Признаки, указывающие на оптимального полового партнера
Признаки, указывающие на оптимального полового партнера
Сегодня специалисты в области поведенческой экологии предлагают для объяснения механизмов женской избирательности и формирования вторичных половых признаков «паразитарную» гипотезу. Следуя этой гипотезе, самцы, обладающие более длинным хвостом и ярким оперением, имеют и более устойчивую иммунную систему (Рис. 3.7). Хорошее состояние перьевого покрова служит надежным индикатором отменного здоровья и хорошей физической формы его обладателя. Выбирая партнера с такими признаками, самки обеспечивают лучшее качество своих детей. Несомненно, эта гипотеза имеет под собой веские основания. Но почему у всех самцов данного вида, больных и здоровых, хвосты вырастают длиннее, чем нужно для полета?
Ответ на этот вопрос можно найти в теории эволюции полового поведения, разработанной Р. А. Фишером, и являющейся непосредственным продолжением развития теории полового отбора Ч. Дарвина. Фишер выдвинул гипотезу «сексуальных сыновей». Суть гипотезы в том, что формирование признаков, связанных с половым диморфизмом (как в случае с окраской тела и длиной хвоста у павлинов), происходит вследствие пара, тельной эволюции самого признака у одного пола, являющегося его носителем, и влечения к нему — у другого пола. Предположим, что вначале длинный хвост являлся индикатором здоровья самца, и что ни одному хищнику не удавалось оборвать этот хвост. Самки, которые выбирали в партнеры самцов с таким признаком, оставили более жизнеспособных сыновей, чем те, которые спаривались с короткохвостыми самцами.
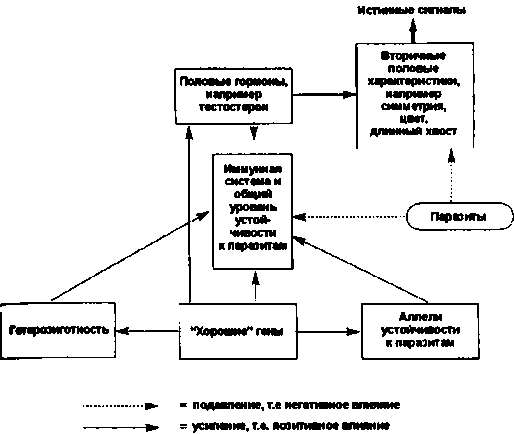
Рис. 3.7. Взаимосвязь между устойчивостью к паразитам, выраженностью половых характеристик, иммунной устойчивостью и избирательностью самок. (Дано по Cartwright, 2000).
Со временем отбор стал благоприятствовать самкам, выбирающим длиннохвостых самцов. Но это означает, что длинный хвост стал выгодным признаком сам по себе, ибо самцы с такими хвостами получали доступ к большему числу партнерш. Самки, которые выбирали длиннохвостых самцов, оставляли не только более здоровых, но и более репродуктивно успешных сыновей, потому что длиннохвостые сыновья оказывались более привлекательными для окружающих самок. Таким путем происходила коэволюция внешности самцов и вкусовых предпочтений у самок. Процесс, благодаря которому происходит формирование вторичного полового признака и закрепляется его предпочтение у представителей противоположного пола, и получил название убегающего отбора.
Конечно, самки не сознательно принимают решение — кого им следует выбирать, ими движет относительно простая поведенческая программа: правило «делай так, как твои товарки». Если представить, что какая-нибудь упрямая курочка проявит оригинальность и пойдет наперекор «общественному мнению», выбрав короткохвостого мужа, то она окажется в проигрыше. Ведь ее сыновья не будут иметь успеха, а ее гены с меньшей вероятностью перейдут в следующее поколение. Короткохвостому и тускло окрашенному самцу легче прятаться от хищников и летать на большие расстояния — но что в этом толку, если ни одна самка не выберет его в мужья и отцы? Так что проявлять индивидуальность в вопросах моды на самцов-производителей у павлинов, райских птиц или павианов гамадрилов — занятие исключительно опасное. В настоящее время доказано: во многих случаях половой отбор идет именно таким путем.
Еще одну оригинальную модель отбора мужских признаков, которые не дают преимущества при выживании, но предпочитаются самками, предложил известный этолог А. Захави. базируясь на собственных наблюдениях за сообществами птичек говорушек. Она известна под названием «принцип гандикапа»: самки выбирают самцов с длинными хвостами, волочащимися по земле, именно потому, что выжить с таким хвостом (или ярким, броским оперением, или громким голосом, привлекающим хищников...) действительно очень нелегко — значит, самец, которому это удается, и есть самый сильный, ловкий и быстрый. На первый взгляд кажется, что эта модель вес переворачивает с ног на голову, но и в ее пользу есть веские доводы. Мы не будем сейчас на них останавливаться. Просто примем во внимание, что половой отбор — одно из самых удивительных явлений в природе, и если у животных оно еще недостаточно изучено, что говорить о человеке?
