Глава 2 Человекообезьяны: грацильные австралопитеки
Когда весной 1924 года южноафриканский преподаватель Раймонд Дарт вскрывал присланные ему ящики с окаменелостями, он уже ждал чего-то необычного. Уже два года он собирал древние кости, которые шахтеры иногда случайно находили в разных рудниках Южной Африки. Но вряд ли Р. Дарт рассчитывал на то открытие, которое он сделал, вскрыв второй ящик. Везение палеонтолога – особого рода, оно связано с долгим трудом и тщательными поисками, часто растягивающимися на долгие годы. Эжен Дюбуа искал своего питекантропа тоже два года, Луис Лики работал в Олдувайском ущелье до первых важных находок 28 лет, а сколько археологов и антропологов так и не находят “свою” сенсацию! Так что череп, который обнаружил Р. Дарт поверх остальных окаменелостей, можно считать поистине подарком судьбы и наградой за неустанные изыскания. Находка эта тем более удивительна, что за все последующие годы в руднике Таунг, откуда были присланы ящики, не было найдено ни одной новой кости австралопитека. Впрочем, когда Р. Дарт осматривал удивительный череп, слова “австралопитек” еще не существовало, его придумал и ввел в научный оборот сам Р. Дарт, опубликовав пятистраничную статью в журнале Nature в следующем году (Dart, 1925). Многие ученые отнеслись тогда к открытию, мягко говоря, осторожно, такова уж особенность научного мышления. Следующие десять лет не принесли подтверждений значимости новооткрытого вида древних приматов. Но потом за поиски австралопитеков в других южноафриканских рудниках взялся зоолог Роберт Брум, и с тех пор окаменелости посыпались как из рога изобилия. Сокровищница разверзлась, и сенсационные находки с той поры и по сей день не прекращают радовать антропологов. Сейчас известно много сотен окаменевших останков австралопитеков из разных частей Африки, и мы знаем о них гораздо больше, чем о большинстве других древних животных.

Рис. 5. Таунг.
Животных или людей? Австралопитеки – кто они, эти странные существа с телом человека и головой обезьяны? “Южные обезьяны”, как гласит их название (которое, кстати, никак не связано с Австралией, где никогда не было обезьян), или обезьянолюди, то самое пресловутое “недостающее звено”? Были ли они нашими предками или исчезли, не оставив потомков? А может, у разных австралопитеков была разная судьба?
Кстати, о переводах…
Мелкие виды австралопитеков, известные лучше всего – прежде всего Australopithecus afarensis и Australopithecus africanus, – суммарно называются грацильными. Такое название дано им за небольшие размеры сравнительно с массивными австралопитеками – парантропами. Эти термины – “грацильный” и “массивный” – глубоко укоренились в антропологической литературе. Тем более забавно, когда журналисты пытаются их перевести: так появились “изящные”, “стройные”, “субтильные”, “деликатные”, “хрупкие”, а также “мощные” и “могучие” австралопитеки. По таким лингвистическим мелочам, как и в любой специфической деятельности, легко распознать профессионала и дилетанта.
Южноафриканские австралопитеки стали первыми известными науке, но они не были древнейшими из открытых на данный момент. Грацильные австралопитеки Восточной Африки имеют более солидный возраст. И конечно, самые известные и изученные из них…
Афарские австралопитеки Australopithecus afarensis – от них сохранились десятки отличных окаменелостей, а если считать изолированные зубы – то и сотни. Подавляющая часть находок сделана в Эфиопии, в пустыне Афар, в местности Хадар, но наиболее древние обнаружены в Ворансо-Милле, другие – в Лаэтоли в Танзании, Южном Таквиле в Кении. и самые молодые – в Омо в Эфиопии. Есть и другие местонахождения: Лотагам, Мака, Дикика, Кантис. Приблизительно с 4 млн лет назад палеонтологическая летопись становится достаточно полной и последовательной. Суммарно вид афарских австралопитеков существовал примерно с 3,6–3,8 до 2,9 млн лет назад, а возможно – даже до 2,3 млн лет назад.
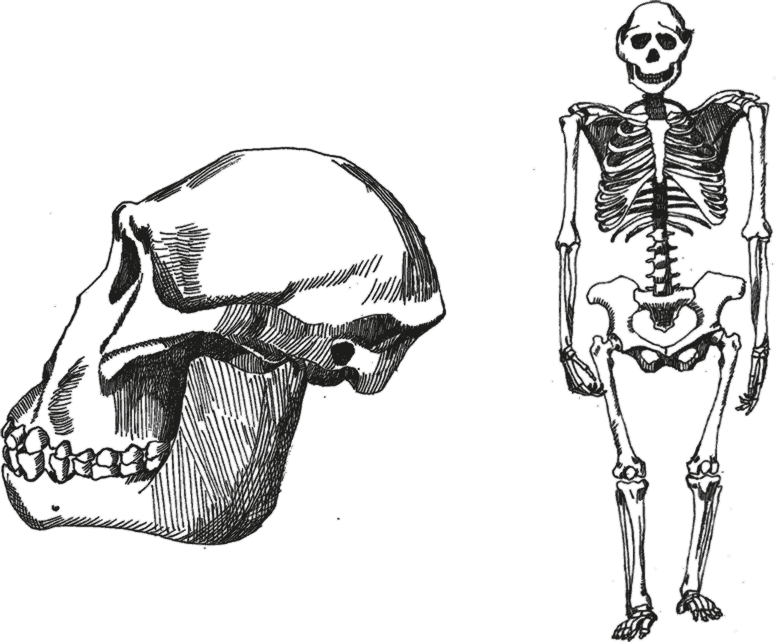
Рис. 6. Череп и скелет Australopithecus afarensis.
Одно из древнейших местонахождений афарских австралопитеков – танзанийское Лаэтоли. Более того, нижняя челюсть LH 4, найденная тут в 1974 году, послужила голотипом – образцом для описания – всего вида (Johanson et al., 1978). Однако самой необычной и доселе непревзойденной находкой стала цепочка следов. 3,66 млн лет назад в Лаэтоли извергся вулкан. Пепел завалил все вокруг, после чего прошел дождь и равнина покрылась слоем грязи. Пробегавшие по ней зверюшки – от зайцев до саблезубых тигров и слонов – оставили пересекающиеся следовые дорожки. Оставили свое наследие и двуногие – рядом с ним нет никаких отпечатков рук. Грязь высохла, а вулкан после этого извергся еще разок, засыпав все сверху новым шлаком. И так следы ждали своего часа прямо под почвой танзанийской саванны вплоть до XX века, чтобы в 1978 году предстать перед изумленными взорами своих далеких потомков.
Сохранилось три цепочки следов австралопитеков. Первая – A – совсем короткая и редко вспоминаемая, к тому же ее следы странно широки, так что есть подозрение, что ее оставил вообще медведь (Tuttle, 2008). Неясно, правда, зачем ему было идти по саванне на двух ногах, да и костей медведей в Лаэтоли не найдено, но в принципе Agriotherium обитал в Эфиопии и Южной Африке.
Две главные цепочки – G – идут рядышком, при этом линия крупных следов – это на самом деле две дорожки, просто второй австралопитек ступал след в след первому. Отпечатки ног мелкого расположены слева от крупного. Поэтому часто упоминается, что это была семья – папа, мама и ребенок. Однако, судя по сохранности и различиям в четкости, следы, скорее всего, были оставлены в разное время, мелкий прошел тогда, когда грязь уже подсыхала, отчего его отпечатки оказались намного четче.
Вокруг следов с тех пор идет дискуссия: одни исследователи считают, что их оставили австралопитеки, другие указывают на обезьян или предполагают неведомых зверей, аки в Лукоморье. Дескать, известные кости стоп афарских австралопитеков неидеально соответствуют следам. Тут надо уточнить, что кости стоп найдены в Хадаре – в Эфиопии, а не в Кении – и они на полторы-две сотни тысяч лет моложе. Все же несомненно, что у существ, прошлепавших по вулканической грязи Лаэтоли, были развиты своды стопы, а большой палец был почти полностью прижат к остальным. На самом деле немножко он все же от прочих отстоял, но и у современных людей, всю жизнь ходящих босиком, большой палец может быть отведен довольно далеко, даже больше, чем у ходоков Лаэтоли. У шимпанзе следы совсем иные, скорее похожие на отпечатки нашей руки. Другое дело, что пальцы лаэтольцев были заметно завернуты внутрь, чего у современных людей практически никогда не бывает, то есть ноги были слегка “косопалые”, хотя и не косолапые.
