Дети севера
Дети севера
Суровые природные условия накладывают свой специфический отпечаток как на образ жизни людей, населяющих высокоширотные районы, так и на их анатомию. Известно, что человеческие популяции, традиционно обитающие в районах с холодным климатом — например, саамы Кольского полуострова или эскимосы Гренландии, — отличаются от коренного населения низких широт целым рядом особенностей строения скелета. Такими особенностями являются, например, укороченные по отношению к длине туловища конечности, сравнительно большая по отношению к росту масса тела, утолщённые трубчатые кости и т. д. Все эти черты наблюдаются и у неандертальцев, которые по форме и пропорциям тела намного ближе к современным жителям Чукотки, Аляски и Гренландии (речь, разумеется, о коренном населении этих регионов), чем к африканцам или, скажем, палеолитическим гомо сапиенс Европы — свежеприбывшим мигрантам из той же Африки.
Особенно показательны в этом отношении два индекса, широко используемые в физической антропологии. Один из них называется круральным и характеризует пропорции ног, а второй называется брахиальным и характеризует пропорции рук. Первый высчитывается как процентное соотношение длины большеберцовой и бедренной костей (длину большеберцовой кости умножить на 100 и разделить на длину бедренной кости), а второй как процентное соотношение длины лучевой и плечевой костей (формула расчёта аналогична).
Давно замечено и многократно подтверждено специально проводившимися измерениями, что у коренных жителей высокоширотных регионов оба этих индекса значительно меньше, чем у жителей тропиков, субтропиков и даже районов с умеренным климатом. Иными словами, у саамов, эскимосов, чукчей и представителей других северных народов нога ниже колена и рука ниже локтя намного короче по отношению к верхним частям этих конечностей, чем, скажем, у центральноафриканских пигмеев, или аборигенов Австралии. Совершенно очевидно, что неандертальцы по обоим этим показателям близки к жителям севера, а палеолитические гомо сапиенс — даже европейские — больше тяготеют к обитателям низких широт, как того и следовало бы ожидать от недавних выходцев из тропической зоны (рис. 4.5–4.7).
Первым, кто обратил серьёзное внимание на существование комплекса сходных особенностей в анатомии ископаемых и современных обитателей северных областей, был американский антрополог К. Кун. Он же выдвинул и обосновал гипотезу, что размер и форма тела аборигенов высоких широт, включая неандертальцев — это следствие адаптации к холодному климату, где особое значение приобретает способность минимизировать потери тепла и энергии[92]. Массивность тела, имеющая следствием уменьшение площади его поверхности, приходящейся на единицу объёма, помогает сэкономить и то, и другое. Не случайно у людей наблюдается обратная связь между географической широтой обитания, с одной стороны, и отношением площади поверхности тела к его массе — с другой. Чем больше первая величина, тем в среднем меньше вторая (рис. 4.8). У широко расселённых форм млекопитающих тоже представители северных видов или подвидов, как правило, крупнее своих южных сородичей (так называемое правило Бергмана). Например, уссурийские тигры превосходят по размеру бенгальских и яванских, северные олени — благородных, белые медведи — бурых и т. д.
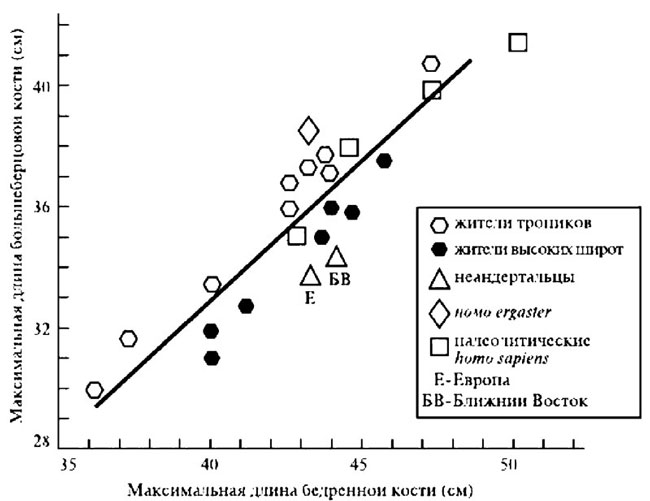