Исследователи полового отбора давно обратили внимание на тот факт, что самцы у большинства видов птиц и животных значительно не избирательны по сравнению с самками. Они спариваются со многими партнершами, и в один сезон размножения преуспевающий самец может оставить потомства много больше, чем самая преуспевающая самка. Такая ситуация наблюдается у райских птиц, страусов, морских львов, оленей и многих видов обезьян — макак, павианов, лангуров, носачей, ревунов, орангутанов, горилл, шимпанзе. Да и человек не исключение из этого правила (правда, сезонность размножения у него отсутствует, впрочем, у большинства перечисленных выше обезьян — ее тоже не наблюдается).
Отбор среди самок идет не по принципу «кто красивее» (здесь мы не касаемся вопросов женской привлекательности в человеческом обществе), а чаще всего по критерию «хороших материнских качеств» и хорошего здоровья. Чем более заботливой и умелой матерью окажется конкретная самка — тем больше ее детенышей выживут и достигнут половой зрелости. Чем здоровее самка — тем выше вероятность выживания ее самой и ее потомства.
Третья гипотеза, направленная на объяснение феномена межполового отбора получила название гипотезы сенсорной эксплуатации. В рамках этой концепции половые предпочтения эволюционировали как побочный продукт уже существующего смещения в сенсорных способностях у данного вида животных. Представитель избираемого пола в этом случае как бы использует предпочтения во вкусах избирающего пола. К примеру, самцы могут эволюционировать в сторону броской расцветки, которая легко воспринимается зрительными органами самок, или могут специализироваться на песне, которая наиболее оптимально фиксируется органами слуха потенциальных партнерш, или специализироваться на секреции запаха, оказывающего максимальное положительное воздействие на обонятельные органы самки. Заметим, что появление таких новых качеств целиком и полностью продукт полового отбора, так как они обеспечивают самцам преимущества в спаривании, но вовсе не дают никаких преимуществ в выживании (скорее наоборот, затрудняют выживание, как уже говорилось выше). Однако, что касается предпочтений со стороны самок, то они являют собой преадаптацию, и, по-видимому, эволюционируют в силу иных экологических или поведенческих причин, а не являются следствием направленной половой избирательности.
Целый ряд феноменов полового поведения легче всего объясним именно с применением гипотезы сенсорной эксплуатации. Возьмем, к примеру, явление полового каннибализма у насекомых. Как известно, у многих хищных насекомых (богомолов, пауков) самка по размерам существенно превосходит самца, и важнейшим моментом ее успешной репродукции выступает необходимый объем запасенных энергетических ресурсов (Рис. 3.8). В этих условиях брачный партнер зачастую служит дополнительным источником ее пропитания. Как показывают наблюдения энтомологов, поведение такого типа может быть специфической адаптацией (крайним вариантом самопожертвования) самцов, а может просто оказываться побочным продуктом какой-то другой жизненно важной стратегии. Так, у одного из видов пауков, охотящихся под водой, Dolomedesfimbriatus, самка поедает самца во время копуляции, но эта модель поведения не может считаться адаптивной. Скорее, ее следует считать побочным продуктом отбора на оптимальное пищевое поведение особей женского пола, обеспечивающего хозяйкам повышенную плодовитость. Как показали Арнквист и Хенриксон (Amqvist, Henriksson, 1997), количество яиц, отложенное самкой этого вида, зависит от размеров самки в ее конечном половозрелом состоянии, а размеры эти непосредственно зависят от способности и желания самки нападать на любую жертву без разбору. Чем ниже избирательность самки в выборе добычи, тем более крупных размеров она достигает к репродуктивному возрасту. Доказано, что от подобного каннибализма не получают никакой выгоды ни один из полов. Самки, практикующие каннибальскую стратегию, не повышают при этом свою плодовитость. Скорее, их плодовитость даже снижается, поскольку часто случается так, что самка съедает самца раньше, чем он успевает ввести ей в половые пути всю имеющуюся у него сперму. Не получает никакой видимой выгоды при этом и самец.
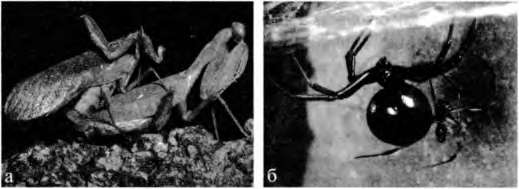
Рис. 3.8. Половой диморфизм по размерам тела у беспозвоночных животных. Самки часто много крупнее самцов: а) богомолы; б) пауки черная вдова — огромная самка и карликовый самец. (Дано по Нири, 1997).
