Глава седьмая. Новый ключ к старым тайнам
Глава седьмая. Новый ключ к старым тайнам
Таким образом, голограмма, которая
вначале была использована как метафора или
аналогия для объяснения некоторых сторон
нарушения деятельности нервной
системы, стала точной моделью
нормальных форм ее работы.
Карл Прибрам. Языки мозга
Многие думают, что память – это нечто вроде запасника картинной галереи: стоят у стенки прислоненные друг к другу тысячи полотен, нужно вспомнить – вытащил, посмотрел...
– Недавно перечитывал Антокольского, – сказал Глезер, – и запомнились строчки:
Что память?.. Кладовая. Подземелье.
Жизнь как попало сброшена туда.
Спят на приколе мертвые суда.
Недвижные, не сдвинутые с мели...
Красивая картина. Очень впечатляющая. В поэзии, конечно, можно все, на то она и поэзия. А в жизни...
Кто посмотрит на эти «суда»? Древние отвечали: душа. Но мы-то знаем, что никакой отдельно живущей от тела души нет. Нет в мозгу у человека маленького человека, который смотрит этакий телевизор: чего, мол, там видит своими глазами человек, какие образы складывает в памяти? Десять, а по другим данным даже 50 миллиардов нервных клеток у нас в мозгу, идут от одной к другой электрические импульсы разной частоты и амплитуды, в межклеточном пространстве и в клетках происходят химические изменения, и кроме этого ничего – понимаете, ничего! – нет. А мы видим, и память существует, и картины прошлого мы с вами вспоминаем. Что же приходит из глаза в мозг?
В средние века считалось, что приходят идеи. Поступают по зрительным нервам и складываются в резервуаре памяти, который полагали находящимся где-то возле затылка. Но опять-таки слово «идеи» ничего не объясняло.
Когда широко распространилось книгопечатание и листы с гравюрами на божественные и светские темы получили повсеместное хождение, стали учить, что в мозгу каким-то путем возникают «отпечатки» изображений, переносятся туда, мол, картинки того, что хрусталик проецирует на заднюю стенку глазного яблока. Подобные гипотезы укрепились особенно в конце XIX в., раскрывшего строение глаза и роль сетчатки с ее светочувствительными палочками и колбочками.
Большой популярностью пользовалось мнение, что от каждого светоощущающего рецептора идет в мозг одно нервное волокно, формируя в коре «рельеф возбуждения» – этакую фотографию увиденного. Долгое время гипотеза представлялась единственно верной, ее защищали крупнейшие физиологи, в частности Иван Михайлович Сеченов. Но все-таки пришлось от нее, несмотря на заманчивую простоту и наглядность, отказаться, когда выяснилось, что чувствительных элементов сетчатки раз в полтораста больше, нежели волокон зрительного нерва: как при таких условиях может сформироваться картинка? (Заблуждения чудовищно цепки и живучи. Даже в середине ХХ в. всерьез защищались такие взгляды: зрительные ощущения суть фотографические копии того, что представляется взору. В солидных книгах писали...)
Проблема человечка-гомункулуса в мозгу явно бесплодна. Что же противопоставляет ему современная наука? В книге «Информационные процессы мозга и психическая деятельность» высказывается мысль, что свойственное человеку «Я» – это то содержимое памяти, которое извлекается именно в тот момент, когда в мозг приходит сигнал от органов чувств: нельзя ощутить внешний сигнал без своего «Я» и нельзя почувствовать своего «Я» без внешнего сигнала. Поэтому бессмысленно искать человеческое «Я» в мозгу, как ищут золотые самородки. Ощутить себя можно лишь во взаимодействии с внешним миром, получая от него какие-то сигналы. Человек располагает своим «Я» только в виде информационной системы, в столкновении памяти с сиюминутным восприятием (у очень многих людей самопортрет, то есть мысленное представление о своей внешности, существенно расходится с тем, что показывает объективное зеркало: «Ах ты, мерзкое стекло, это врешь ты мне назло!»). Но как бы то ни было, вопрос о том, в каком виде картины внешнего мира приходят в мозг и получают там вид образов, не снимается словами об «информативном столкновении».
Казалось, в тоннеле забрезжил свет, когда в 1959 г. физиологи Дэвид Хьюбел и Торстен Визел, работавшие в Гарвардском медицинском институте, ввели в затылочную кору кошки (туда, где оканчиваются волокна зрительного нерва) микроэлектрод и обнаружили нейроны, к которым сходились сигналы уже не от нескольких сотен фоторецепторов, как к ганглиозным клеткам сетчатки, а сразу от многих тысяч. Это выдающееся открытие было следствием новой техники эксперимента. Раньше, чтобы обнаружить поле, связанное с ганглиозной клеткой, требовался простой сигнал: тонкий, словно спица, луч. Яркая точка на экране – вот что возбуждает «он»- и «офф»-поля сетчатки. Клеткам коры нужны иные стимулы для возбуждения – прямые линии и прямоугольники.