Следы мелкие, рост существ был всего 1, 1,2 и 1,4 м – в самый раз австралопитечий. Несмотря на апокалиптическое извержение, разразившееся у них над головой, скорость существ была вполне прогулочной, около пяти километров в час. Основные показатели ходьбы в принципе не отличаются от современных, только шаг был слегка семенящий. С другой стороны, когда неподалеку из горы пышет пламя и валит дым, еще неизвестно, какими бы шагами пошли современные люди…
Кадануумуу – австралопитек, не похожий на человека
Ворансо-Милле – замечательная местность в Эфиопии, в которой нашли уже много интереснейших находок. Самой сенсационной из них стал скелет австралопитека KSD – VP-1/1 возрастом 3,58–3,6 млн лет назад, обнаруженный в 2005 г. в локальном местонахождении Корси-Дора (Haile-Selassie et al., 2010a). Скелет был отнесен к виду Australopithecus afarensis, хотя находки зубов из соседних местонахождений с такими же датировками имеют еще много черт предыдущего вида – A. anamensis. К сожалению, череп не сохранился, зато скелет уникален сразу по трем причинам: во-первых, он зело древний, во-вторых, весьма сохранный, в-третьих, большой. Не зря его назвали Кадануумуу – Большой Человек. И было за что – рост гиганта достигал аж 1,52–1,68 м! На фоне метровой Люси он выглядел просто каланчой. Патриархи афарских австралопитеков могли бы говаривать своей малышне, поглаживая седые бакенбарды: “Дааа… Были австралопитеки в былые времена! Богатыри, не мы…” Впрочем, они еще не умели говаривать…
С другой стороны, скелет KSD – VP-1/1 по размерам вполне вписывается в масштабы изменчивости афаренсисов, судя по многочисленным известным находкам из Хадара: например, известны останки крупной особи AL 438–1. Люси, кстати, была очень мала даже для австралопитеков. В изменчивости вида, существовавшего более миллиона лет, нет ничего удивительного. Если взять, например, пигмеев, эскимосов и масаев – у них тоже рост и пропорции будут весьма различаться, притом что они относятся к одному виду и существуют одновременно, а между Большим Человеком и Люси – несколько сотен тысяч лет.
Кроме исполинского роста, Кадануумуу удивил исследователей прогрессивностью своих конечностей. В строении ключицы, лопатки, таза, костей рук и ног они отметили многочисленные продвинутые черты, сближающие древнейшего грацильного австралопитека с людьми и отличающие от африканских человекообразных обезьян. Посему некоторые журналисты, а вслед за ними креационисты, как обычно не вникнув в суть происходящего, растиражировали байку про скелет “совсем современного человека невиданной древности”.
Тут стоит сделать несколько ученых замечаний.
Отличия австралопитеков от Homo в посткраниальном скелете довольно многочисленны и ярко выражены у KSD – VP-1/1.
Ребра Большого Человека имеют скорее человеческий, нежели обезьяний изгиб, однако второе ребро KSD – VP-1/1 намного массивнее, чем у человека, хотя размер тела у австралопитека меньше. Ключица KSD – VP-1/1 крайне массивная (выходит за рамки изменчивости человека) и, вероятно, была относительно очень длинной (в верхних пределах изменчивости человека). Примечательно, что по массивности шимпанзе и даже гориллы больше похожи на человека, чем KSD – VP-1/1. Соотношение надостной и подостной ямок лопатки у KSD – VP-1/1, по всей вероятности, было больше похоже на шимпанзе и гориллу, чем на человека, хотя сохранность лопатки KSD – VP-1/1 очень плохая. Наклон суставной впадины лопатки относительно латерального края (если линия проходит через центр суставной впадины) находится в нижних пределах человеческой изменчивости и в верхних – гориллы, хотя и выходит за рамки изменчивости шимпанзе (у Люси входит в рамки шимпанзе и выходит за рамки человека). Другие черты Кадануумуу более гоминидны: наклон оси лопатки относительно латерального края (если линия проходит через низ суставной впадины) вполне вписывается в изменчивость человека и далеко выходит за рамки изменчивости шимпанзе и гориллы.
На плечевой кости мыщелок ориентирован крайне горизонтально, медиальный надмыщелок очень велик, блок слабо выступает вниз, головка мыщелка относительно очень большая – все эти признаки отличаются от человеческого варианта. На локтевой кости крайне нетипично для человека мощное развитие локтевого отростка в ширину. Угол локтевой вырезки (линия проходит через вершины локтевого и венечного отростков) к задней стороне локтевой кости намного ниже индивидуального минимума человека, Люси и даже ардипитека (хотя у Люси и ардипитека тоже ниже человеческого минимума). Показательно, что шимпанзе почти не отличается от человека, а горилла имеет в среднем больший угол, но значения тоже вполовину пересекаются с человеком.
Ряд показателей локтевой и плечевой кости, учтенных относительно длины большой берцовой кости, вписывается в верхние пределы изменчивости человека, но почти совпадает с Люси и уклоняется в сторону шимпанзе и гориллы.
Крыло подвздошной кости у афарских австралопитеков было заметно сильнее наклонено, чем у человека. Этот признак – увеличенная передне-задняя длина и сильная развернутость подвздошного гребня – хорошо выражен на скелете KSD – VP-1/1, хотя его таз и фрагментарен (более фрагментарен, чем хорошо известный таз Люси). Связанный с предыдущим признак – крайне широкие боковые части крестца; он выражен у других австралопитеков и у KSD – VP-1/1, причем последний выходит за рамки изменчивости современных людей, хотя и оказывается “самым очеловеченным” из австралопитеков и даже “более человечным”, чем человек из Гоны (правда, шимпанзе трансгрессируют с человеком, а австралопитеки – нет!).
Классический признак таза австралопитеков (известного, к слову сказать, не только по Люси, но и по ряду других находок, в частности Sts 14 и Stw 431 в Южной Африке) – увеличенное расстояние между вертлужной впадиной и седалищным бугром (лучше определять относительный размер, так как австралопитеки были мелкие): у KSD – VP-1/1 оно даже больше, чем у других австралопитеков, далеко побивает индивидуальный рекорд человека и занимает промежуточное положение между человеком и группой ардипитек – шимпанзе – горилла. Другой австралопитековый признак – большое расстояние между задними верхней и нижней подвздошными остями – у KSD – VP-1/1 выражен очень типично, совсем непохоже на человека. Гребень подвздошной кости у KSD – VP-1/1 и других австралопитеков имеет менее выраженный сигмовидный изгиб, чем у человека. Передняя ягодичная линия подвздошной кости у KSD – VP-1/1 смещена заметно дальше вперед, чем у человека. У KSD – VP-1/1 узкие и вытянутые ушковидные поверхности подвздошной кости (у человека намного более короткие и широкие), умеренные размеры вертлужной впадины (у человека намного больше) – в общем, отличий от человека хватает.
Форма мыщелков бедренной кости при взгляде сбоку у KSD – VP-1/1 хотя и эллиптическая, но по показателям формы занимает промежуточное положение между человеком и шимпанзе, как и у прочих австралопитеков. У человека межмыщелковая впадина гораздо крупнее (и длиннее, и шире), а мыщелки намного длиннее и относительно намного уже и вытянутее. Даже в индивидуальных случаях такой формы, как у KSD – VP-1/1, у людей не бывает. Нижний конец большой берцовой кости у KSD – VP-1/1 сильно поврежден, но видно, что он больше похож на человеческий, чем на шимпанзиный. Однако у людей такой формы суставной поверхности и такого расширения нижнего эпифиза большой берцовой не бывает. Кроме того, диафиз большой берцовой KSD – VP-1/1 хотя и уплощен с боков, но скруглен сильнее, чем обычно у людей.
В итоге утверждать сапиентность скелета KSD – VP-1/1 может только человек, совсем незнакомый с эволюционной морфологией скелета и, кроме того, намеренно искажающий факты. Вообще, если бы кто нашел скелет сапиенса с датировкой под 3 млн лет – это была бы такая сенсация, что ни один антрополог не удержался бы от пафосной публикации во всех ведущих журналах! Антропологи тоже любят славу. Глупо думать, что антропологи способны намеренно скрывать факт “слишком раннего” существования сапиенсов. Наоборот, стремятся всеми силами запихать в сапиенсы находки с датировками уже под 200 тыс. лет и даже больше! Ндуту, Нгалоба, Омо, Херто – тому лучшие примеры. Про Схул и Кафзех я уж молчу. Со скрипом получается, а пихают…
Наиболее известны австралопитеки Хадара, особенно скелет самки, получивший прозвище Люси. В горячий полдень 30 ноября 1974 года участники американо-французской экспедиции антрополог Дональд Джохансон и палеонтолог Том Грей уже собирались заканчивать поиски окаменелостей, но решили все же пройтись по одному из местных оврагов, который уже дважды тщательно осматривали другие исследователи, и… До настоящего времени этот скелет остается самым полным по сохранности из всех австралопитековых. Его датировка – 3,18 млн лет назад. Счастливые первооткрыватели всю ночь веселились, а над Хадаром разносились звуки битловской песни “Люси в небе с брильянтами” – так и возникло имя Люси. Другой скелет из Хадара, гораздо хуже сохранившийся, – AL 438–1.
