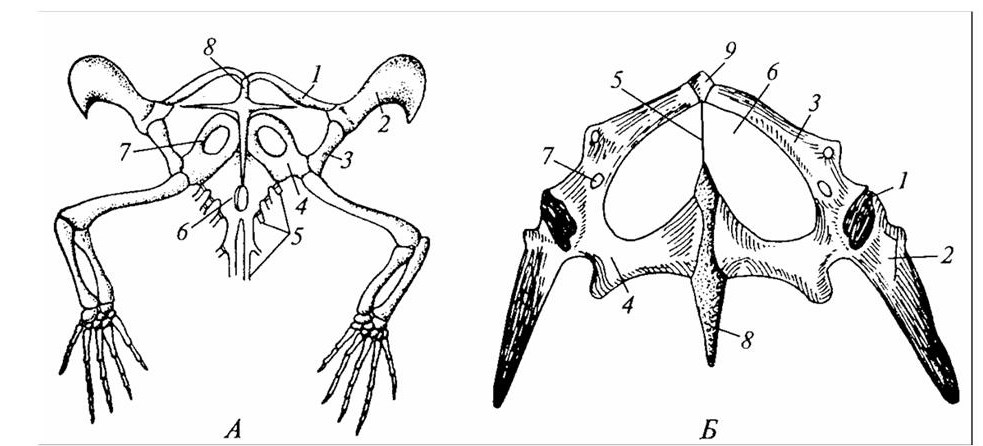
А: 1 – ключица; 2 – надлопаточный хрящ; 3 – лопатка; 4 – коракоид; 5 – ребра; 6 – грудина; 7 – переднекоракоидный хрящ; 8 – надгрудинник. Б: 1 – суставная впадина для головки бедра; 2 – подвздошная кость; 3 – лобковая кость; 4 – седалищная кость; 5 – связка; 6 – «окно»; 7 – запирательное отверстие; 8 – задний хрящевой отросток; 9 – передний хрящевой отросток.
Рисунок 28 – Плечевой (А) и тазовый (Б) пояс ящерицы
Передняя и задняя конечности имеют типичное строение. Особенность конструкции суставов в конечностях рептилий иная, чем у амфибий: сочленение в передней конечности лежит между костями запястья (межзапястный, или интеркарпальный, тип), а в заднем – между костями предплюсны (межпредплюсневый, или интертарзальный, тип). Конструкция сустава создает сгибательно-разгибательную подвижность конечности. Сходное строение имеют суставы птицы.
У птиц скелет передних конечностей (пояс и свободная конечность) имеет черты, преемственные от рептилий, но в связи с приспособлением к полету отличается рядом особенностей. Пояс включает лопатки, коракоиды (вороньи кости) и ключицы, которые своими проксимальными концами формируют площадки для причленения плечевых костей. Лопатка длинная, саблевидно изогнутая, лежит на ребрах, по которым она может свободно скользить. Коракоид развит очень хорошо. Одним концом он упирается в грудину, другой конец дает опору для причленения плеча. Грудина, являющаяся местом прикрепления мощной вентральной группы мышц плечевого пояса и выполняющая основную функциональную нагрузку при полете получает наиболее сильное развитие. Левая и правая ключицы срастаются между собой, образуют характерную для птиц вилочку, придающую поясу особую упругость.
Скелет крыла состоит из всех типичных для пятипалой конечности отделов (рисунок 29). При этом плечо и предплечье существенно не изменены, а кисть, наоборот, имеет ряд своеобразных особенностей. Сильно видоизменено запястье. Его проксимальные части срастаются в две косточки, а дистальные – с пястью. Пясть состоит из двух удлиненных костей, сращенных как в проксимальном, так и в дистальном отделах и образующих в итоге сложную пястно-запястную кость, или пряжку.
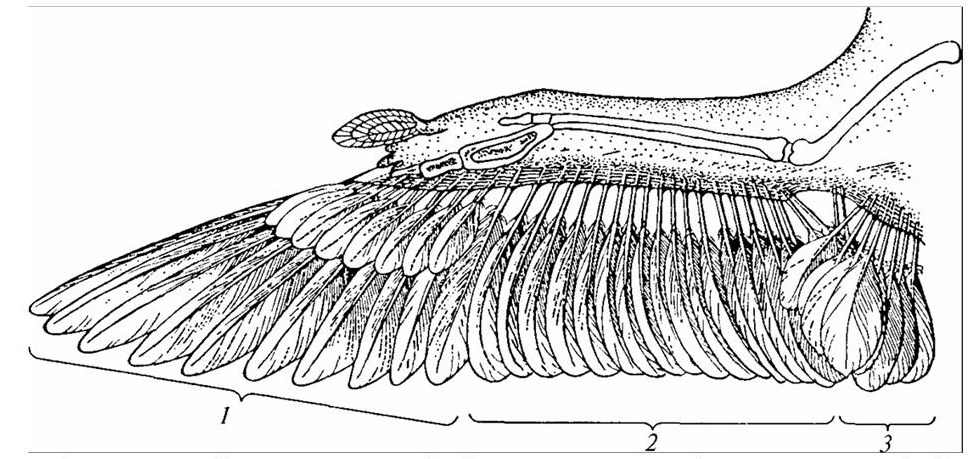
1 – кисть (первостепенные); 2 – предплечье (второстепенные); 3 – плечо (третьестепенные).