Однако не всякий стимул заставит заговорить клетку. «Нередко требуются многочасовые поиски, чтобы обнаружить отдел сетчатки, связанный с определенной клеткой коры, и подобрать оптимальные для этой клетки раздражители», – писал Хьюбел. По виду стимула и разделили американские исследователи обнаруженные ими поля клеток зрительной коры (поля коры, чтобы каждый раз не говорить «клеток»).
Простые поля выделяют только прямые тонкие линии. Едва линия попадает в область сетчатки, где дислоцировано поле, как нейрон коры буквально кричит: вижу, вижу! Убрали линию в сторону – замолкла и клетка, словно погасла сигнальная лампочка.
Сложные настроены на перепады яркостей типа «прямой край», «угол», «дуга». Они срабатывают и тогда, когда в поле зрения появляется движущийся предмет, – в чем-то сродни лягушачьим детекторам. Однако то, что клетки-сигнализаторы находятся не в сетчатке, а в коре мозга, говорит нам о куда большей сложности и гибкости зрительного аппарата млекопитающих.
Все эти поля ощущают ориентацию, – нужно тридцать таких полей, чтобы выделить наклоны одной-единственной линии через каждые шесть градусов во всем диапазоне углов от нуля до 180 градусов.
Есть поля, которые видят, скажем, только горизонтальную линию, движущуюся сверху вниз, а на вертикальную, гуляющую вправо-влево, внимания не обращают.
Сверхсложные поля выделяют уже не просто линии, а линии вполне определенной длины. Небольшое отклонение размера в ту или иную сторону, и реакцию нейрона не обнаружишь, «лампочка» не вспыхивает.
А то вдруг микроэлектрод натыкается на клетку, которой природа задала задачу: реагировать только на информацию, поступающую сразу от обоих глаз, и молчать, если один из них не видит стимула на экране. Сдвинули контакт чуть глубже или в сторону (не забывайте, что толщина коры головного мозга у кошек равна максимум двум миллиметрам, а у человека – четырем с половиной) – здесь нейрон, воспринимающий сигналы преимущественно от правого глаза, а рядом – в основном от левого: они имеют прямое отношение к проблеме объемного, бинокулярного зрения, которой мы еще займемся.
Полей коры – тысячи, сотни тысяч и миллионы. Перекрывая друг друга, именно они позволяют зрительному аппарату оценивать с помощью одних и тех же рецепторов сетчатки и элементы контура, и яркость, и цвет, и многое другое, делать это сразу по всему зрительному пространству, открывающемуся глазу, одновременно.
В области наиболее четкого зрения – центральной ямке сетчатки – находятся маленькие поля, позволяющие тонко распознавать форму предметов.
Ближе к краям сетчатки – поля крупные, с помощью которых форму не различишь, но зато уловишь яркость и движение, так что даже боковым зрением удается заметить мчащийся автомобиль или вспыхнувший фонарик (эти специфические поля обнаружены у всех млекопитающих, с какими только работали по этой теме ученые). Стоит появиться на краю поля зрения чему-то движущемуся, как через 15...17 сотых долей секунды туда непроизвольно обращается взор. Причем исключительно точно: пауза, потом быстрый скачок (скорость плавно нарастает до максимальной и так же плавно сбрасывается до нуля), и центральная ямка направлена прямехонько на объект, чтобы дальше безошибочно отслеживать его движение. Что все это значит?
Только то, что данные о движении предмета – его скорости, направлении, ускорении – зрительный аппарат вырабатывает перед началом ясного видения, по каналу (или каналам), с опознанием формы никак не связанным.
И действительно, нейроанатомы нашли, что подкорковая структура «ядро глазодвигательного нерва» получает сигналы прямо от сетчатки, минуя НКТ, а также от затылочной коры, такое двойное подчинение, по-видимому, и обеспечивает перемещение глазного яблока задолго (по скоростям мозговых процессов) перед тем, как мы увидим ясно то, на что обратили взор.
Поля – врожденные структуры. Их отыскали даже в коре еще не прозревших котят. «Существовавшее ранее представление о том, что большинство связей мозга функционально отбирается из совокупности случайно сформированных соединений, в настоящее время кажется неправомочным. Уже на ранних стадиях развития большинство связей устанавливается точно; существует много доказательств того, что образованные связи специфичны не только для данной области мозга, но и для данного нейрона (а в некоторых случаях и для данных частей нейрона) внутри этой области», – читаем мы в книге «Мозг».
И возникла мысль, что поля, реагирующие на линии, углы, дуги, площадки и так далее, – не что иное, как те самые детекторы, задача которых состоит в выделении признаков разных изображений. А затем, мол, эти признаки объединяются между собой в высших отделах мозга, формируют сложные признаки, потом еще более сложные, и в конце концов находится где-то гностический (от греческого «гнозис» – знание, познание) нейрон, сигнализирующий, что в поле зрения появился данный предмет. По этому поводу шли дискуссии, и Джордж Сомьен писал в книге «Кодирование сенсорной информации в нервной системе млекопитающих», что «вопрос о возможности существования клеток «детекторов кошки», т.е. нейронов или групп нейронов, специализированных для идентификации определенных классов предметов, вновь поднят со всей серьезностью».