Не менее удивительна находка так называемого “Первого семейства” AL 333 – две с половиной сотни костей от семнадцати австралопитеков, лежавших на небольшом участке склона, на которые буквально села жена французского кинооператора, приехавшая в Хадар без малейшего представления об ископаемых приматах. Девять взрослых, три подростка и пять детенышей, похоже, сидели себе спокойно в каком-то овраге, когда их нежданно-негаданно накрыло наводнение от прошедшего дождя. Зря они собрались тут – не к добру это было! Для них. А для нас их беззаботность предоставила уникальный шанс изучить группу афарских австралопитеков. Правда, потом над останками поработали падальщики, так что до нас, к сожалению, дошли не семнадцать скелетов, а мешок огрызков, но лучшей коллекции все равно нигде больше нет.
В Хадаре найдены три почти целых черепа – самца AL 444–2, самки AL 822–1 и детеныша AL 333–105. Хватает и фрагментов, так что мы можем оценивать возрастную, половую и хронологическую изменчивость афаренсисов.
В 2006 году из эфиопского местонахождения Дикика с датировкой 3,3 млн лет назад описан почти целый череп и скелет маленькой трехлетней самочки (или уже девочки?) афарского австралопитека, названной дочкой Люси, или Селам. Теперь мы гораздо больше знаем о детях австралопитеков, особенностях их роста, а также пропорциях тела.
Природа австралопитеков двойственна: практически во всех костях сочетаются человеческие и обезьяньи признаки (отличные обзоры современных данных: Kimbel et Delezene, 2009; Ward et al., 2012). Череп афарских австралопитеков внешне похож на шимпанзиный: мозговая коробка вытянутая, с резко покатым лбом, не слишком большим надбровьем, зато со здоровенными гребнями для прикрепления жевательной мускулатуры, в том числе сагиттальным гребнем у самцов, с мощными, развернутыми в стороны скуловыми дугами; лицевой отдел большой, сильно уплощенный, а при взгляде сбоку даже вдавленный; верхняя челюсть выступает вперед, нижняя челюсть массивная, с огромной восходящей ветвью. Для афарских австралопитеков типична не слишком сильно, но все же развитая заклыковая мегадонтия – увеличение размеров моляров относительно челюсти. Пройдет еще миллион лет, и эта особенность выльется в появление парантропов с огромными челюстями и зубами. Слава изменчивости – не все австралопитеки ступили на этот заманчивый, но тупиковый путь; самые мелкозубые, не поддавшиеся на сладкую приманку вегетарианства, стали людьми.
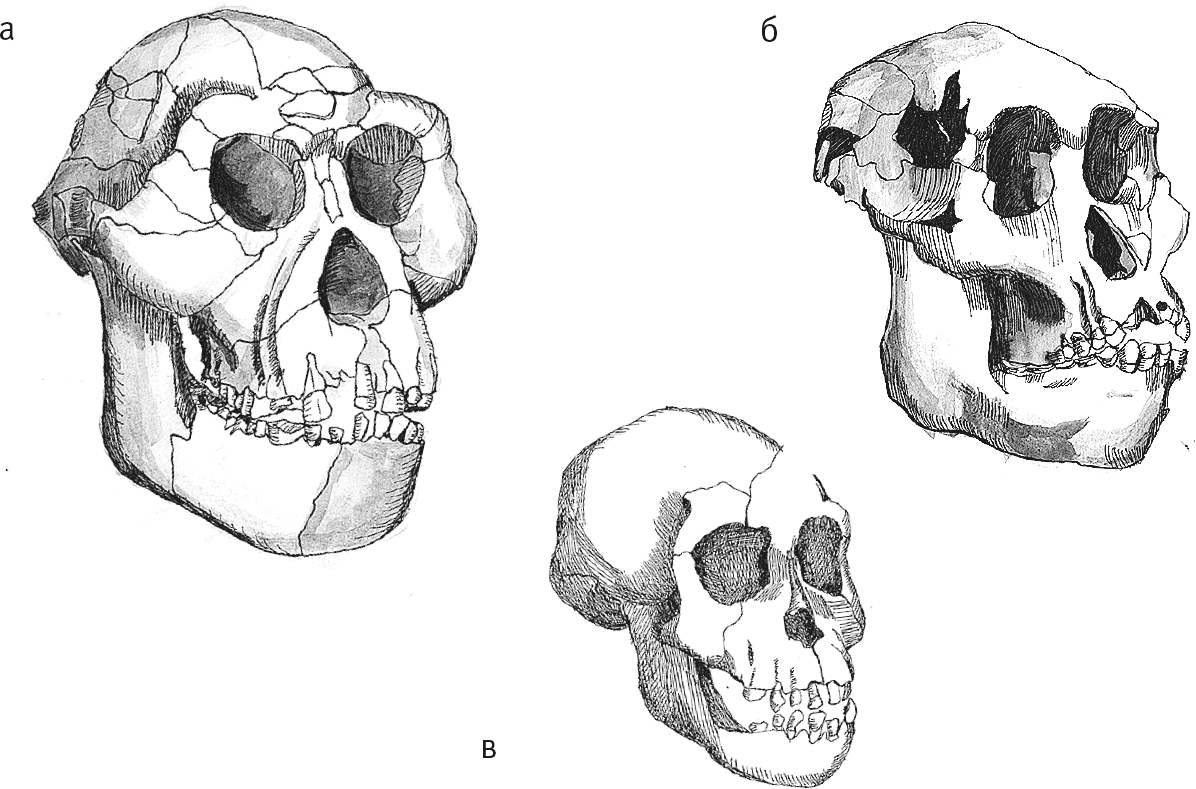
Рис. 7. Черепа самца AL 444–2 (а), самки AL 822–1 (б) и детеныша Дикика (в).
При всей обезьяноподобности афарские австралопитеки имели два ключевых человеческих признака черепа: большое затылочное отверстие у них было явно смещено ближе к середине основания черепа, чем у обезьян, а клыки хотя и намного крупнее человеческих, все же не слишком выступали за линию прочих зубов. Диастема между клыком и премоляром на нижней челюсти обычно либо была маленькой, либо вообще отсутствовала, а первый нижний премоляр был уже почти симметричен, в отличие от ранних австралопитеков.
Очевидно, у австралопитеков с их маленькими клыками взаимоотношения в группах строились не на демонстрационной агрессии, а защищались они от хищников с помощью чего-то еще, вероятно используя необработанные палки и камни. По всей видимости, здесь лежат истоки богатых возможностей человеческого общения – жестикуляции, выразительной мимики и, конечно же, речи. Вместе с тем и способность к использованию орудий труда стала следствием сочетания незащищенности и двуногости австралопитеков. Учитывая, что руки австралопитеков были “развязаны” прямохождением, способности к использованию орудий у них наверняка были развиты гораздо лучше, чем у современных шимпанзе. Впрочем, изготавливать каменные орудия они еще точно не умели, да и руки их не были к этому приспособлены, а посему нам остается только строить предположения на этот счет.
Кстати, об умелых ручках…
Как узнать о трудовой деятельности, если она предполагает не изготовление орудий труда, а лишь использование подходящих природных объектов – палок, костей и камней? Конечно, по следам этой деятельности! Именно этим путем пошли исследователи эфиопского местонахождения Дикика. Пересмотрев несметное количество обломков костей, они таки нашли, что искали: в слое с датировкой 3,4 млн лет назад обнаружились фрагменты правого ребра животного размером с корову и бедренной кости животного размером с козу со следами каменных орудий (McPherron et al., 2010). Царапины были тщательно исследованы – это не следы зубов хищников и не природные повреждения. Выходит, афарские австралопитеки могли периодически пользоваться подходящими острыми камнями, чтобы срезать мясо с костей? Неужели у нас в руках следы первой трудовой деятельности – настолько дремучей, что для нее и орудий-то еще не было? Конечно, не могли не найтись критики, показавшие, что царапины на костях из Дикики с наибольшей вероятностью имеют-таки естественное происхождение (Dom?nguez-Rodrigo et al, 2010). Дискуссия пока продолжается. Однако с чем согласны все археологи: афарские австралопитеки не умели намеренно обрабатывать камни и делать из них орудия. Тысячи тонн просмотренной породы за многие годы исследований – лучшее тому доказательство.
Объем мозга афарских австралопитеков не очень сильно отличался от типичного для шимпанзе – примерно 350–550 см?. Но все же в пересчете на массу тела австралопитеки оказываются более мозговитыми. Впрочем, форма австралопитекового мозга крайне мало отличается от обезьяньего.
Особенности посткраниального скелета позволяют без всякого сомнения говорить, что афарские австралопитеки были уже полностью прямоходящими. Это очевидно следует из особенностей строения таза, коленного и голеностопного суставов, стопы. То есть ниже головы это были уже почти совсем люди. Впрочем, почти – не значит совсем. Строго говоря, человечность в скелете афаренсисов, да и любых австралопитеков убывает снизу вверх: тогда как стопа, ноги и таз почти людские, руки и особенно голова – почти обезьяньи. Другими же словами – мы очеловечивались от ног к голове, можно сказать – от матушки-земли (какой простор открывается тут для псевдофилософских измышлений!).
