Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация
В определении понятия «гормон» было указано несколько этапов распространения гормонального сигнала (рис. 2.6).
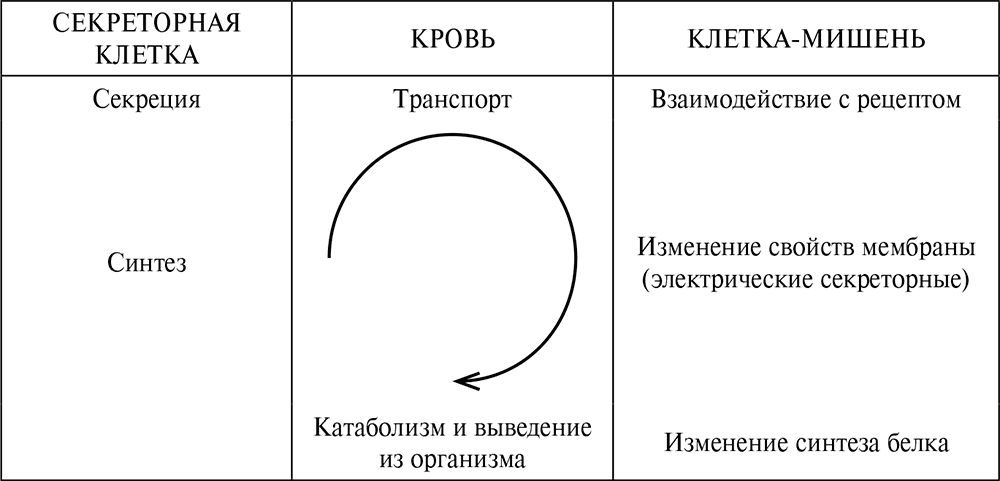
Рис. 2.6. Этапы распространения гормонального сигнала. Внутри клетки происходит синтез гормона. Секреция – не пассивное выделение вещества в окружающее пространство, а активный процесс, на который могут влиять факторы, не изменяющие интенсивность синтеза. В крови гормоны связываются с белками-носителями. В связанной форме гормоны неактивны. Таким образом, их биологический эффект зависит и от содержания в крови транспортных белков. Для реализации биологического эффекта гормон должен связаться с клеточным рецептором – сложной структурой, расположенной внутри клеточной мембраны или внутри клетки, в ее цитозоле. После связывания молекулы гормона с рецептором следует целый каскад химических реакций, которые приводят к изменению активности клетки. Это проявляется в изменении синтеза белка в клетке, а также в изменении свойств ее мембраны, которые происходят при передаче нервного импульса, сокращении мышечных клеток и секреции из них различных веществ. Освободившись из комплекса с рецептором, молекула гормона инактивируется в крови (пептиды) или в печени (стероиды). К изменению гормонального эффекта приводят изменения не только синтеза молекул гормона в эндокринной железе, но и на любом этапе передачи гормонального сигнала
Следует различать этапы синтеза и секреции как два независимых процесса. В этой книге выражения «изменение (увеличение, снижение) секреции» и «изменение синтеза» обычно употребляются как синонимы, но это сделано только для облегчения восприятия. В действительности секреция не является пассивным процессом выделения из клетки вещества вследствие его накопления. Это многостадийный процесс, на который влияют как внутри-, так и внеклеточные регуляторы. Зачастую эти два процесса – синтез и секреция – оказываются несогласованными, и при увеличении синтеза не происходит увеличения секреции. Таким образом, некое воздействие, о котором известно, что оно приводит к увеличению синтеза гормона, не обязательно ведет к увеличению содержания этого гормона в крови.
Гормон, поступивший в кровь, переносится к тканям-мишеням в разных формах. Определенная доля молекул находится в том виде, в котором они были секретированы, но б?льшая часть связывается с белками крови. Некоторые из этих белков связывают многие гормоны (альбумин), а другие обладают высокой специфичностью по отношению к определенным гормонам, т. е. связывают только один или строго ограниченный круг гормонов. (Так, кортикостероид-связывающий глобулин связывает глюкокортикоиды, минералокортикоиды и прогестины, но не связывает эстрогены; белок, который связывает пролактин и гормон роста, не присоединяет другие пептидные гормоны.) Связывание гормонов белками крови имеет три функции.