Увы, дальнейшие исследования показали, что детекторная гипотеза «не проходит». Ну хотя бы потому, что не отвечает на такой вопрос: почему мы одинаково хорошо способны узнать льва и на реалистическом, во всех подробностях, рисунке, и в изощренно-стилизованном или наивно-детском изображении? Фрагменты каждый раз разные, сработают разные выделяющие детекторы, а результат один и тот же. Что же, для каждого такого льва существует отдельный детектор? Сомнительно, покачал бы головой философ Уильям Оккам (ок. 1285 – 1349), провозгласивший принцип: не нужно делать посредством большего то, что можно достичь посредством меньшего...
Где же выход? Оказалось, в новом подходе к роли полей, в той гипотезе, которая была выдвинута коллективом Лаборатории профессора Глезера.
...Кошке сделали трепанацию черепа, просверлили дырочку в черепной коробке. Кошки переносят операцию завидно хорошо, к вечеру уже прыгают.
Но эта лежит неподвижно. Она кураризирована: в вену ей мелкими каплями подают кураре, тот некогда таинственный яд, которым южноамериканские аборигены смазывали наконечники своих копий и стрел. Кураре, словно выключатель, останавливает действие мышц, и глаза кошки направлены строго в одну точку, туда, где ей на экране показывают «кино». Тихо шуршит аппарат искусственного дыхания. Кошка лежит на теплой грелке и, не исключено, блаженствует. Во всяком случае не сердится и не искажает своей злостью результатов опыта.
А по экрану проплывает светлая полоска, ведь неподвижные глаза иначе ничего не увидят. Вот полоску сменила «зебра» – две светлые полоски с темным промежутком между ними, а то по команде экспериментатора появятся «зебра» из трех полосок, четырех, пяти... Решетки... Пространственные частоты, каждая из которых – речь, обращенная к мозгу...
– Они открыли нам, что мозг действительно занимается голографией, – сказал Глезер. И стал рассказывать прежде всего не о своей лаборатории, а о работах Хьюбела и Визела.
Эти нейрофизиологи в конце 60-х годов выяснили, что в затылочной коре кошки можно обнаружить не одну клетку, настроенную на выделение линии определенного размера, а несколько. Требовалось только двигать микроэлектрод строго перпендикулярно к поверхности коры, и такие клетки встречались одна за другой, словно лежащие столбиком монетки. А рядом другой столбик, настроенный на такую же линию, только иного наклона...
В итоге на площади около 0,8 x 0,8 миллиметра – в модуле – собираются столбики нейронов, отхватывающих все ориентации линии, от нуля до 180 градусов (см. иллюстрацию справа). Замечательно, что у человека и обезьян в каждом столбике примерно 260 клеток, и это число удивительно стабильно по всей затылочной коре. А у всех иных млекопитающих – там только по 110. И хотя серьезные доказательства отсутствуют, есть мнения, что такое различие имеет далеко идущие следствия. Вполне возможно, что именно оно определяет более высокие интеллектуальные способности приматов. Ведь во всех других, не-зрительных участках коры, даже у человека столбики состоят из 110 нейронов...
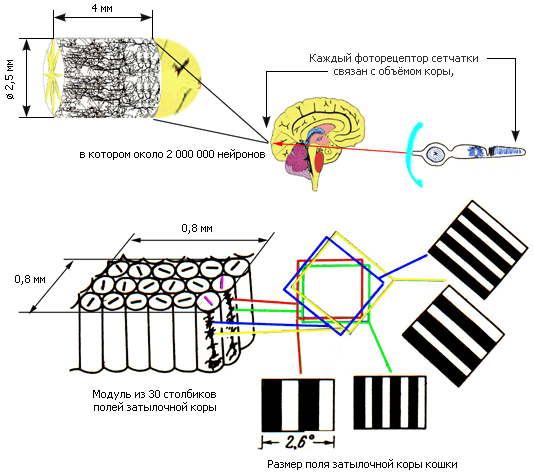
Рис. 45. Модульная организация коры головного мозга. Столбики модуля распознают линии и решетки всех ориентаций от 0 до 180 градусов через каждые 6 градусов. Размер поля затылочной коры кошки определяет остроту ее зрения. У человека поле впятеро меньше – 0,5 градуса – и соответственно острота зрения выше
Английский нейрофизиолог Вернон Маунткасл, настойчиво пропагандирующий мысль о модульной организации коры, открыл это свойство мозга в конце 50-х годов прошлого века. Он показал, что вертикальные связи нейронов каждого столбика гораздо интенсивнее, чем горизонтальные, передающие сигналы от столбика к столбику. И что каждый столбик работает относительно независимо, делает свое дело как суверенная система с собственными входом и выходом. Маунткасл исследовал соматосенсорную кору, область мозга, на которую проецируются окончания нервов, несущих сведения от кожи и внутренностей. Эксперименты Хьюбела и Визела подтвердили, что таким же способом устроена зрительная кора.