Удивительно много понгидных черт сохранялось в строении позвоночника (хотя крестец весьма продвинут – прямохождение обязывает). Руки же больше прочих частей тела отставали в своем прогрессе. Так, лопатка Люси весьма понгидна, а у детеныша из Дикики больше всего напоминает лопатку гориллы, хотя и уклоняется в человеческую сторону. Руки афарских австралопитеков были заметно удлинены, хотя и не в такой степени, как у ардипитеков. Хотя плечевая, локтевая и лучевая кости имеют и прогрессивные черты, в их строении едва ли не больше признаков человекообразных обезьян, они были мощными и хорошо приспособленными для лазания по деревьям. В развитии дорзального гребня нижнего конца лучевой кости Люси многие антропологи усматривают след приспособления к костяшкохождению; впрочем, это скорее наследие анамских предков или специализация, вообще не связанная с локомоцией. Кисть афаренсисов больше похожа на человеческую общими пропорциями и многими частностями, но обезьяна кроется в деталях ее строения – укороченности большого пальца, сильном изгибе пястных и фаланг, суженности конечных фаланг. Хватает и специфических черт вроде увеличенных головки головчатой и крючка крючковидной костей. Чего не было, да и не могло быть у афарских австралопитеков, так это трудового комплекса кисти – и в этом она очевидно примитивна.
Вот что удалось афаренсисам на славу – так это таз. Он очень широкий и очень низкий; у Люси его пропорции оказываются даже более человеческими, чем у человека, за счет аллометрических закономерностей. Кости ног однозначно похожи на наши. Особенно знаменито “колено Джохансона” – фрагменты коленного сустава AL 129–1. По нему впервые было однозначно установлено прямохождение афарских гоминид. Но с тех пор появились десятки новых костей, в том числе, например, почти целая бедренная кость AL 827–1.
Стопа афарских австралопитеков известна по массе изолированных костей, судя по которым на ней были вполне развиты продольный и поперечный своды, а плюсневые кости соединены практически неподвижно (в частности, имелся контакт между основанием третьей плюсневой и промежуточной клиновидной костью). Большой палец стопы вряд ли был сильно оттопырен, уж точно не как у шимпанзе или ардипитека.
Рост афарских австралопитеков колебался от метра до полутора – практически универсальный размер вплоть до появления Homo ergaster. Интересно, что некоторые особи – видимо, самцы – были едва ли не в полтора раза крупнее других – очевидно, самок. Это предполагает резкий половой диморфизм по общим размерам тела – чуть ли не как у горилл, что явно противоречит ослабленному по размерам клыков. Как это понимать – непонятно. Не исключено, что мы несколько преувеличиваем уровень полового диморфизма, слишком много внимания уделяя крайним вариантам и недооценивая средние (Reno et al., 2003). Дискуссия на сей счет продолжается.
Образ жизни афарских австралопитеков, видимо, был непохож на известный у современных приматов. Жили австралопитеки маленькими группами, судя по “Первому семейству” – от десятка до двух особей. Днем австралопитеки кочевали по просторам Африки – саваннам и лесам, берегам рек и озер в поисках пропитания, а вечером забирались на деревья, как это делают современные шимпанзе. Питались они в основном растительной пищей.
По всей видимости, австралопитеки были способны перемещаться на довольно большие расстояния. Эта способность явилась одной из важнейших для дальнейшего расселения и эволюции человека, поскольку при миграциях гоминиды попадали в самые разнообразные экологические ниши. Это стало залогом пластичности современного человека и будущего завоевания мира.
Афарский австралопитек замечателен тем, что это ключевой вид для всего последующего разнообразия: поздние грацильные австралопитеки и локальные географические варианты, парантропы и варианты “ранних Homo” – все они произошли от A. afarensis. Да и сам этот вид нельзя расценивать как единый и на миллион лет застывший во времени: особи из Ворансо-Милле и Мака были очень крупными, в Хадаре от ранних к поздним, видимо, увеличивался размер мозга, хотя и очень медленно, менялись и зубы.
Родство афарских австралопитеков с южноафриканскими грацильными расценивается по-разному. Некоторые антропологи аргументируют значительную самостоятельность восточноафриканских афаренсисов и необходимость выделения их в самостоятельный род Praeanthropus и вид Praeanthropus africanus (Strait et al., 1997). Название взято из старого описания челюсти Гаруси, которая в последующее время считалась принадлежащей афарским австралопитекам. Однако на современном уровне знаний вероятнее ее принадлежность анамским австралопитекам, которые тогда должны бы называться Praeanthropus africanus, тогда как афарские – Praeanthropus afarensis. Впрочем, большая часть антропологов не разделяет этих воззрений и не считает необходимым и обоснованным возведение афаренсисов в ранг выше вида.
Сравнение морфологических черт позволяет ученым судить о том, какие грацильные австралопитеки имеют к нам отношение, а какие – нет. Из описанных видов наиболее подходит на роль предка именно афарский австралопитек. Хотя некоторые его черты не вписываются в образ прогрессивного предка – обезьянья форма позвонков и лопаток, костные гребни для мощных жевательных мышц и некоторые другие специализации, – антропологи не видят в этом угрозы для эволюционной теории; никто ведь не утверждает, что нашими предками были все без исключения представители афарских австралопитеков. Новые виды всегда возникают только из некоторых, уклоняющихся популяций старых видов. Афарские австралопитеки были очень изменчивы. Восточная Африка – немалый регион. В какой его области жили те самые австралопитеки, что были нашими непосредственными предками? Можно ли вообще найти ту самую группу? А может, они обитали в Северной или Центральной Африке? Если в Северной Африке найдены хотя бы несколько фрагментов костей австралопитеков, то огромная область в центре континента, несомненно кем-то густо заселенная в эпоху Великих Предков, в настоящее время – terra incognita для палеоантропологии. Тропический лес почти никогда не оставляет от животных ископаемых следов, слишком активна там жизнь. Кроме того, вечная политическая нестабильность государств Центральной Африки не способствует научным изысканиям. Но кто знает? Пятьдесят лет назад никто не ведал и о восточноафриканских австралопитеках.
Никто не ведал и о североафриканских австралопитеках. Пока в 1995 году французские палеонтологи не нашли в Республике Чад, в местности Коро-Торо, в долине реки Бахр-эль-Газал, нижнюю челюсть KT 12/H1 (Абель), к которой потом добавились еще одна нижняя KT 40 и верхняя KT 13–96-H1, а также изолированный зуб KT 12 P3/H2. Находка была описана как новый вид…
Австралопитек бахрэльгазальский Australopithecus bahrelghazali (Brunet et al., 1995, 1996). Первоначально он был датирован по фауне 3,0–3,5 млн лет назад, но после цифру уточнили и удревнили до 3,58 млн лет назад Строго говоря, отличия новоявленного североафриканского вида от афарского австралопитека не так уж велики – передняя сторона симфиза вертикальна у KT 12/H1 и округла у KT 40, но у обоих не скошена назад; на задней стороне симфиза рельеф очень слабый. Нижний клык выступает выше соседних зубов, но не запредельно – в рамках приличного афаренсисам.
Экологические условия в Коро-Торо в те времена отличались от восточноафриканских широким распространением саванн. Вот уж кто был настоящим покорителем просторов! Это отразилось и на диете бахрэльгазальцев: они ели гораздо больше саванных растений, чем все грацильные австралопитеки; по этому показателю их превзошли только восточноафриканские парантропы (Lee-Thorp et al., 2012).
В итоге бахрэльгазальский австралопитек выглядит как крайний вариант афарского; оно и логично – ведь хронологически эти виды совпадают, а географически разнесены примерно как анубис и гамадрил среди павианов. Расстояние имеет значение даже в африканской саванне. Фауна Коро-Торо вообще достаточно специфична – отсюда известно много эндемичных видов млекопитающих, близких к восточноафриканским, но отличающихся в мелочах. Видимо, и гоминиды менялись вдоль Сахеля, а это значит, что в Западной Африке нам стоит искать своего “гвинейского павиана” среди австралопитеков.
Но пока антропологи не добрались до гвинейских австралопитеков, им есть много работы и на востоке Африки. В 2015 году ряд эфиопских окаменелостей был описан как…
Австралопитек дейремеда Australopithecus deyiremeda (Haile-Selassie et al., 2015). Местность Ворансо-Милле, расположенная немного севернее Хадара, прославилась великолепными находками австралопитеков. Тут были найдены зубы, промежуточные между анамскими и афарскими австралопитеками, тут был откопан Большой Человек Кадануумуу, тут была открыта таинственная нога неведомой гоминидины. Сенсации так и прут из эфиопской земли. 2015 год не мог стать исключением. Строго говоря, удивительные находки были сделаны между 2006 и 2013 годами, но опубликованы в журнале Nature в 2015 году (Haile-Selassie et al., 2015).
В местонахождениях Бартеле (том самом, откуда происходит странная стопа) и Вайталейта в слоях с датировками 3,5–3,3 млн лет назад были обнаружены фрагмент лобной кости, два обломка верхних и два – нижних челюстей, а также восемь зубов. Не сказать, чтобы останки были чересчур представительны, но их признаков хватило, чтобы описать новый вид Australopithecus deyiremeda. Коллектив исследователей из США, Испании, Эфиопии и Германии представил доказательства особого статуса австралопитеков из Бартеле и Вайталейта. Особые признаки дейремеды – маленькие премоляры и моляры в сочетании с немаленькими резцами и клыками, крайне массивной нижней челюстью, а также не слишком выступающей вперед верхней челюстью. То есть, кратко говоря, – имеем странную смесь признаков грацильных и массивных австралопитеков.