Первая функция – регуляторная. Связанный гормон не взаимодействует с тканями-мишенями, т. е. та часть молекул секретированного гормона, которая связалась с белком, биологически не активна.
Реализация гормонального сигнала происходит в пять этапов: 1) синтез гормона 2) секреция гормона 3) транспорт гормона 4) взаимодействие гормона с клеточным рецептором 5) инактивация гормона и вывод его из организма
В результате эффективная концентрация гормона всегда меньше, чем его общая концентрация в крови. Чем выше в ней содержание белка, связывающего молекулы гормона, тем меньше эффективная концентрация гормона, т. е. возможный биологический эффект. Следовательно, биологическая активность гормона регулируется содержанием в крови белков, которые связывают этот гормон. Например, при беременности возрастает количество белка, связывающего кортизол. При возникновении потребности тканей в гормоне увеличиваются расщепление гормон-белкового комплекса и эффективная концентрация, а содержание биологически активного гормона в крови возрастает. Таким образом, регулирующая функция белков, связывающих гормоны, тесно связана со второй функцией – запасающей.
Вторая – запасающая — функция связывания гормонов с белками крови определяется тем, что в крови имеется постоянный запас гормона, который может быть легко переведен в активное состояние. При снижении концентрации свободного гормона часть связанного высвобождается из комплекса с белком.
Третья функция связывания молекул гормона белками крови – транспортная. В некоторых случаях гормон, связанный с транспортным белком, быстрее проникает в клетку-мишень.
После синтеза, секреции и транспорта начинается взаимодействие гормона с клеткой-мишенью. Оно обеспечивается клеточными рецепторами. Они представляют собой сложные белки и расположены в клеточной мембране (рецепторы пептидных гормонов), в цитоплазме клетки (рецепторы стероидных гормонов) и в клеточном ядре (рецепторы трийодтиронина и тироксина).
Основная характеристика рецепторов – их специфичность. Благодаря этому свойству рецепторов гормоны, распространяясь по всему организму, действуют не на все ткани и органы, а только на те, клетки которых содержат рецепторы данного гормона. Рецепторы обладают гораздо более высокой специфичностью, чем транспортные белки крови. Почти все рецепторы связывают только один определенный гормон, причем в гораздо большей степени, чем его ближайшие химические аналоги. В то же время большинство гормонов связываются с несколькими подтипами рецепторов, и специфичность этих подтипов различна. Таким образом достигается тонкое регулирование функций клетки-мишени.
Связывание гормона с рецептором вызывает цепь сложных биохимических реакций, конечным итогом которых является изменение синтеза определенных белков, что и принято считать биологическим эффектом данного гормона. Поскольку гормональный эффект проявляется только в результате взаимодействия с рецептором, некоторые исследователи полагают, что ключевая молекула в системе гуморальной регуляции – рецептор, а не гормон. Не останавливаясь здесь на критике этой концепции (неполное соответствие гормонального эффекта и количества рецептора в ткани и т. п.), отметим, что мы будем рассматривать именно гормоны как основное звено в системах гуморальной регуляции по следующим причинам.
Во-первых, методически определить гормоны гораздо легче, чем рецепторы. Финансовые затраты на приобретение реактивов для определения гормонов значительно меньше, чем на покупку реактивов для установления содержания рецепторов. Самые сложные из гормонов представляют собой цепочку из нескольких десятков аминокислот. В то же время рецепторы – это сложные белки, которые могут состоять из десятков тысяч аминокислот, обладают сложной структурой и легко разрушаются при биохимических манипуляциях. Концентрация гормонов в тканях организма значительно выше концентрации рецепторов в тех же тканях. Часто для определения содержания гормона достаточно одной сотой миллилитра крови, а для анализа рецепторов требуется порой несколько граммов ткани. Гормоны находятся в крови, часть которой можно взять для анализа почти безболезненно и без существенных последствий для живого организма. Рецепторы находятся в тканях, что требует более серьезного вторжения в организм. В некоторых случаях вовсе невозможно определить содержание рецепторов у живого организма, например в структурах головного мозга человека.