Хитрость только в том, что каждый фоторецептор сетчатки соединен не с одним нейроном, а сразу со многими тысячами. «Линия связи» от светочувствительной клетки захватывает в коре целый цилиндр диаметром примерно два с половиной миллиметра. А ведь на каждом квадратном миллиметре коры вглубь уходит около 100 тысяч нейронов!
Гипотеза XIX в. о прямых связях «фоторецептор – нейрон коры», как мы знаем, не подтвердилась. Но некое зерно ее все же оказалось жизнеспособным. Выяснилось, что соединения между сетчаткой и зрительной корой, несмотря на промежуточные преобразования, упорядочены топографически (припоминаете гипотезу Бернштейна?). Иными словами, если по сетчатке начнет блуждать яркая звездочка, максимум возбуждения нейронов затылочной коры – локус – повторит все ее эволюции. Пойдет точка вправо, и в соответствующую сторону двинется локус, точка вверх – и локус в то место коры, которое соответствует верхней части сетчатки.
Восхитительная сложность организации зрительной системы на этом уровне еще не кончилась. Ведь что означает разветвляющаяся линия связи «фоторецептор – цилиндр коры»? Только то, что мозаика фоторецепторов, этих вполне отделенных друг от друга образований (дискретных, сказал бы специалист), представлена в коре колоссальным множеством перекрывающихся нейронных цилиндров. То есть, по сути, непрерывно. Дискретная топография преобразовалась в «гладкую» – вот разгадка того изумлявшего ученых парадокса, что мы видим линии сплошными, хотя они воспринимаются с помощью дискретных элементов сетчатки, то есть палочек и колбочек.
Наконец, модули, входящие в цилиндры коры, правильно чередуются: один связан с правым глазом, соседний с левым, и так далее... Четкость, предопределенная генетически, видна повсюду в нервной системе. «Хотя, вообще говоря, схема соединений мозга очень запутана, работы последнего времени показали, что эти соединения гораздо более упорядочены, чем можно было думать», – отметил Френсис Крик. Действительно, у одного из червей нейронная сеть состоит всегда из 279 клеток, не больше и не меньше, и каждая соединена с другими клетками одинаково точно и занята только ей присущим делом. Вам кажется некорректным сравнение человека с червем?
Но вот что говорит Хьюбел: «Принципы нейронной функции удивительно сходны у столь далеких друг от друга животных, как улитка и человек; большая часть того, что известно о нервном импульсе, изучено на кальмаре. Даже основные структуры головного мозга так сходны, например, у кошки и человека, что нередко не имеет значения, чей мозг изучать».
То, что зрительная кора и НКТ организованы в топографическом плане соответственно сетчатке, то есть ретинотопически, объясняет немало зрительных феноменов. Ведь это наиболее простой и эффективный способ выделения простейших пространственных признаков любой картины и ее деталей: «справа», «слева», «сверху», «снизу», «большой», «маленький», «подвижный», «неподвижный» и так далее. Конечно, такого описания мира еще недостаточно, чтобы составить его полный образ, но некоторые, причем весьма важные, сведения живое существо получает.
Что же касается модулей и входящих в них столбов из нейронов, то не имеют ли выделяемые ими линии разной ориентации какого-нибудь отношения к голографии? Такой вопрос поставили перед собой сотрудники Лаборатории. И принялись показывать кошкам «кино» – решетки с разными пространственными частотами.
Почему именно решетки, а не что-нибудь другое? Откуда у Глезера и его коллег взялась уверенность, что найдутся нейроны, реагирующие не только на одиночную линию, но не на «зебры»? Прозорливость эта основывалась на сущности голографического процесса, прямо вытекала из анализа с помощью рядов Фурье.
Ведь граница между светлым и темным участками картинки – не что иное, как перепад яркостей. Значит, он может быть представлен совокупностью пространственных частот, может состоять из решеток с одной линией (ее-то и обнаруживали всегда, исследуя зрительную кору «по Хьюбелу и Визелу»), с тремя, пятью и так далее: таков и только таков ряд Фурье в подобном случае. Следовательно, если мозг действительно занимается голографией, если зрительная кора умеет делать такие преобразования, в ней обязаны находиться нейроны, настроенные на восприятие «зебр» с разным нечетным числом полосок.
Еще в 1966 году выдающийся английский нейрофизиолог Ф. Кэмпбелл установил, что зрительная система в целом работает подобно многоканальному Фурье-фильтру. Каждый канал такого фильтра настроен на выделение определенной пространственной частоты. Он доказал это так. Сначала испытуемому показывали решетку, у которой контраст между «прутьями» и «пустотой» был очень мал, однако таков, что решетка была все-таки заметна. Затем человек переводил взор на очень яркую, очень контрастную решетку, смотрел на нее примерно минуту и сразу же после этого пытался увидеть малоконтрастную. Но она как бы прикрывалась шапкой-невидимкой. Несмотря на все старания, испытуемый не видел ничего: мощный сигнал от контрастной решетки резко понизил чувствительность зрительного канала.