Особую интригу привносит тот факт, что Бартеле расположен всего в тридцати пяти километрах севернее Хадара, откуда известны десятки классических находок афарских австралопитеков – с такими же в точности датировками. Так что – получается, в Эфиопии сосуществовали как минимум два вида грацильных австралопитеков? А если вспомнить еще и кениантропа – так и три? Чрезвычайно загадочно – как они могли жить одновременно? Что-то же должно было их разделять – скорее всего, некие особенности экологии. Но высшие приматы знамениты своей экологической пластичностью. Странно все это!
Не исключено также, что часть ранее известных костей, причислявшихся доныне к афарским австралопитекам, на самом деле принадлежит дейремеде. Может, именно этим объясняется слишком сильный половой диморфизм, высчитанный для афаренсисов некоторыми антропологами?
Но!
Действительно ли отличия челюстей из Бартеле достаточны для описания нового вида? Может быть, просто в очередной раз вернулась мода на дробительство? Пара челюстей, без сомнения, выделяется на общем афарском фоне, но лишь по паре признаков выходит за пределы известной изменчивости вида A. afarensis. А может, мы просто плохо знали популяционную вариабельность афаренсисов? Отличия дейремеды и афаренсисов не факт что превосходят различия, скажем, австралийских аборигенов и бушменов – что же, и современное человечество разделять на виды? Может, стоит вспомнить заветы Оккама?
Как обычно, только новые находки – а они обязательно последуют! – решат спор. А пока придется привыкать к странному слову – дейремеда.
Статус дейремеды находится под большим вопросом, но еще спорнее описанный в 2001 году не только вид, но и особый род…
Кениантроп плосколицый Kenyanthropus platyops (Leakey et al., 2001). От него известно не так много останков, зато каких! В Ломекви на севере Кении найден практически целый череп KNM-WT 40000, оттуда же происходят височная кость, верхние и нижние челюсти, а также целая россыпь зубов. Они имеют возраст 3,2–3,5 млн лет.
KNM-WT 40000 в целом похож на афарских австралопитеков, но отличается несколькими особенностями. Самая яркая его черта – плоское широкое лицо и вытянутый назад и вверх свод. Однако череп найден в крайне деформированном состоянии, так что оценивать его общую форму без тщательной реконструкции – дело безнадежное. Более существенными особенностями представляются очень высокие скуловые кости, уплощенный альвеолярный отросток верхней челюсти, короткое и широкое небо, чрезвычайно маленькое слуховое отверстие. Объем мозга из-за перекошенности черепа посчитать сложно, но он был примерно таким же, как у прочих грацильных австралопитеков. Нижние челюсти кениантропов массивные, с горизонтальной поверхностью за резцами. Зубная система кениантропа несколько специфична: резцы низкие, премоляры похожи на моляры, первый и второй моляры сравнительно с прочими грацильными австралопитеками заметно уменьшены, а вот третьи, напротив, очень крупные.
Как расценивать такой комплекс – большой вопрос. Авторы открытия считают его достойным возведения в самостоятельный род. Более того, высокое уплощенное лицо и пропорции зубов позволили предположить, что кениантроп – предок “человека рудольфского”, то есть с большой вероятностью и наш тоже. Или напротив, это тупиковая линия, зашедшая в могилу, а не устремившаяся к прогрессу?
Реконструкция черепа и более полные описания находок из Ломекви позволят уточнить статус кениантропа. Пока же его самостоятельность под большим сомнением, ведь по большинству черт он слабо отличается от давно и хорошо известного вида Australopithecus afarensis, а по крайней мере часть отличий запросто может быть следствием деформации (White, 2003). Даже если уплощенность лица, высоту скул, форму неба и пропорции зубов не списывать на тафономические искажения, все равно они вряд ли тянут на родовой статус, иначе расы современного человека тоже придется разносить по разным родам. Никто не отменял и индивидуальную изменчивость. А посему большинство антропологов используют термин “кениантроп” скорее как удобное личное имя черепа KNM-WT 40000, тем более что он, на удивление, до сих пор не обрел никакого эффектного прозвища.
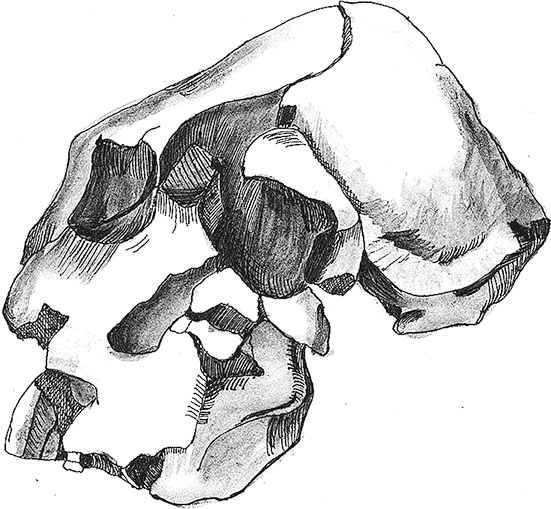
Рис. 8. Череп Kenyanthropus platyops.
В 2015 году в слоях Ломекви с датировкой 3,3 млн лет назад были найдены обломки, определенные как орудия особой ломеквийской культуры (Harmand et al., 2015). Кроме крайней примитивности, их отличают крупные размеры. А ведь именно величина – одна из основных характеристик орудий, используемых шимпанзе, капуцинами и макаками-крабоедами, в противоположность сравнительно небольшим олдувайским чопперам.
Значит ли это, что кениантропы обскакали всех прочих австралопитеков и действительно были предками Homo? Или же это был удивительный всплеск разума, обогнавший свое время и быстро затухший? Может, кениантропы были просто особо умными обезьянами? Ведь между орудиями из Ломекви и из Гоны сотни тысяч лет пустоты – преемственности между ними нет. Пока эти вопросы далеки от решения…
До сих пор речь шла о Восточной и Северной Африке, но примерно половина находок сделана на другой оконечности континента – в Южной Африке. Более того, тут австралопитеки были впервые открыты и обрели свое имя…
Австралопитек африканский Australopithecus africanus. Этот вид известен из ряда местонахождений – Таунг, Стеркфонтейн, Макапансгат, Глэдисваль. Название “африканский” не подразумевает, что были еще и какие-то неафриканские австралопитеки. Просто для Р. Дарта, давшего имя “южным обезьянам” в 1925 году, они были первыми ископаемыми человекообразными Африки. Сейчас их во избежание путаницы обычно называют либо южноафриканскими грацильными австралопитеками, либо африканусами.
Южноафриканские грацильные австралопитеки жили с 3,1 до 2,6 млн лет назад или, возможно, в большем интервале – вероятно, с 3,3 или даже 4,17 до 1,5 млн лет назад. Проблема в том, что в Южной Африке нет вулканов, а кости австралопитеков найдены в брекчиях известняковых пещер, практически напрочь лишенных стратиграфии. Посему датировать приходится по фауне, а она, во-первых, быстро не меняется, а во-вторых, не совсем идентична датированной восточноафриканской. В последнее время сделаны попытки датировать брекчию палеомагнитным методом и по соотношению космогенного алюминия-26 и бериллия-10. К сожалению, часто эти методы дают противоречивые результаты.
От южноафриканских грацильных австралопитеков известны все части скелета от множества особей, поэтому реконструкции их внешнего облика и образа жизни весьма достоверны.
Череп африканусов небольшой, с объемом мозга 428–515 см? или даже 370–540 см? (что зависит от реконструкции фрагментарных черепов), с умеренным надбровьем, у крупных самцов иногда, возможно, с сагиттальным гребнем (что, впрочем, только реконструируется из сильного сближения височных линий). Лицевой отдел крупный, выступающий вперед, хотя далеко не в такой степени, как у афаренсисов. Глазницы маленькие, нос в среднем шире, чем у Australopithecus afarensis, передняя сторона альвеолярного отростка верхней челюсти сильно уплощена, небо в передней части глубокое. Характернейшая черта африканусов – передние лицевые валики, утолщения верхней челюсти, идущие вертикально по бокам от носового отверстия; это не то же самое, что клыковые возвышения, типичные для существ с увеличенными корнями клыков. Замечательно, что передние лицевые валики типичны и для южноафриканских парантропов, что рассматривается как явное доказательство преемственности этих видов.
По некоторым признакам черепа африканусов выглядят прогрессивнее, чем у афаренсисов, что логично, учитывая их хронологию. Например, у A. africanus выше и округлее свод, слабее рельеф, сагиттальные гребни если и были, то низкие, а настоящих затылочных неизвестно вовсе. Однако большинство антропологов не считают африканусов нашими прямыми предками, так как в строении их конечностей хватает примитивностей и специализаций. К тому же древнейшие Homo Восточной Африки явным образом отличаются от африканусов. Правда, недавнее описание Australopithecus sediba из Южной Африки позволяет взглянуть на картину иначе, но об этом позже.