Рис. 4.5. Соотношение длины большеберцовой и бедренной костей у разных групп современных и ископаемых людей. Очевидно, что неандертальцы по этому показателю близки жителям севера, а палеолитические гомо сапиенс больше тяготеют к обитателям низких широт, как того и следовало бы ожидать от выходцев из тропической зоны (источник: Lewin and Foley 2004)
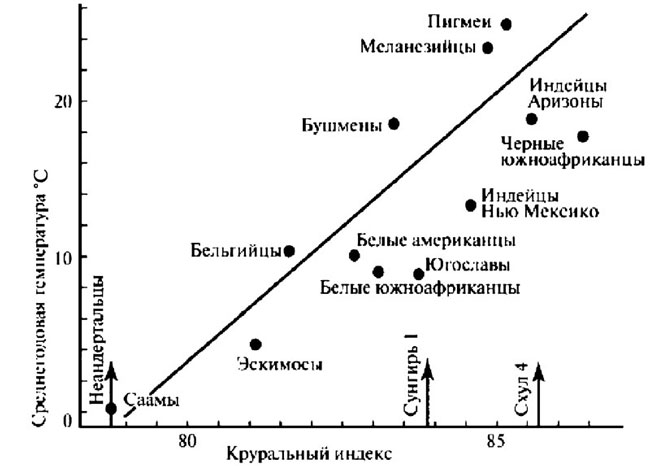
Рис. 4.6. Соотношение среднегодовой температуры с круральным индексом, рассчитанным для разных групп современных людей. Стрелками показано, какое место занимали бы в этой картине неандертальцы, ранние гомо сапиенс Ближнего Востока (возраст около 100 тыс. лет назад) и верхнепалеолитические гомо сапиенс Восточной Европы (возраст около 25 тыс. лет назад). Источник: Lewin and Foley 2004, с изменениями и дополнениями
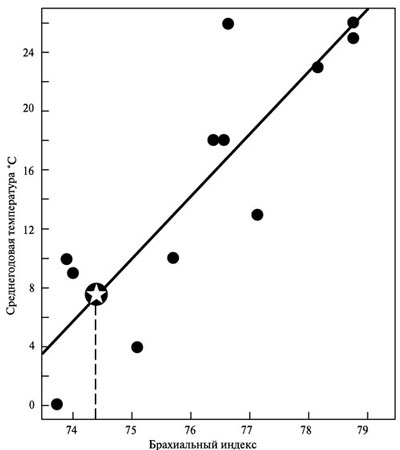
Рис. 4.7. Соотношение среднегодовой температуры с брахиальным индексом, рассчитанным для 12 разных групп современных людей. Звёздочка и исходящая из неё пунктирная линия показывают, какое место занимали бы в этой картине неандертальцы
Кстати, коль уж я упомянул медведей, обязательно нужно сказать хотя бы несколько слов о самом, может быть, знаменитом представителе этого славного семейства, который был «земляком» и современником неандертальцев, а очень часто ещё и их ближайшим соседом «по квартире». Речь, разумеется, о пещерном медведе — Ursus spelaeus. Количество совпадений и общих черт в истории и биологии этих двух видов настолько велико, что человек, склонный к мистике, вполне может заподозрить здесь некую сверхъестественную связь.
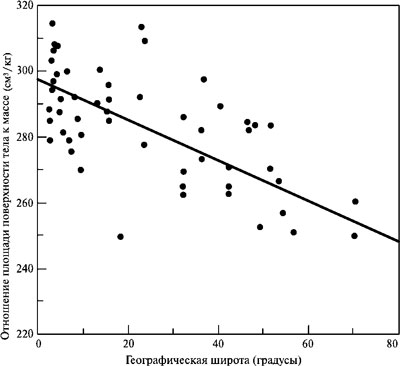
Рис. 4.8. Соотношение площади поверхности тела и его объёма у жителей высокоширотных районов и жителей регионов с умеренным или жарким климатом (источник: Lewin and Foley 2004)
Начнём с того, что пещерные медведи, как и неандертальцы, происходят от европейских представителей вида, имевшего более широкое распространение (медведь Денингера — Ursus deningeri). Их ареалы в значительной степени перекрываются, а периоды существования совпадают почти полностью, если не полностью: от конца среднего плейстоцена до последнего ледникового максимума[93]. Пик процветания и тех, и других, судя по численности ископаемых находок, тоже приходится на одно и то же время, а именно на конец изотопной стадии 4 и первую половину стадии 3 (примерно от 65 до 35/40 тыс. лет назад). В отличие от предковых видов — человека гейдельбергского и медведя Денингера, предпочитавших, за редкими исключениями, равнины и низкогорья, неандертальцы и пещерные медведи питали явную склонность к жизни высоко в горах: следы их пребывания часто встречаются на высоте более 2000 м над уровнем моря. Наконец, нельзя не отметить и ещё одно совпадение: оба вида не только исчезли примерно синхронно, но и были замещены более грацильными формами с более разнообразным, как считает большинство исследователей, рационом. При этом обращает на себя внимание то обстоятельство, что среди анатомических особенностей, отличающих пещерных медведей от сменивших их бурых (Ursus arctos), есть целый ряд черт, по которым и неандертальцы тоже отличались от пришедших им на смену гомо сапиенс. Кроме большей массивности тела, список таких черт включает ещё, например, относительно укороченные дистальные сегменты конечностей, увеличенные размеры суставов трубчатых костей, утолщённые фаланги, сравнительно крупные коренные зубы и некоторые другие признаки[94]. В обоих случаях часть этих особенностей явно была вызвана к жизни необходимостью обеспечить их обладателям более эффективную терморегуляцию.