Ясно, что во всех трех случаях изображение попадало на разные участки сетчатки, так что об «утомлении» фоторецепторов не могло быть и речи. Чувствительность подавлялась на иных, более высоких участках тракта, по-видимому, в коре.
Если же «слабая» и «сильная» решетки резко различались по своим пространственным частотам, подавление не наступало, работали в каждом случае разные каналы приема зрительного сигнала.
Но действительно ли с корой связаны эти каналы? Кэмпбелл не смог тогда ответить на этот вопрос.
Ответ нашли сотрудники Лаборатории.
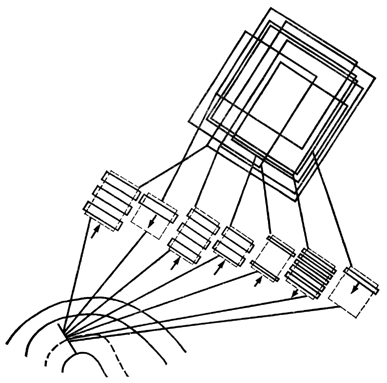
Рис. 46. Когда микроэлектрод идет строго перпендикулярно коре, он встречает нейроны, реагирующие на различные решетки, однако все эти решетки наклонены под одним и тем же углом
Во-первых, они отыскали нейроны, существование которых было предсказано ими, что называется, с помощью карандаша и бумаги. Одиночные полосы таким клеткам безразличны. Нейроны были бы признаны «молчащими», проверяй их по методике Хьюбела и Визела. Но полноценный сигнал немедленно появлялся, как только проекционный фонарь показывал кошке движущуюся решетку.
Второе открытие заключалось в том, что для целого ряда полей решетка должна выглядеть прямоугольником вполне определенной длины и ширины. Все пространство сетчатки анализируется корой по «полосатости» каждого кусочка – факт чрезвычайно важный, как мы очень скоро увидим.

Рис. 47. Косое движение микроэлектрода – и поля, перекрывая друг друга, располагаются под разными углами
Третье открытие оказалось самым сенсационным. Стало ясно, зачем в столбе любого модуля зрительной коры так много нейронов. Они вовсе не резервируют друг друга, как могло бы показаться (всем известна колоссальная надежность нейронных сетей!), дело в ином. Хотя все нейроны столба соединены с одним и тем же полем сетчатки, каждый дает максимальный ответ только на свою решетку, то есть на вполне определенную пространственную частоту. Математик скажет, что поле обладает «весовой функцией», которая в реальности представлена именно этой решеткой, а математически описана формулой с дифференциалами и интегралами.
Хотя и грубая, но наглядная аналогия работы поля, выделяющего «свою» решетку, – это человек, примеряющий шляпы в магазине. На его голову удобно сядет только шляпа вполне определенного размера, но, вообще говоря, напялить можно любую, хотя одни будут держаться еле-еле, а другие болтаться. Реакция примеряющего окажется каждый раз иной.
Так же и нейрон, обладающий данной весовой функцией, способен отозваться не только на оптимальную решетку, но и на другие. Как говорят, он обладает широким спектром входных сигналов, один из которых – «тот самый»». С позиций математического анализа любой ответ есть «свертка» весовой функции с распределением освещенности в данной решетке – операция, немного напоминающая умножение с последующим суммированием.
Выходит, объединенные в столб нейроны способны отреагировать на любую решетку данного наклона, попавшую на их рецептивное поле сетчатки. Каждый нейрон ответит по-своему, оптимально или не оптимально – это без разницы. В целом сформируется ансамбль ответов, подобный тому, который получался, когда робот анализировал картинку с помощью набора фильтров Фурье и фотоэлементов. Объединенные в модуль, все столбы своими сигналами обозначат решетку любого наклона и любой пространственной частоты.
Целостная картинка, имеющаяся на задней стенке глазного яблока, представлена на уровне затылочной коры сигналами модулей. Они разбивают эту картинку, превращают в огромное множество фрагментов (по числу полей). И каждый фрагмент, в свою очередь, выглядит внутри модуля ансамблем сигналов – результатом разложения по функциям Фурье.
То есть фрагмент представлен голографически. Точнее, кусочно-голографически, а уж если быть совсем точным – кусочно-квазиголографически. Почему «квази», то есть «как бы»?
– Потому что обычную голограмму непременно связывают с лазерами, когерентным излучением, сплетением опорного и предметного пучков, а здесь ничего этого нет, да и не нужно, – отвечает Глезер, выдвинувший эту гипотезу в 1970 г. – Ибо голография в точном смысле этого понятия есть разложение некоего колебательного процесса в ряд Фурье и запоминание того, что получилось. «Холос» – греческое слово, от которого получился термин, – означает «цельный, целостный». То есть речь идет о полной, во всех деталях, записи информации. Этим зрительный аппарат и занимается.