Посткраниальный скелет, как уже говорилось, имеет некоторые особенные черты. Например, африканусы имели шесть поясничных позвонков. Как и афаренсисы, южноафриканские грацильные австралопитеки обладали весьма архаичными костями рук, причем от лопаток до кистей степень примитивности падает. Лучевая кость Stw 46, с одной стороны, имеет сильно изогнутый нижний конец, с другой же – не имеет характерного для костяшкоходящих понгид костного гребня – ограничителя разгибания кисти. Кисть A. africanus находится на том замечательном уровне, когда и явного трудового комплекса вроде бы нет, но и задатки его вроде бы есть. Вероятно, они могли использовать орудия труда, но не изготовлять их.
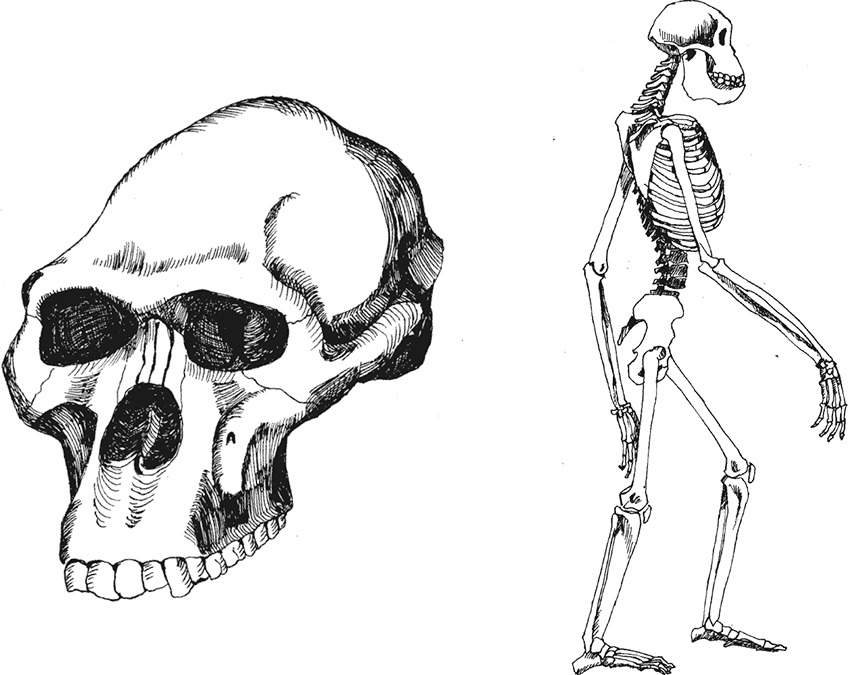
Рис. 9. Череп и скелет Australopithecus africanus.
В отличие от рук, таз и ноги африканусов очень продвинуты. Прямохождение этих существ несомненно. Имеются, конечно, и примитивные или специализированные черты – куда без них, – но в целом ноги отличались от наших несущественно. Одной из особенностей африканусов является большая ширина, на которую были разнесены вертлужные впадины таза и, соответственно, тазобедренные суставы. Думается, походка из-за этого должна была быть несколько замедленной, возможно – вразвалочку, но это, в сущности, мелочи. Интерес представляет большая берцовая кость Stw 514, в строении которой больше шимпанзоидных особенностей, чем на костях афарских австралопитеков. Впрочем, другие берцовые кости из Стеркфонтейна далеко не столь понгидны.
Рост африканусов был все тот же – от метра до полутора, вес – от 20 до 40 кг. Руки африканусов относительно тела и ног сильно удлинены, даже в большей степени, чем у более древних афаренсисов. Обращает на себя внимание, что кости рук A. africanus крупнее, чем у A. afarensis, а кости ног обоих видов либо равны, либо у A. africanus грацильнее.
Кстати, о поколениях…
В 1978 г. в уютном отнорке Стеркфонтейна под названием грот Сильберберг, представляющем собой кривую, сырую и холодную шахту четырнадцатиметровой глубины, была найдена лодыжка и четыре сочлененных кости левой стопы. Они были определены как останки мартышки и мирно пролежали в ящиках с костями млекопитающих до 1994 г., когда Р. Кларк и П. Тобайас извлекли их из забвения и обнаружили, что кости на самом деле австралопитечьи (Clarke et Tobias, 1995). Останки получили номер Stw 573 и сразу два имени – Маленькая Стопа и Синдерелла. Прошло еще несколько лет, и в 1998 г. в пещере нашлось эффектное продолжение – череп с нижней челюстью и, вероятно, целый скелет. Была только одна проблема – кости Синдереллы оказались намертво вмурованы в скалу, так что снаружи они выглядели как нарисованные. С тех пор и по сию пору палеонтологи продолжают упорно выцарапывать бесценные мощи из каменного плена. Дело это невероятно изматывающее, ведь работать приходится в узкой каменной щели, хитро изогнувшись наискосок, маленькими иголками, чтобы не повредить уникальную находку. К 2014 г. череп почистили настолько, что он оставался прикрепленным к стене только правой щекой, а в начале 2015-го наконец-то отковыряли! Но в стене поныне остаются многие другие кости.
Особый интерес Синдерелла представляет по двум причинам. Во-первых, это первый целый скелет южноафриканских австралопитеков и потенциально самый целый среди вообще всех австралопитеков. Во-вторых, отложения грота, согласно анализу космогенного алюминия и бериллия, могут иметь возраст 3,3, 3,67 или даже 4,17 млн лет назад – существенно больший, чем в других местонахождениях Южной Африки; впрочем, это не доказано окончательно: по фауне, магнитостратиграфии и результатам урано-свинцового анализа получается лишь 2,2 млн лет назад – тогда Синдерелла, напротив, оказывается одним из самых молодых австралопитеков.
Маленькая Стопа – сенсация, длящаяся уже целое поколение, может растянуться еще на много лет.
В другом месте Стеркфонтейна – Яковец-Каверн – с аналогичной и опять неясной датировкой (возможно, 4 млн лет, но, быть может, вдвое меньшей) найдены части другого скелета с черепом (Partridge et al., 2003). Эти фрагменты похожи на прочих австралопитеков, но имеют некоторые необычные черты: лоб чрезвычайно узкий и крайне покатый, ключица резко уплощена, шейка бедренной кости очень длинная и сильно опущенная. По некоторым признакам австралопитек из Яковец-Каверн вполне годится на роль предка парантропов.
Не так много мы знаем о жизни южноафриканских австралопитеков, как хотелось бы. Особенности строения стремечка Stw 151 позволили заключить, что слух грацильных австралопитеков был более чувствительным в области высокочастотных звуков по сравнению с современным человеком, то есть как бы более обезьяньим (Moggi-Cecchi et Collard, 2002). Диета разных особей, судя по микроизношенности эмали и изотопному анализу, была весьма разнородной: у большинства индивидов больше половины рациона составляли древесные плоды и листья, некоторые же столовались преимущественно в саванне (Ungar et Sponheimer, 2011). Этим африканусы, кстати, сильно отличались от анаменсисов и афаренсисов, кои питались существенно однообразнее. Вероятно, это связано с умеренностью климата Южной Африки и более выраженной там сезонностью. Более того, у африканусов выражена “внутризубная” изменчивость соотношений Sr/Ca и Ba/Ca: судя по всему, они периодически переходили с трав на мясо и обратно (Balter et al., 2012). Пластичностью они, кстати, превосходили даже “ранних Homo” Южной Африки, которым вроде как по статусу наших предков положено быть во всем первыми.
Систематика австралопитеков не так однозначна, как может показаться при чтении учебников. Южноафриканские антропологи, прежде всего Р. Кларк, давно аргументируют существование как минимум двух видов в Стеркфонтейне (например: Clarke, 2008). Один из них – “классический” Australopithecus africanus, тогда как второй – гораздо более крупный и массивный парантропоподобный, который, собственно, был предком массивных южноафриканских австралопитеков.
Уголок занудства
Предполагаемый парантропоподобный вид грацильных австралопитеков Южной Африки отличается целым комплексом черт: надбровье слабо выступает, вдавленность над ним минимальна, надглазничный край сравнительно тонкий; у самцов есть маленький сагиттальный гребень на задней части свода; затылок высокий и слабо выступающий назад; лицо очень высокое и длинное, вдавлено в поперечном направлении, скуловые кости резко выступают и развернуты вперед, при взгляде сбоку закрывают края носового отверстия; из-за этого лобный отросток верхней челюсти расширен вбок и ориентирован фронтально; межглазничное пространство широкое; точка “назион” (место соединения носовых костей с лобной) расположена выше лобно-верхнечелюстного шва, близко к “глабелле” (самой выступающей точке надбровья); нижняя челюсть большая и массивная; резцы и клыки увеличены относительно заклыковых зубов; премоляры и моляры большие, со вздутыми боками и бугорками, приближенными к центру зуба.
