Номенклатура, физико-химические свойства и функции нуклеозидов и нуклеотидов
Положение фосфатной группы в молекуле нуклеотида указывается цифрой. Например, аденозин с фосфатной группой, присоединенной к 3-му углероду рибозы, должен быть обозначен как 3?-монофосфат. Штрих после цифры ставят для того, чтобы отличить номер углерода в пуриновом или пиримидиновом основании от положения этого атома в остатке дезоксирибозы. При нумерации атомов углерода основания штрих не ставится. Нуклеотид 2?-дезоксиаденозин с фосфатным остатком при углероде-5 молекулы сахара обозначается как 2?-дезоксиаденозин-5?-монофосфат. Нуклеозиды, содержащие аденин, гуанин, цитозин, тимин и урацил, принято обозначать буквами A, Г, Ц, Т и У соответственно. Наличие буквы d (или д) перед сокращением обозначает, что углеводным компонентом нуклеозида является 2?-дезоксирибоза. Гуанозин, содержащий 2?-дезоксирибозу, может быть обозначен дГ (дезоксигуанозин), а соответствующий ему монофосфат с фосфатной группой, присоединенной к третьему атому углерода дезоксирибозы, – дГ-3?-МФ. Как правило, в тех случаях, когда фосфат присоединен к углероду-5 рибозы или дезоксирибозы, символ 5? опускается. Так, гуанозин 5?-монофосфат принято обозначать ГМФ, а 5?-монофосфат 2?-дезоксигуанозина сокращают как дГМФ. Если к углеводному остатку нуклеозида присоединены 2 или 3 остатка фосфорной кислоты используются аббревиатуры ДФ (дифосфат) и ТФ (трифосфат). Таким образом, аденозин + трифосфат с тремя фосфатными группами в 5?-положении углевода будет обозначаться АТФ. Поскольку в молекулах нуклеотидов фосфаты находятся в виде ангидридов фосфорной кислоты, т. е. в состоянии с низкой энтропией, их называют макроэргами (обладающими большим запасом потенциальной энергии). При гидролизе 1 моля АТФ до AДФ высвобождается 7,3 кКал потенциальной энергии.
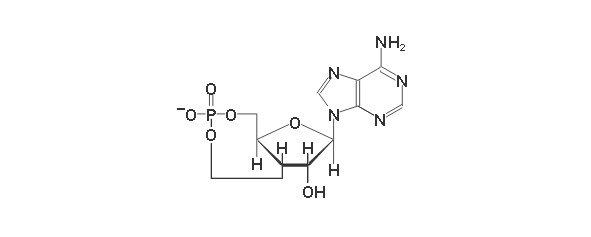
Рисунок 35. Структура цАМФ
Физико-химические свойства нуклеотидов
Так как в состав нуклеотидов входят азотистые основания, то такие свойства как таутомерия и способность поглощать в ультрафиолетовой части спектра также характерны и для нуклеотидов, причем спектры поглощения азотистых оснований и содержащих эти основания нуклеотидов сходны. Наличие сахара и остатков фосфорной кислоты делает их более гидрофильными чем азотистые основания. Все нуклеотиды являются кислотами, так как содержат остатки фосфорной кислоты.
Функции природных нуклеотидов
Нуклеотиды являются мономерами нуклеиновых кислот (РНК, ДНК). В состав ДНК входят дезоксирибонуклеотидфосфаты – производные аденина, тимина, гуанина и цитозина. Также некоторые молекулы гуанина и цитозина в составе ДНК метилированы, то есть содержат метильную группу. Как основные мономеры в состав РНК входят рибонуклеотидфосфаты – производные аденина, урацила, гуанина и цитозина. Также в состав РНК входят нуклеотиды, содержащие различные минорные азотистые основания, например ксантин, гипоксантин, дигидроуридин и др.
Нуклеотиды являются мономерами коферментов (НАД, НАДФ, ФАД, ко-энзим А, метионин-аденозин). В составе коферементов они участвуют в ферментативных реакциях. Более подробно эта функция будет рассмотрена ниже.
Энергетическая (АТФ). АТФ выполняет функцию основного внутриклеточного переносчика свободной энергии. Концентрация наиболее распространенного свободного нуклеотида в клетках млекопитающих – АТФ – составляет около 1 ммоль/л.
Сигнальная (цГМФ, цАМФ) (Рисунок 35). Циклический AMФ (3?-, 5?-аденозинмонофосфат, цАМФ) – медиатор различных внеклеточных сигналов в клетках животных – образуется из АТФ в результате реакции, катализируемой аденилатциклазой. Активность аденилатциклазы регулируется комплексом взаимодействий, многие из которых инициируются через рецепторы гормонов. Внутриклеточная концентрация цАМФ (около 1 мкмоль/л) на 3 порядка ниже концентрации ATФ. Циклический цГМФ (3?-, 5?-гуанозинмонофосфат, цГМФ) служит внутриклеточным проводником внеклеточных сигналов. В некоторых случаях цГМФ выступает в роли антагониста цАМФ. цГМФ образуется из ГТФ под действием гуанилатциклазы – фермента, имеющего много общего с аденилатциклазой. Гуанилатциклаза, как и аденилатциклаза, регулируется различными эффекторами, в том числе и гормонами. Как и цАМФ, цГМФ гидролизуется фосфодиэстеразой до соответствующего 5?-монофосфата.
Регуляторная (ГТФ). Активность группы белков (G-белков), выполняющих в основном регуляторную функцию, зависит от того: какой нуклеотид они связывают. В неактивной форме эти белки связывают ГДФ, при активации белка происходит замена ГДФ на ГТФ. При выполнении своей функции белок гидролизует ГТФ до ГДФ и фосфата, выделившаяся, энергия затрачивается на функционирование белка.
Активация при метаболизме липидов и моносахаридов (УТФ, СТФ). Производные урациловых нуклеотидов участвуют в качестве активирующих агентов в реакциях метаболизма гексоз и полимеризации углеводов, в частности при биосинтезе крахмала и олигосахаридных фрагментов гликопротеинов и протеогликанов. Субстратами в этих реакциях являются уридин-дифосфатсахара. Например, уридиндифосфатглюкоза служит предшественником гликогена. Также превращение глюкозы в галактозу, глюкуроновую кислоту или другие производные моносахаридов происходит в виде коньюгата с УДФ. СТР необходим для биосинтеза некоторых фосфоглицеридов в тканях животных. Реакции с участием церамида и ЦДФ-холина приводят к образованию сфингомиелина и других замещенных сфингозинов.
Участие в дезактивации различных спиртов и фенолов (УДФ-глюкуроновая кислота). Уридиндифосфатглюкуроновая кислота – выполняет функцию «активного» глюкуронида в реакциях конъюгирования, например, при образовании глюкуронида билирубина.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.