Во-вторых, мы не можем в процессе эксперимента менять содержание рецепторов в тканях, тогда как изменить уровень гормона удается легко и с незначительными финансовыми затратами. Довольно просто ввести дополнительное количество гормона, сделав инъекцию, а уменьшить его количество можно введением вещества, блокирующего синтез или секрецию гормона, а также удалив отдельную железу (в эксперименте на животном). Молекулярно-генетические методы, позволяющие изменить количество рецепторов в тканях, появились только в последние годы. Они остаются еще очень дорогими и громоздкими, а кроме того, имеют существенные методические ограничения.
Таким образом, экспериментальных данных о содержании гормонов в крови при разных состояниях организма в тысячи раз больше, чем о содержании рецепторов в тканях. Что же касается работ, выполненных на человеке, то количество исследований рецепторов в миллионы раз меньше, чем количество исследований уровня гормонов. Таким образом, в этой книге основное внимание уделено изменению уровня гормона в крови, а не активности рецепторного аппарата клетки.
Итак, первые четыре этапа передачи гормонального сигнала – это синтез, секреция, транспорт белками крови и связывание с рецепторным аппаратом в клетках тканей. Заключительным, пятым, этапом передачи гуморального сигнала является инактивация гормона и вывод его из организма. Нарушение метаболизма гормона может привести к изменениям в работе всей эндокринной системы. При замедленной инактивации однократное введение гормона способно вызвать длительный или же неожиданно сильный эффект. В норме время полужизни пептидных гормонов составляет несколько минут, а стероидных – несколько часов. Многие синтетические производные гормонов значительно эффективнее природных аналогов именно потому, что они медленно подвергаются инактивации в процессе обмена веществ.
Рассматривая здесь гормон как основное звено эндокринной системы, будем все-таки помнить, что уровень гормона в крови – не исчерпывающая характеристика состояния системы «железа – гормон – кровь – орган-мишень – биологический эффект». Общее состояние эндокринной системы зависит от этапов синтеза и секреции гормона, его транспорта, т. е. связывания белками крови, от состояния системы рецепторов в клетке-мишени и метаболизма гормона.
Биологическая активность эндокринной системы может меняться, хотя содержание гормона в крови остается неизменным. Изменения связаны с нарушениями в транспорте, рецепции или катаболизме молекул гормона. Например, длительный прием оральных контрацептивов достаточно часто приводит к нарушениям либидо. У таких женщин обнаружено повышенное содержание транспортного белка – глобулина, связывающего эстрогены. Другой пример – для прерывания беременности на ранних сроках применяются вещества, нарушающие связывание прогестерона с его клеточными рецепторами. Биологический эффект обеспечивается изменением этапа взаимодействия гормона с рецептором. Еще один пример – препараты лакрицы (солодки), широко используемые для лечения бронхитов, трахеитов и прочих болезней верхних дыхательных путей. Эти лекарства содержат вещества, тормозящие инактивацию глюкокортикоидов в процессе обмена веществ. В результате повышается содержание в крови глюкокортикоидов, прежде всего кортизола, подавляющего воспалительные процессы. Эти лекарства не влияют на систему синтеза и секреции гормонов и не нарушают ее функционирование. Поэтому препараты лакрицы широко применяются как безопасные противовоспалительные средства.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
2. Действие стероидных гормонов
2. Действие стероидных гормонов Стероидные гормоны являются относительно простыми органическими соединениями с небольшим молекулярным весом. О механизме их действия известно сейчас больше, чем о действии других гормонов. Скелет стероидных гормонов образован четырьмя
3. Действие белковых гормонов