Проблема целостной записи относится, кстати, не только к зрению, но и физиологии восприятия вообще. Мелодию увертюры к опере «Кармен» человек запоминает не как последовательность звуков, а как некий образ, во всей полноте, так что потом мелодия звучит для него в любой тональности, с любыми вариациями, вплоть до джазовых синкоп, – вот какое широкое обобщение! Почти наверняка можно предположить, что когда доберутся до этой тайны, она окажется связанной с рядами Фурье и квазиголографией...
Ну а насчет зрения – голографический подход объясняет немало. Например, возникновение иллюзий, с чем иные модели зрительной системы плохо справляются.
Вадим Давыдович порылся в бумагах на столе и вытащил фотографию. На ней виднелись светлые пятна на темном фоне.
– Вот это голографическая интерпретация иллюзии Мюллера – Лиера...
Несмотря на мудреное название, иллюзия знакома всем, это линии с «хвостиками»: длина линий одинакова, но «хвостики» направлены у одной наружу, а у другой внутрь, так что в итоге линии кажутся разной длины. Долгое время считали, что глаз ошибается в размерах потому, что скользит от одного края картинки до другого. И если «хвостики» направлены в ту же сторону, что движение, взор «протягивается» по ним, психологически удлиняя размер. Наоборот, встречая противодействующие «хвостики», взор тормозится – линия кажется короче. Эту версию опроверг голографический эксперимент, а перед тем – психологический.

Рис. 48. Четыре голограммы (слева) и то, что они показывают (справа). В кружке иллюзия Мюллера-Лиера
Для психологического эксперимента изображение делали неподвижным относительно глаза: помещали диапозитив в миниатюрную присоску Ярбуса на глазном яблоке. Теперь водить зрачком по линиям возможности нет, а испытуемые все равно видят их иллюзорно разной длины.
Стало ясно, что рождается иллюзия не в глазу как таковом, а гораздо глубже: на уровне коры.
Голографический эксперимент заключался в том, что картинку с иллюзией Мюллера – Лиера превращали в голограмму, а потом выбрасывали из нее высокочастотные члены разложения Фурье. После этого восстановленное (и несколько менее четкое, естественно) изображение было именно таким, каким оно кажется: иллюзорно большая линия выглядит на голограмме действительно более длинной, нежели та, у которой «хвостики» вовнутрь.
Точно так же был вскрыт голографический механизм иллюзии «птичек» – разбросанных по листу бумаги равносторонних треугольников (они слева вверху на этой картинке).

Рис. 49. Голографический механизм иллюзии «птичек»
«Птички» летят, когда на них смотришь, то в одну сторону, то в другую, то в третью... Причина их своенравного поведения в том, что зрительный аппарат каждый раз использует не весь «мозговой фильтр Фурье», а только какую-то его часть.
Этот эффект великолепно демонстрирует ЭВМ: она показывает на экране телевизора «полет птичек» в любом направлении (нижний ряд). Для этого компьютер смотрит на треугольнички через Фурье-фильтр (справа вверху), соответственно синтезированный компьютером, но выбирает то одну часть фильтра, то другую, то третью (см. второй ряд изображений на рисунке справа).
А какой биологический, природный смысл был формировать такую сверхсложную опознающую мозговую систему, как Фурье-анализатор и кусочная квазиголография? Огромный.
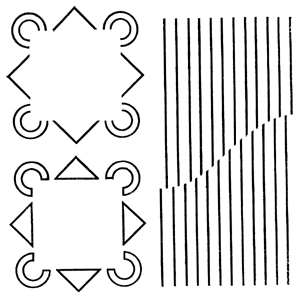
Рис. 50. Слева вверху – светлый квадрат, которого нет на самом деле. Справа – линия, которой также нет на самом деле. Причина? Голографическое устройство зрительного аппарата, обеспечивающее анализ текстур
Помните, рассматривая «обманы зрения», мы много говорили о текстурах? А теперь посмотрите вокруг: мир – это мозаика текстур. Но что такое эти текстуры, как не естественные, природой изготовленные решетки?
Все огромное многообразие природных текстур может быть представлено в затылочной коре соответствующим многообразием кодов – ансамблей ответов каждого модуля (паттернов, как для краткости называют такой ансамбль). То есть мозг получает сведения, насколько дробны и упорядочены элементы текстур, насколько и в каком направлении наклонены. Любая картинка оказывается представленной в затылочной коре некоторой мозаикой паттернов – соответственно множеству полей нейронов и множеству модулей коры.
Для каждой картинки, для каждого предмета мозаика оказывается особой. То есть получается мозаика признаков, позволяющих отличать предмет от предмета, пейзаж от пейзажа, лицо от лица...
– Мы назвали эти признаки простыми, – сказал Глезер. – С их помощью можно описать и запомнить любую картинку, от чрезвычайно пестрой до контурной, ведь гладкую поверхность можно рассматривать как выродившуюся «до нуля» текстуру. А математически это означает, что совокупность сигналов модулей выглядит многомерным пространством, и каждое конкретное изображение есть точка в нем.