Рисунок 29 – Скелет крыла птицы и расположение маховых перьев
Из пальцев сохраняются только три – второй, третий и четвертый, при этом только третий палец имеет две фаланги, а второй и четвертый – по одной. В связи с наличием особых мускулов, сухожилий и кожистых перепонок кости крыла соединены между собой так, что все крыло не может быть вытянуто по прямой, и его части расположены под некоторым углом друг к другу. Отделы конечности могут двигаться только в одном направлении – в плоскости крыла, складывая и расправляя его.
Скелет задних конечностей (рисунок 30) (пояс и свободная конечность) птиц также имеет ряд особенностей, связанных с тем, что при движении по земле птица переносит на них всю тяжесть тела. Прочность осевого корпуса тела определяется срастанием больших подвздошных костей по всей их длине со сложным крестцом. С подвздошными костями срастаются также весьма крупные седалищные кости. Лобковые кости, наоборот, малы; в виде тонких палочек они присоединены к наружному краю костей седалищных. Все три кости таза участвуют в образовании вертлужной впадины. В отличие от других наземных позвоночных (рептилий, млекопитающих) у птиц отсутствует лобковый симфиз – сочленение тазовых костей внизу: две половины тазового пояса широко расставлены, что связано с откладыванием крупных яиц, покрытых твердой скорлупой, и необходимостью в широком тазовом выходном отверстии. Такой тип таза называют открытым. Закрытый таз имеют только африканские страусы.
Бедро типичного строения. Голень состоит из двух типичных костей, однако полностью развивается только большаяперцовая кость, малая берцовая кость рудиментарна и прирастает к первой. К дисталыюй части голени прирастает проксимальный ряд косточек предплюсны. Лежащий вслед за голенью отдел конечности носит название цевки. У взрослой тины он состоит из одной длинной кости, подвижный сустав у птиц (как и у пресмыкающихся) расположен между двумя рядами костей предплюсны, а не между предплюсной и голенью, как у амфибий. Это сочленение именуется межпредплюсневым, или интертарзальпым.
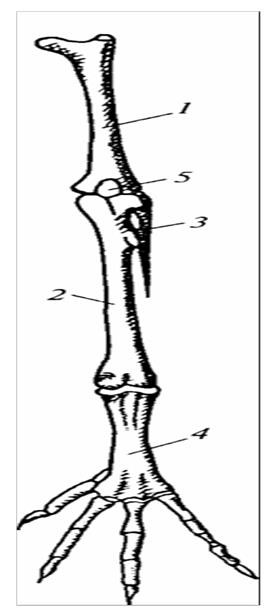
1 – бедро; 2 – большая берцовая кость; 3 – редуцироманная малая берцовая кость; 4 – цевка; 5 – чашечка.
Рисунок 30 – Скелет задней конечности голубя
Дальнейшие дифференциация и специализация грудных и тазовых конечностей наземных позвоночных стали возможными лишь после поворота конечностей из трансверзальной (поперечной) плоскости в боковую, отрыва тела животного от земли и подведения конечностей под туловище. Эти преобразования привели к усилению дорсальных отделов поясов и превращению конечностей в активные органы локомоции. На грудных конечностях в связи с усилением грудной кости и развитием лопатки произошла полная редукция коракоида, а с утратой элементов мультифункциональности – значительное ослабление или редукция ключицы. Редукция вентральных звеньев плечевого пояса привела к значительному усилению мышц, обеспечивающих укрепление тела животного междуконечностями в подвешенном состоянии.
Из-за того, что тазовые конечности стали выполнять роль основных движителей в осуществлении поступательного движения, в тазовое поясе произошли значительное усиление дорсального отдела, смещение его в дорзосокраниальное положение и прочное сращение с позвоночным столбом. Кости вентрального звена срослись между собой по вентральной срединной линии и приняли участие в образовании вентральной срединной полости, которая стала продолжением брюшной. Включение тазового пояса в состав осевого отдела привело к тому, что он стал основным передатчиком двигательных усилий, исходящих от тазовых конечностей. Свободные отделы грудных и тазовых конечностей, имеющих в примитиве аналогичные по строению и гомологичные по происхождению звенья, с перемещением конечностей из трансверзальной плоскости в парамедианную испытали характерные изменения в их взаимоотношения.