Но достаточно о медведях (впрочем, нам ещё предстоит встретиться с ними в главе 7), вернёмся к неандертальцам. Можно не сомневаться, что помимо размеров и пропорций тела они должны были иметь и какие-то чисто физиологические средства приспособления к холодному климату. Первым эту идею тоже развил К. Кун, и он же указал на некоторые анатомические детали, которые её подтверждают. В частности, Кун обратил внимание на необычайно крупный размер подглазничных отверстий на верхних челюстях неандертальцев, с одной стороны, и эскимосов Гренландии — с другой. Поскольку через эти отверстия проходят сосуды, обеспечивающие приток крови к щекам, логичным кажется предположение, что их увеличение у коренных жителей севера было связано с необходимостью более интенсивного обогрева во избежание обморожения. Аналогичным образом можно истолковать и ещё несколько специфических черт в строении неандертальских костей.
Таким образом, есть все основания считать, что суровый и притом крайне нестабильный климат плейстоценовой Европы действительно оказал большое воздействие на эволюцию и анатомию неандертальцев. Это воздействие отразилось, в частности, в пропорциях их скелета, в некоторых деталях системы кровоснабжения и ряде других признаков, адаптивный смысл которых кажется более или менее понятным. Со многими другими неандертальскими особенностями, однако, ясности гораздо меньше. Какова была их адаптивная роль, и играли ли они вообще такую роль, остается неизвестно.
Одной из наиболее трудноразрешимых проблем является объяснение специфического строения неандертальского носового отверстия — более широкого и глубокого, чем у современных людей. Популярная некогда гипотеза, согласно которой это было необходимо для предотвращения попадания слишком холодного и сухого воздуха в легкие при дыхании, вступает в непреодолимое противоречие с тем фактом, что у современных людей широкий нос в норме гораздо больше характерен для обитателей регионов с мягким климатом, нежели для жителей севера. Про неандертальское носовое отверстие написаны десятки статей, но какие именно факторы обусловили его специфический формат — по-прежнему непонятно. Возможно, что холод, как и климат вообще, тут был совершенно ни при чём, а определяющую роль сыграли чисто анатомические закономерности и требования, накладываемые общими особенностями строения лицевого скелета неандертальцев. Например, не исключено, что широкий нос был всего лишь механическим следствием свойственного им среднелицевого прогнатизма[95]. Правда, носовое отверстие неандертальцев характеризуется ещё и возросшей по сравнению с их предшественниками высотой, и эта черта, по мнению ряда авторов, могла быть результатом взаимодействия структурных ограничений, налагаемых особенностями исходного (предкового) состояния, с одной стороны, и адаптивных изменений в ответ на специфические требования природной среды — с другой[96].
Многие другие особенности неандертальского скелета, возможно, представляют собой результат адаптации не столько к холодному климату, сколько к большим физическим нагрузкам. Таковы, например, толстые стенки трубчатых костей, сравнительно сильно изогнутые проксимальные сегменты конечностей, хорошо выраженные (рельефные) участки крепления мускулов, широкие (как бы расплющенные) фаланги пальцев ног и некоторые другие признаки. Не исключено, что, по крайней мере, некоторые из них не были предопределены на генетическом уровне, а образовывались (или, по крайней мере, усугублялись) в течение жизни человека вследствие частых и долгих переходов, транспортировки тяжестей без вспомогательных средств, выслеживания и преследования охотничьей добычи, столкновений с крупными и опасными животными и т. д.
Наконец, ещё какая-то часть свойственных всем неандертальцам или отдельным их группам признаков могла закрепиться у них просто случайно, в результате процесса, который биологи называют дрейфом генов[97]. Воздействию этого процесса более всего подвержены небольшие по численности популяции, особенно когда они оказываются в условиях полной или частичной изоляции от других популяций своего вида. В таких условиях аллели (т. е. варианты состояния или, иначе говоря, формы генов), бывшие ранее редкими, могут быстро стать преобладающими. Например, при дроблении популяции в ходе расселения или резком уменьшении её численности вследствие какой-либо катастрофы генетический состав вновь образовавшихся или уцелевших групп почти наверняка будет во многом отличаться от первоначального, предкового. Первопроходцы, уходящие на новые земли, унесут с собой лишь часть существующих аллелей, часть исходного генетического разнообразия, и чем меньше их (первопроходцев и аллелей) будет, тем больше вероятность того, что в основанной ими новой популяции нормой станет то, что раньше было отклонением от неё. Точно так же норма и отклонение на уровне вида, популяции, или хотя бы отдельной группы могут поменяться местами после массовой гибели в результате, скажем, извержения вулкана, слишком холодной и долгой зимы, схода снежной лавины, обвала в пещере, либо каких-то ещё природных катаклизмов. Более чем вероятно, что нечто подобное не раз происходило и с неандертальцами, которые, напомню, ведут своё происхождение от группы (скорее всего, очень небольшой) переселенцев из Африки, и которым выпало жить далеко не в самом благодатном краю и не в самые лёгкие времена.