3. Действие белковых гормонов Полипептидные гормоны известны, по-видимому, еще далеко не все. Среди них есть настоящие белки, хотя и небольшого молекулярного веса, а есть и полипептиды, состоящие всего из 8—11 аминокислот, замкнутых в кольцо. Настоящие белки синтезируются
Клетки-коллективисты и клетки-одиночки
Клетки-коллективисты и клетки-одиночки В основе тесной кооперации клеток, входящих в состав многоклеточного организма, лежат по меньшей мере две важнейшие причины. Во-первых, каждая отдельно взятая клетка, будучи сама по себе на редкость умелым и исполнительным
Глава XVIII Речной транспорт
Глава XVIII Речной транспорт Первая клетка не смогла бы выжить, не будь особого «климата» жизни, создаваемого морем. Точно так же каждая из сотен триллионов клеток, составляющих организм человека, погибла бы без крови и лимфы. На протяжении миллионов лет, с тех пор как
КЛЕТОЧНОЕ СТРОЕНИЕ ОРГАНИЗМОВ СТРОЕНИЕ КЛЕТКИ. ПРИБОРЫ ДЛЯ ИССЛЕДОВАНИЯ СТРОЕНИЯ КЛЕТКИ
КЛЕТОЧНОЕ СТРОЕНИЕ ОРГАНИЗМОВ СТРОЕНИЕ КЛЕТКИ. ПРИБОРЫ ДЛЯ ИССЛЕДОВАНИЯ СТРОЕНИЯ КЛЕТКИ 1. Выберите один наиболее правильный ответ.Клетка – это:A. Мельчайшая частица всего живогоБ. Мельчайшая частица живого растенияB. Часть растенияГ. Искусственно созданная единица для
2. ПЕРЕДАЧА И ПРЕОБРАЗОВАНИЕ СИГНАЛОВ
2. ПЕРЕДАЧА И ПРЕОБРАЗОВАНИЕ СИГНАЛОВ Процессы передачи и преобразования сигналов обеспечивают поступление в высшие сенсорные центры наиболее важной (существенной) информации о сенсорном событии в такой форме, которая удобна для надёжного и быстрого анализа. Что
5.6. Передача и переработка соматосенсорной информации
5.6. Передача и переработка соматосенсорной информации Чувствительность кожи и ощущение движения связаны с проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спиноталамическому, значительно различающимся по своим
2.1. Общая характеристика сигнала
2.1. Общая характеристика сигнала ССП выделяют при помощи специальных методов из ЭЭГ. Частотный диапазон ССП включает полосу от 0 Гц до 3 кГц и ограничен, с одной стороны, сверхмедленной электрической активностью мозга [Илюхина, 1977], а с другой – спайковой активностью
2.3. Считывание сигнала
2.3. Считывание сигнала Для компьютерной обработки сигнала «сырая» ЭЭГ переводится в цифровую форму. Частота считывания мгновенных значений сигнала при этой процедуре («квантование», sampling) определяется согласно теореме Шеннона–Котельникова. Для того чтобы описать
Механизм передачи гормонального сигнала через внутриклеточные рецепторы
Механизм передачи гормонального сигнала через внутриклеточные рецепторы Передача сигнала гормонов с липофильными свойствами (стероидные гормоны) и тироксина возможна при прохождении их через плазматическую мембрану клеток-мишеней. Рецепторы гормонов находятся в
Транспорт глюкозы из крови в клетки
Транспорт глюкозы из крови в клетки Поглощение глюкозы клетками из кровотока происходит, также путем облегченной диффузии. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента ее концентрации. Исключение составляют клетки мышц и жировой
Глава 19. Липиды тканей, переваривание и транспорт липидов
Глава 19. Липиды тканей, переваривание и транспорт липидов Липиды – неоднородная в химическом отношении группа веществ биологического происхождения, общим свойством которых является гидрофобность и способность растворяться в неполярных органических растворителях.
Глава 22. ПЕРЕДАЧА ЭЛЕКТРОНОВ
Глава 22. ПЕРЕДАЧА ЭЛЕКТРОНОВ Львиная доля производимой в процессе обмена веществ энергии выделяется в ходе реакций, в которых участвует атмосферный кислород. Перед тем как мы сможем свободно рассуждать об этих реакциях, надо сначала обговорить основные
Роль гормонального статуса и возраста в половом поведении
Роль гормонального статуса и возраста в половом поведении Фазы полового цикла у самок приматов и их связь с обменом веществ, высшей нервной деятельностью и социальными структурами детально описаны Л. В. Алексеевой. Гормональная регуляция полового поведения лучше всею
Жуков. Дмитрий Анатольевич
Просмотр ограничен
Смотрите доступные для ознакомления главы 👉