На грудных конечностях при подведении их под туловище при незначительной изменении положения костей кисти поворот отдельных звеньев осуществлялся в плечевом и локтевом суставах, что привело к Хобразному перекресту костей предплечья. Каудальная поверхность плеча стала медиальной, а вершина локтевого сустава, имевшая дорсолатеральное направление, стала располагаться с каудальной стороны конечности. На тазовых конечностях, в отличие от грудных, поворот осуществлялся всем свободным отделом так, что бывшая латеральная поверхность стала краниальной, а краниальная – медиальной. Кости стопы, имевшие латерокаудальное направление пальцев, стали располагаться краниально. В связи с одновременным перемещением всех звеньев конечностей Х-образного перекреста костей голени не произошло, и поэтому у всех видов животных малая берцовая кость на голени занимает латеральное положение. Дальнейшая специализация аппарата движения происходила в зависимости от характера выполняемых функций и типа опоры животного.
У животных с развитием и совершенствованием скорости передвижения в процессе эволюции происходит смена опоры от стопо- через пальце- к фалангохождению (рисунок 31).
При смене опоры изменения в строении конечностей касаются не только редукции боковых пальцев, как это имеет место у парнокопытных (жвачные, нежвачные, мозоленогие), и непарнокопытных (тапиры, носороги и лошади), но и всех выше расположенных звеньев, которые выразились в изменении их длины, толщины, рельефа суставных поверхностей, степени развития связок, соответствующих мышц сосудов и нервов.
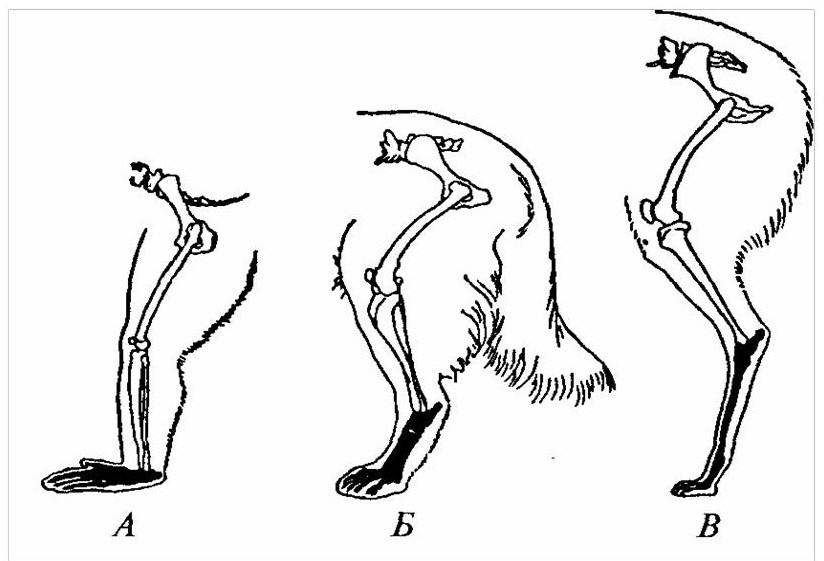
А – обезьяна павиан; Б – собака; В – лама.
Рисунок 31 – Задние конечности пальце- и стопоходящих млекопитающих. Элементы стопы затемнены
Знание коррелятивных изменений в строении периферического скелета при различных адаптациях позволяет при анализе палеонтологических материалов вскрыть основные закономерности эволюции конкретных видов животных, как это было сделано В.О. Ковалевским в отношении современной лошади. Он убедительно доказал, что одним из древнейших предков современной лошади был фенакод, который жил в начале третичного периода (около 50 млн. лет назад) в восточной части Северной Америки, где условия того времени характеризовались теплым влажным климатом и мягкими почвами с богатой сочной растительностью. Фенакод имел небольшие размеры (68 см в высоту) и пятипалую конечность. Из пяти пальцев два боковых были самыми короткими, а из трех других средний имел наибольшую длину с роговым башмачком на конечной фаланге, что свидетельствует о ведущей роли при передвижении.
Таким образом, внутренний скелет хордовых энто- и мезодермального происхождения поддерживает тело, служит для укрепления скелетных мышц, участвует в движении, обеспечивает защиту внутренних органов. Прогрессивные изменения связаны с заменой в осевом скелете единого нечлененного упругого тяжа – хорды сначала хрящевым, а затем костным позвоночником, его последовательной дифференцировкой на отделы и появлением мозгового черепа. В висцеральном скелете в эволюционном ряду позвоночных висцеральные дуги редуцируются, развиваются челюсти, формируются слуховые косточки. Мозговой и висцеральный скелет объединяются в единое морфофункциональное образование – череп с лицевой и мозговой частями. Скелет органов движения у первичноводных хордовых – первоначально непарных плавников – прогрессивно развивается в связи со специализацией к условиям обитаний, затем дополняется парными плавниками, используемыми как стабилизаторы и рули при движении и поворотах. Появление мускулатуры на унисериальных парных плавниках, использовавшихся в качестве опоры при движении по грунту дна, послужило основой для формирования пятипалой конечности наземного типа.
Данный текст является ознакомительным фрагментом.