К этому виду относят черепа и челюсти Stw 252, Sts 71, Stw 505, Stw 183, Stw 498, Stw 384, Sts 1, MLD 2 и некоторые зубы.
При этом индивиды Sts 19, Sts 51 и Sts 52a, Stw 183, Stw 252, Stw 255 каждый в отдельности обладают специфическими чертами, иногда уникальными, иногда сближающими своих носителей с парантропами или “ранними Homo”. Одни исследователи относят это на счет индивидуальной изменчивости, другие – эволюционной переходности. Возможно, совершенствование методов датирования позволит прояснить ситуацию.
Впрочем, другие исследователи считают, что нет нужды разделять африканусов на два вида, а перечисленные выше различия объясняются половым диморфизмом, возрастной и индивидуальной изменчивостью или, возможно, хронологическими изменениями.
Заметим в скобках, что Р. Брум в 1936 году на основании разрушенного черепа Sts 60 из Стеркфонтейна описал вид Plesianthropus transvaalensis, а Р. Дарт в 1948 году по задней части мозговой коробки MLD 1 из Макапансгата – Australopithecus prometheus. Эти названия использовались некоторое время для обозначения грацильных австралопитеков из соответствующих местонахождений, но ныне обычно признаются полными синонимами Australopithecus africanus. Впрочем, некоторые исследователи считают название Australopithecus prometheus подходящим для обозначения того самого парантропоподобного вида, особенно если признать его б?льшую древность.
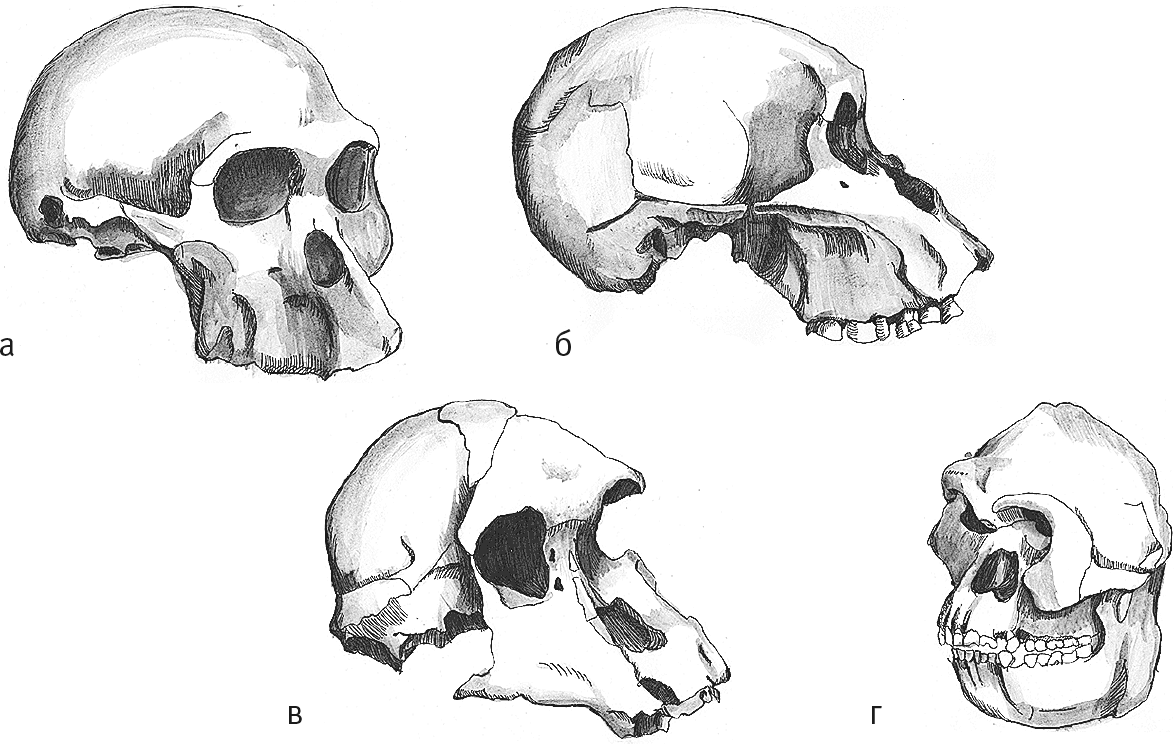
Рис. 10. Черепа Sts 5 (а), Sts 71 (б), Stw 505 (в), Stw 573 (г).
Другой важный вопрос – наличие Australopithecus africanus в Восточной Африке. К этому виду были отнесены грацильные австралопитеки из средних слоев Омо с датировками около 2,3 млн лет назад (например: Hunt et Vitzthum, 1986). Некоторые антропологи считают, что к африканусам можно причислить останки из местонахождения Южный Таквил (3,5 млн лет назад) и находки из Боури, описанные как A. garhi, в частности BOU-VP-12/130 (2,5 млн лет назад). С одной стороны, окаменелости из этих местонахождений очень фрагментарны, с другой – имеют отличия от “классических” афаренсисов из Лаэтоли и Хадара, с третьей – похожи на южноафриканских грацильных, но все же не совсем идентичны им. Учитывая, что грацильные австралопитеки Южной Африки в среднем моложе восточноафриканских, можно расценивать их различия как обусловленные временем, тогда находки из Омо и Боури логично вписываются в концепцию “хронологического A. africanus”. Если же считать, что грацильные австралопитеки Южной и Восточной Африки возникли независимо друг от друга, то надо допустить миграции африканусов с юга до Эфиопии около 2,5 млн лет назад, если не раньше.
Большинство современных антропологов считают, что A. africanus не участвовал в сложении нашего вида, но так думали не все и не всегда. В первой половине XX века, до открытий в Восточной Африке, африканусы были единственными известными обезьянолюдьми, и неудивительно, что многие ученые старательно выискивали в них человеческие черты. Так, черные прослои и пятна на костях из Макапансгата были приняты за следы древнейших очагов и обугленности. После выяснилось, что это окислы марганца. Р. Дарт собрал впечатляющую коллекцию костей, рогов и зубов из южноафриканских пещер и описал остеодонтокератическую – костно-зубо-роговую – культуру (Dart, 1957). Сломы на концах костей и рогов он принял за результаты использования, а разрушения на черепах павианов – за следы ударов костями (причем якобы повреждено аж 80 % найденных черепов, а расположение проломов позволило утверждать о праворукости охотников), а также считал, что двойные вмятины на некоторых из этих черепов соответствуют мыщелкам длинных костей копытных, найденных рядом.
Надо отдать Р. Дарту должное, он не был голословен: среди богатых этнографических коллекций, собранных среди бушменов, он обнаружил удивительно точные параллели остеодонтокератическим обломкам. Некоторые изделия, кажется, делал один человек, только вот на бушменских есть орнамент, а на австралопитечьих – нет. Однако исследование некоторых образцов из “остеодонтокератической” коллекции новейшими методами показало, что характерные повреждения их концов – это погрызы гиен, а не рабочая изношенность. Так что, видимо, “культура” австралопитеков на самом деле является объедками падальщиков; впрочем, проверялись далеко не все собранные Р. Дартом кости, так что надежда на реабилитацию южноафриканских австралопитеков еще остается.
Светлым лучиком в беспробудной тьме, окружающей интеллект австралопитеков, является одна уникальная находка: в Макапансгате в брекчии с датировкой 2,5–3 млн лет найден круглый камень красного цвета – размером с кулак без следов обработки. Самое интересное, что ближайшее местонахождение такой породы расположено за 32 км от пещеры. Ясно, что камень не сам прополз эту дистанцию. А ведь это целый дневной переход – немалое расстояние, тем более что нести камень надо было все время в руке, корзин и мешков-то еще никто не изобрел! Более того, естественные выщерблины на камне напоминают глаза и рот, образуя чудную рожицу. Как знать, может, некий первобытный эстет, гуляя по родной степи, усмотрел в этом булыжнике свое отражение и не поленился притащить его домой? Камень из Макапансгата – древнейшее “прикольно” или “хочется чего-то этакого, чтоб душа развернулась, а потом обратно завернулась”. А может, это самый первый смайлик?
Мозговитый Беби из Таунга – несостоявшийся прогресс?
Чем больше антропологи смотрят на старые находки, тем больше интересного видят. Ныне взгляд обратился на дартовского Беби из Таунга – классический череп детеныша австралопитека, первый в великолепном ряду последующих открытий. Кажется, за предыдущие без малого 90 лет про Таунга выяснили все что можно, например, что его заклевал злой орел. И вот теперь международная (какая же еще в век глобализации?) группа исследователей заострила свое внимание на наличии следов метопического шва на эндокране – естественном слепке мозга – Таунга. Метопический шов – это шов между двумя половинками лобной кости. То есть лобная кость у ребенка вообще-то изначально парная, но правая и левая части обычно срастаются примерно к двум годам. Однако же следы шва, а то и он весь регулярно остаются незаращенными. Этот факт всегда обращал на себя внимание антропологов, но в палеоантропологическом аспекте им занимались мало, обычно просто констатируя случаи сохранения шва на взрослых черепах.