Чтобы опознать изображение, надо сначала запомнить все признаки, и потом, когда поступит из сетчатки новое изображение, сравнить новые признаки с прежними, записанными в памяти. И если в итоге аппарат сравнения выйдет в прежнюю точку многомерного пространства, можно будет сказать: «Да, это тот же самый предмет». А если нет – нет.
Тут очень к месту вспомнить пульсирующие поля НКТ. Поскольку паттерны модулей затылочной коры опираются на эти поля, ясно, что в начальный момент после каждого саккадического скачка будут работать корковые нейроны, отвечающие лишь на самые низкие пространственные частоты. Только позже, по мере стягивания полей НКТ, станут восприниматься более высокие частоты, описываться все более тонкими составляющими паттернов.
Выходит, при запоминании картинки «точка» в многомерном пространстве простых признаков будет ставиться постепенно: сначала на основании грубых признаков, потом все более мелких. И соответственно опознание совершается после каждого скачка в несколько этапов; от грубого до наиподробнейшего. В этой последовательности – разгадка одного озадачившего всех опыта, произведенного в 1969 г. американскими физиологами.
Они формировали на телеэкране картинку так, что сначала появлялись низкие пространственные частоты (грубые контуры), а потом все более высокие (мелкие подробности). Если промежутки между появлением этих частот были невелики, человек не отличал такое последовательное изображение от обычного, когда все частоты подавались на экран одновременно. Сейчас понятно, почему картинки казались одинаковым: темп появления высоких пространственных частот соответствовал стягиванию полей НКТ, и для зрительного аппарата было безразлично, что высокочастотные составляющие пришли в мозг чуть позже низкочастотных.
– Вадим Давыдович, – спросил я, – а как же в таком случае мы видим контуры? Поля покажут характер текстуры, размер области, которую она заполняет, – где же контурная линия?
– Этим занимаются особые поля, только для того и предназначенные. Когда мы их обнаружили, нас удивило, что они выглядят «трехслойным пирогом»: средняя часть возбуждает нейрон коры, а боковины тормозят. Если такая текстура попадает на все поле или, что то же самое, поле оказывается внутри текстуры, сигнала нет, тормозные фланги подавили возбуждающий сигнал центра. Однако едва хотя бы один фланг вышел за границу, попал на иную текстуру или даже на такую же, но по-иному повернутую своими элементами, возбуждающий центр поля пересиливает торможение одиночного фланга. В итоге контур подобраза оказывается выделен цепочкой, составленной возбужденными нейронами, – можно сказать, выстрижен из фона примерно так же, как хозяйка обводит колесиком с зубцами линии выкройки...
– Вы сказали – подобраза? Что это значит?
– Так называются текстурно однородные участки. Взгляните на дерево на большом лугу: в картине три текстурных подобраза – ствол, крона и луг как фон. Или возьмите любую гравюру: все мастерство художника построено именно на умении пользоваться разнообразием текстурных подобразов. Впрочем, подобраз не есть что-то абсолютное и приговоренное навсегда им оставаться. Для образа «дерево» подобразами будут «ствол» и «крона», но та же самая крона – образ для подобразов «ветка» и «лист».
Мир велик и разнообразен, и столь же разнообразна иерархия образов и подобразов. Но еще важнее, что открылась, наконец, причина, ради которой поставила природа между сетчаткой и выходными сигналами затылочной коры столько преобразующих ступенек. Без этих преобразований нет возможности выделить текстурный подобраз из фона, созданного иной текстурой, а такая задача для живого существа жизненно необходима, как для хищника, так и для жертвы.
К тому же описание мира через систему подобразов оказывается очень экономичным. В чем заключается цель зрения и в конечном счете мышления? Установить, каково пространственное расположение предметов и как оно изменяется во времени. Это самая общая формулировка любой познавательной деятельности. Сейчас наша задача скромна: нужно всего лишь увидеть, как возле дерева на лугу ходит человек. И действительно, передвижение его подобраза в ретинотопическом пространстве заднетеменной коры не затрагивает подобразов «луг» и «дерево». Мозгу, следовательно, нужно реагировать только на сравнительно небольшие изменения общей картины, и обработка информации получается наиболее простой. А покупать победу малыми силами – первое требование, которое природа предъявляет к живому. Организм ведь существует не ради информации, а ради того, чтобы жить...
И уж коль затронута ретинотопика, надо отметить, что «выстриженные из образов» подобразы (а значит, и образы тоже) располагаются в нейронном пространстве затылочной коры именно в таких между собой взаимоотношениях, в каких они спроецированы хрусталиком на сетчатку. Так, как на самом деле располагаются предметы в пространстве перед нами. Но – и это принципиально важный момент! – мы в нейронных сетях затылочной коры не увидим никаких контуров.