Явление неадекватного полового каннибализма у паука Dolomedes flmbriatus находится в полном контрасте с адаптивным половым каннибализмов у других видов паукообразных. Например, доказано, что у садового паука вида Araneus diadematus масса тела самки существенно возрастает в результате поедания полового партнера, а, следовательно, повышается и ее плодовитость. Пауки самцы получают прямую выгоду от такого крайнего варианта самопожертвования, так как при этом они оставляют больше потомства. У австралийского красноспинного паука Latrodectus hasselti выгода от самопожертвования носит еще более прямой характер. Самцы, съеденные супругами во время полового акта, оплодотворяют примерно в два раза больше яиц (следовательно, оставляют больше потомков), так как дольше находятся в коитусе с самкой, пока та их поедает, чем те, кто ретируется с поля сексуальной битвы живыми и невредимыми.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Охрана партнера
Охрана партнера Поскольку собственническое поведение столь широко распространено в природе, специалисты дали ему название: охрана партнера. (38) По их мнению, это стремление к сексуальной эксклюзивности является одним из центральных аспектов ухаживания у множества
Мозг в поисках партнера
Мозг в поисках партнера Почему мы, люди, так сильно отличаемся друг от друга?Мои мысли на эту тему вытекают из гениальной идеи Чарльза Дарвина о половом отборе.Дарвину не давало покоя многообразие украшений, созданных природой. (77) Это и малиновые «воротники», и синие
ВЫБОР ПАРТНЕРА: КТО КОГО ВЫБИРАЕТ?
ВЫБОР ПАРТНЕРА: КТО КОГО ВЫБИРАЕТ? Вернемся к описанию репродуктивного цикла. Во время демонстраций происходит выбор репродуктивного партнера, то есть это — индивидуализация объекта, на который направлено дальнейшее поведение.Инициатива выбора у противоположных полов
8.1. НАРУШЕНИЯ ПОЛОВОГО ЦИКЛА
8.1. НАРУШЕНИЯ ПОЛОВОГО ЦИКЛА Нарушения полового цикла возникают в результате самых раз-личных расстройств в системе гипоталамус — гипофиз — яичники и могут служить признаком некоторых генитальных и экстрагени-тальных заболеваний. Встречаются достаточно часто и
9. ВЫБОР ОПТИМАЛЬНОГО ВРЕМЕНИ ДЛЯ ИСКУССТВЕННОГО ОСЕМЕНЕНИЯ
9. ВЫБОР ОПТИМАЛЬНОГО ВРЕМЕНИ ДЛЯ ИСКУССТВЕННОГО ОСЕМЕНЕНИЯ Причиной бесплодия часто бывает неправильный выбор времени искус венного осеменения.По клиническим проявлениям. От появления у суки первых коровянистых выделений искусственное осеменение проводят на 11 — 14
Ритуализация полового поведения
Ритуализация полового поведения Поведение играет колоссальную, если не сказать ведущую, роль, как для самого процесса размножения, так и для полового отбора.Отбор идет по пути жесткой выбраковки партнеров, которые ведут себя не совсем «правильно» или не достаточно
Формирование полового поведения
Формирование полового поведения Для формирования нормального полового и родительского поведения собак обоего пола очень важны условия, в которых растет щенок. Многочисленные исследования, проведенные на животных разных видов, показали, что самки, выкормленные
Период полового созревания
Период полового созревания Пятый период (после 7 месяцев) — период полового созревания. К этому моменту рост щенка в основном заканчивается. Завершается смена молочных зубов на постоянные. Совершенствуются и развиваются регуляторные механизмы и функциональные системы.
Выбор партнера: кто кого выбирает?
Выбор партнера: кто кого выбирает? Вернемся к описанию репродуктивного цикла. Во время демонстрации происходят выбор репродуктивного партнера, индивидуализация объекта, на который направлено дальнейшее поведение. Инициатива выбора у противоположных полов всегда
Выбор партнера
Выбор партнера Склонность женских особей к накоплению проявляется не только в их физиологии, но и в поведении. Накопленный ресурс не должен быть растрачен попусту. Этим принципом определяется выбор женщинами репродуктивного партнера.То, что женщины выбирают мужчин, а не
Выбор брачного партнера
Выбор брачного партнера В моногамном обществе действуют специфические закономерности, знание которых может (по крайней мере, теоретически) помочь женщине угадать, какие мужчины будут лучшими мужьями и отцами. По американским моделям четко видно, что вероятность того,
Выбор постоянного и временного партнера: ассортативность и самооценка
Выбор постоянного и временного партнера: ассортативность и самооценка Как показано выше, к настоящему времени в научной литературе накоплена масса сведений о факторах, определяющих внешнюю привлекательность мужчин и женщин. Однако реальные взаимоотношения партнеров
Выбор постоянного партнера: внешнее сходство партнеров с родителями противоположного пола
Выбор постоянного партнера: внешнее сходство партнеров с родителями противоположного пола В повседневной жизни часто можно встретиться с представлениями о том, что супруги бывают похожи друг на друга. Исследования антропологов и психологов как будто бы подтверждают
Выбор партнера: взаимность чувств самое главное
Выбор партнера: взаимность чувств самое главное Попробуем теперь ответить на вечный вопрос: что главное в любимом человеке? Совсем недавно мы (М. Бутовская, О. Смирнов, 2003) опубликовали результаты проведенного опроса молодых русских москвичей (от 18 до 28 лет): какие признаки
Выбор партнера, доминирование и репродуктивный успех
Выбор партнера, доминирование и репродуктивный успех Некоторые авторы считают, что доминантные самцы (у макаков и павианов) имеют преимущество в выборе самки и чаще, чем подчиненные, спариваются с самками в эструсе. Однако преимущества нарушаются самками, которые не