В нынешнем же исследовании ученые сравнили частоту незаращения шва у шимпанзе, ископаемых гоминид и современных людей (Falk et al., 2012). Выяснилось, что у обезьян метопический шов закрывается почти у всех детенышей к моменту прорезывания второго молочного моляра, а ко времени прорезывания третьего постоянного у всех поголовно особей исчезают даже его следы. У современных же людей, хотя между первым и вторым молочными молярами частота наличия шва резко падает, частенько шов или его следы остаются на всю жизнь.
У ископаемых гоминид картина тоже весьма показательна. Среди массивных австралопитеков – южно- и восточноафриканских – не известно ни одного случая сохранения хотя бы следов шва. А вот среди грацильных австралопитеков, “ранних Homo”, эректусов и неандертальцев частота шва или его следов весьма высока, видимо, не меньше, чем среди современных людей.
Авторы статьи предположили три причины такой ситуации. Во-первых, шов мог сохраняться для облегчения родов большеголовых детенышей прямоходящими самками. Правда, остается неясным, зачем тогда шву быть открытым многие годы после этого радостного события? И почему тогда парантропы остались в стороне от прогресса? Мозгов у них было примерно столько же, что и у грацильных австралопитеков, а челюсти – крупнее. Во-вторых, долгое сохранение шва после родов могло способствовать росту мозга у детеныша. Наконец, в-третьих, оно может отражать прогрессивную перестройку новой коры головного мозга, в особенности лобной доли – самой продвинутой, отвечающей за мышление как таковое. Очевидно, все эти причины взаимосвязаны и, скорее всего, действовали одновременно. В этом свете логично, что шимпанзе и парантропы рано обретают “твердолобость” – жевательные мышцы для них важнее интеллекта.
Красиво, хотя моя ехидная память подсказывает одно маленькое но. Помнится, в 1995 г. одна из авторов нынешней статьи – Дин Фальк, известная своими исследованиями ископаемых эндокранов, в составе авторитетного авторского коллектива доказывала, что Беби из Таунга с наибольшей вероятностью – детеныш не африканского австралопитека, а парантропа (Falk et al., 1995). Уникальная закавыка систематики, если учесть, что Беби – голотип Australopithecus africanus! Впрочем, может, среди парантропов были свои прогрессивные представители. С другой стороны, он же погиб ребенком, явно не оставив своих продвинутых генов в потомстве; крючковатые когти орла пресекли прогресс парантропов… Родители-то были еще дремучие, не отмахались от грозной птицы, не уберегли драгоценное чадо! Приятно сознавать, что наши предки, обогащенные позднесрастающимся метопическим швом, оказались смышленее и прогрессивнее. Не то эту статью писал бы этакий клацающий челюстями “щелкунчик”-постпарантроп. Однако повторим великую истину: палеонтология не терпит сослагательного наклонения.
Кстати, об ужасах систематики…
Как было упомянуто выше, Беби из Таунга запросто может быть детенышем массивного австралопитека (Falk et al., 1995). В пользу этого говорят три основных признака, типичных именно для парантропов: во-первых, увеличенный затылочно-краевой синус – сосуд на задней стороне мозга для оттока венозной крови; во-вторых, наднебное расширение верхнечелюстных синусов – полостей в теле верхней челюсти; в-третьих, сравнительная площадь бугорков на первом нижнем моляре. По всем этим показателям Таунг оказывается парантропом. Но можно ли тогда вообще называть его этим словом? Ведь Таунг – голотип вида Australopithecus africanus, значит, это название по правилу приоритета должно использоваться для обозначения южноафриканских массивных австралопитеков. Тогда южноафриканские грацильные будут носить имя Plesianthropus transvaalensis, восточноафриканские грацильные – Plesianthropus afarensis, нынешние анаменсисы должны бы именоваться Plesianthropus africanus (учитывая название Meganthropus africanus, данное Г. Вейнертом в 1950 г. челюсти из Гаруси), а восточноафриканские массивные – Australopithecus boisei. И это без учета мнений, согласно которым восточноафриканские грацильные достойны собственного родового названия Praeanthropus, а восточноафриканские массивные – Zinjanthropus.
Вот к чему приводят случайности! Если бы Р. Дарту первым попался череп взрослого австралопитека, этой ужасной путаницы удалось бы избежать. А теперь систематики вынуждены иногда просыпаться в холодном поту и с липкими руками – когда им снится кошмар, что кто-то особо въедливый и принципиальный поставил-таки вопрос ребром о пересмотре названий. Избежать этого ужаса можно, доказав, что все же Таунг – грацильный австралопитек, или переопределив голотип. Впрочем, антропологи осознают, какие названия и по отношению к кому они используют, друг друга они понимают, а потому живут без лишних формальностей.
На исходе века, в 1999 году, научный мир узнал о новом виде австралопитеков…
Австралопитек гари Australopithecus garhi – крайне любопытное существо, фрагменты черепа и конечностей которого найдены в Эфиопии в местонахождениях Боури, Гамеда, Матабайету, а также, возможно, Ндоланья и Кооби-Фора и имеют возраст 2,5 млн лет (Asfaw et al., 1999). Это – самый поздний из грацильных австралопитеков. Здорово, что в Боури обнаружен фрагментарный скелет BOU-VP-12/1, отлично описанный (DeGusta, 2004), только отчего-то непропиаренный и потому малопопулярный.
Череп BOU-VP-12/130 своеобразен: лобная кость с достаточно мощным надбровьем, а височные линии, видимо, еще на лобной кости сходились в сагиттальный гребень, который заканчивался уже на теменной, – такой вариант резко отличается от версии A. afarensis и A. africanus, у которых гребень был смещен далеко назад. Уникальна верхняя челюсть: альвеолярный отросток выпуклый как поперечно, так и продольно, крайне сильно выдается вперед, отчего его передний конец аж поднимается наверх, а резцы оказываются торчащими вперед. Такого прогнатного гоминида планета не видела ни до, ни после. Форма альвеолярной дуги – как верхней, так и нижней – крайне архаичная, U-образная, больше всего похожая на типичную для A. anamensis – гораздо более древнего вида. Другой особенностью A. garhi являются очень большие зубы – как задние, так и передние. Моляры этого вида сопоставимы со значениями не самых крупных парантропов, но вот огромные резцы ставят A. garhi явно особняком.
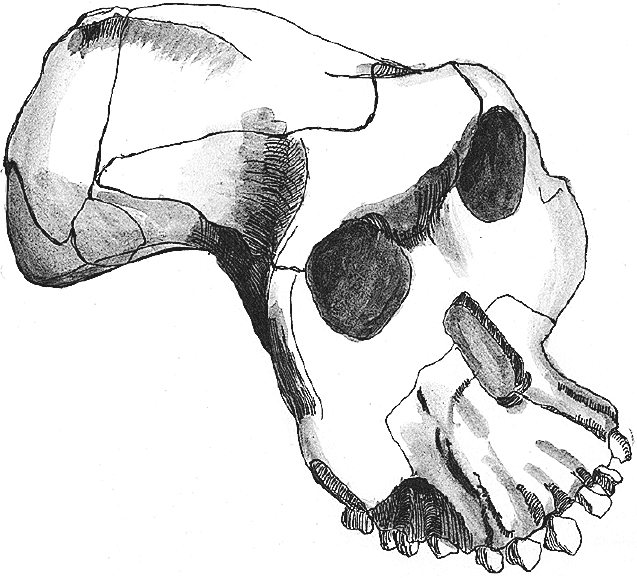
Рис. 11. Череп Australopithecus garhi.
Крайне интересно, что посткраниальные кости A. garhi больше похожи на таковые “ранних Homo”, чем A. afarensis. Это довольно неожиданно, коли уж гари не был нашим предком. Закономерно же то, что по строению скелета A. garhi родственнее восточноафриканским гоминидам, чем южноафриканским.
Рост A. garhi был стандартнейшим для австралопитеков – от метра до полутора. А вот пропорции конечностей BOU-VP-12/1 уникальны, такие не встречены больше ни у каких гоминид: руки были, видимо, резко удлинены относительно ног, причем в основном за счет увеличенных предплечий.
Получается, что гари был каким-то очень странным австралопитеком – последним грацильным, но очень уж крупнозубым, специализированным по черепу, но продвинутым по скелету. Он явно не входил в число наших предков, иначе пришлось бы допустить быстрое увеличение зубов и челюстей, а после такое же быстрое уменьшение, да и специализации его слишком своеобразны.
Как уже говорилось выше, австралопитеки гари с большой вероятностью изготавливали каменные чопперы и умели разделывать ими антилоп. А ведь к этому времени уже появились “ранние Homo” – наши прямые предки. Выходит, в Восточной Африке существовали как минимум две независимые, хотя и родственные группы, находившиеся на пути очеловечивания. Судя по тому, что Читатель этой книги явно не потомок австралопитеков гари, победили Homo, хотя мы и не знаем почему. Плюс к этому появившиеся тогда же массивные австралопитеки несколько позже тоже, видимо, стали изготавливать каменные орудия, причем восточноафриканские делали это независимо от южноафриканских.
Кстати, о массивных австралопитеках…
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.