То, что существует реально на сетчатке как контурная картинка, предстает в коре столь же реально, но математической моделью контуров и пространства. То есть подобно тому, как в аналитической геометрии нет линий, точек, плоскостей и так далее, а имеются одни уравнения. Примерно такой математикой с помощью кусочной квазиголографии и занимается мозг.
В итоге мы видим открывающееся перед нами пространство не только в виде светлых и темных пятен, но и в виде текстурных участков, имеющих четко очерченные границы. Видим сразу по всему полю зрения. А так как плотность расположения фоторецепторов неравномерна по пространству сетчатки, получается картинка разной четкости. И мы направляем центральную ямку, область наиболее четкого зрения, туда, где хотим разглядеть что-нибудь пояснее.
– Хотелось бы обратить внимание на то, – сказал Глезер, – что квазиголографический механизм, формирующий простые признаки деталей изображения, сформирован в зрительной системе генетически. Это значит, что он у всех людей одинаков. Поэтому если мы с вами смотрим вместе на какую-то вещь, она отражается в нейронных сетях затылочной коры у меня и у вас одинаково. Вот что дальше будет, какой смысл каждый извлечет из полученной картины, – это зависит от образования, жизненного опыта и так далее, словом, от социальных факторов. Но зрительная основа образов у всех одна.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава седьмая Искусственные Эго Машины
Глава седьмая Искусственные Эго Машины Начиная с этого момента, мы будем называть любую систему, способную к порождению сознательной самости, Эго Машиной. Эго Машина не обязательно должна быть живым существом; она может быть чем угодно, что обладает сознательной
Глава седьмая Новейшие ключи к происхождению человека
Глава седьмая Новейшие ключи к происхождению человека Глубочайшие истины средневековья теперь вызывают у школьников только смех. Пройдет еще два-три века, и над глубочайшими истинами американской демократии будут смеяться даже школьные учителя. Генри Л. Менкен
Глава седьмая Система животного мира
Глава седьмая Система животного мира Со времен классической древности, начиная от Аристотеля[107] и Плиния, и в течение первых 10 веков нашей эры, наука о животных — зоология, представляла собою собрание разрозненных сведений и рассказов о животных, об их наружном виде и
ГЛАВА СЕДЬМАЯ ЭМУ ПРОИГРЫВАЮТ ВОЙНУ
ГЛАВА СЕДЬМАЯ ЭМУ ПРОИГРЫВАЮТ ВОЙНУ Вторая по величине птица на Земле. — Почему их назвали эму. — С пулемётами против страусов. — Отгородимся от них забором — а там хоть трава не расти. — Преданные папаши и легкомысленные мамаши. —Два дня страусёнок просидел возле
Глава седьмая Крабовые собаки и птицы-плотники
Глава седьмая Крабовые собаки и птицы-плотники Вот уже сутки, как мы снова в Джорджтауне; муравьед и прочие животные как следует устроены в клетках и вполне освоились со своим новым положением. После просторов Рупунуни нам с Бобом было тесно и непокойно в городе, и мы
Глава седьмая. ПЕВЦЫ НА ДЕРЕВЬЯХ
Глава седьмая. ПЕВЦЫ НА ДЕРЕВЬЯХ Из нор появились ползучие твари И воззрились на них с удивлением. «Охота Ворчуна» Таман Негара (прежнее название — Национальный парк имени короля Георга V) был создан в 1937 году. Это огромный сплошной массив девственного леса площадью
Глава седьмая Кванты Упрощение понимания
Глава седьмая Кванты Упрощение понимания Если кто-то заявляет, что знает, что такое квантовая теория, он не понял ее. Ричард Фейнман Великая идея: волны ведут себя как частицы, а частицы ведут себя как волныМы зависли на краю квантовой теории, погрузив большой палец ноги в
Тайнам нет конца
Тайнам нет конца Совершив краткие путешествия в букет цветов или энтомозаказник и познакомившись бегло с родственницами мелиттобий — эвхаритидами, мы вернемся к нашим крохотным героиням, но уже совсем ненадолго, так как я давно уже начал подозревать, что мои воспетые
Глава седьмая Забота о здоровье
Глава седьмая Забота о здоровье Телу животного, непосредственно соприкасающемуся с внешней средой, здорово достается. Удивительно, как оно выдерживает такую трепку и сохраняется так долго. Это происходит благодаря чудесной системе восстановления тканей и, кроме того,
Глава седьмая Болюс «Черри»
Глава седьмая Болюс «Черри» Что исследуют в лаборатории оральных процессов?Рассказывая о «Долине еды», хочется описывать это местечко как Силиконовую долину питания. Пятнадцать тысяч ученых неустанно трудятся над улучшением пищевых продуктов или, в зависимости от
Глава 9 Новый взгляд на любовь
Глава 9 Новый взгляд на любовь Представленная нами гипотеза может кому-то показаться довольно мрачной. Любовь – это зависимость, причем не в образном, а в прямом смысле слова. Некоторые из нас от природы склонны к внебрачному сексу. Даже пингвины на полюсе, те самые, из