4.1.2 Череп
Череп принято рассматривать, как единое образование, подразделяющееся на мозговой (осевой) и висцеральный отделы. Следует, однако, иметь в виду, что и по выполняемой функции, и по характеру закладки и онтогенетического развития эти два отдела представляют собой совершенно самостоятельные образования, и их объединение во внешне единую структуру – явление вторичное. У круглоротых и рыб осевой и висцеральный отделы черепа выражены отчетливо и слабо связаны друг с другом.
Мозговой череп. Черепная коробка (мозговой череп) формируется как продолжение осевого скелета из хрящевых закладок соединительнотканной оболочки переднего конца хорды (парахордалии) и самостоятельных очагов образования хряща сбоку (боковые хрящи) и впереди от них (трабекулы). Они составляют основу дна черепа. С боков и спереди к дну примыкают хрящевые капсулы органов чувств – обонятельные, глазные и слуховые.
Отделы мозгового черепа формируются путем смешанного сочетания элементов. Парахордалии и затылочные хрящи закладываются из ткани, общей с позвоночником, – мезенхимы склеротомов, трабекулы и капсулы органов чувств – из скелетогенной ткани кожного происхождения, которую продуцирует нервный гребень (Ф.Я.Дзержинский, 1998).
У позвоночных выделяют два типа строения мозгового черепа: платибазалъный и тропибазалъный. Платибазальный череп имеет широкое основание. Глазницы широко раздвинуты, мозговая полость продолжается между ними. Такой тип черепа характерен для хрящевых рыб, круглоротых, амфибий. Настоящие наземные позвоночные имеют тропибазальный тип черепа: глазницы в нем сближены и часто разделены лишь межглазничной перегородкой. Мозговая полость при этом оттеснена назад. О различии в организации двух типов мозгового черепа можно судить по расположению в них головного мозга (рисунок 10).
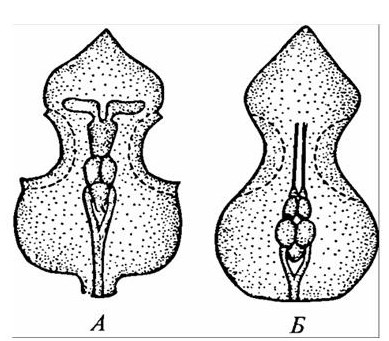
Рисунок 10 – Положение мозга в платибазальном (А) и тропибазальном (Б) черепах
У круглоротых и хрящевых рыб дно черепа закладывается в виде парных хрящей – парахордалии и трабекул (рисунок 11). У ним примыкают капсулы органов чувств: непарная обонятельная, парные глазничные и слуховые (рисунок 12). У хрящевых рыб хрящевой мозговой череп приобретает вид оформленной коробки, так как капсулы органов чувств, прирастая к дну черепа, образуют бока черепа и частично формируют крышу. Крыша никогда не бывает полной, в ней присутствует чаще одно (иногда больше) отверстие – фонтанелъ. Эта неполнота хондральной крыши проявляется в онтогенезе большинства других позвоночных. У взрослых акул образуется единый хрящевой череп, в котором можно выделить несколько морфологически и функционально различающихся основных отдела мозгового черепа: затылочный, слуховой, глазничный и обонятельный.
Затылочный отдел прикрывает головной мозг сзади, в нем имеется большое затылочное отверстие, через которое проходит спинной мозг. Под ним располагается затылочный мыщелок, который смыкается с первым позвонком.
Слуховой отдел соответствует вросшим в стенку черепа слуховым капсулам, окружающим внутреннее ухо. Спереди от затылочного отдела мозговая коробка расширяется, образуя слуховыекапсулы, окружающие мешочки и каналы внутреннего уха. Затем спереди мозговая коробка сужается, образуя глазничные впадины: в них расположены глазные яблоки и их мускулатура. Здесь, в дне мозговой коробки, имеется срединное углубление, к которому прилегает гипофиз.
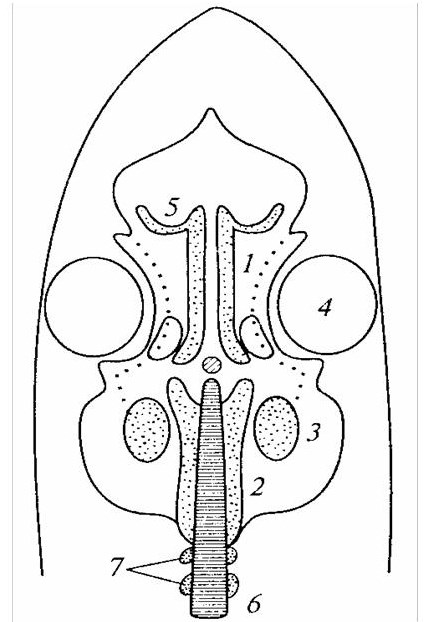
1– трабекулы; 2 – парахордалии; 3 – слуховая капсула; 4 – глазница; 5 – обонятельная капсула; 6 – хорда; 7 – закладка.
Рисунок 11 – Расположение хрящевых закладок мозгового черепа акулы
Обонятельный отдел находится в передней части черепа и представляет собой парные обонятельные капсулы, выстланные с внутренней поверхности обонятельным эпителием. Снизу черепную коробку подстилает дно черепа, а сверху она закрыта хрящевой крышей с небольшим отверстием – фонтанелью. Заканчивается мозговой череп спереди рылом – рострумом, состоящим из трех хрящей (рисунок 13).
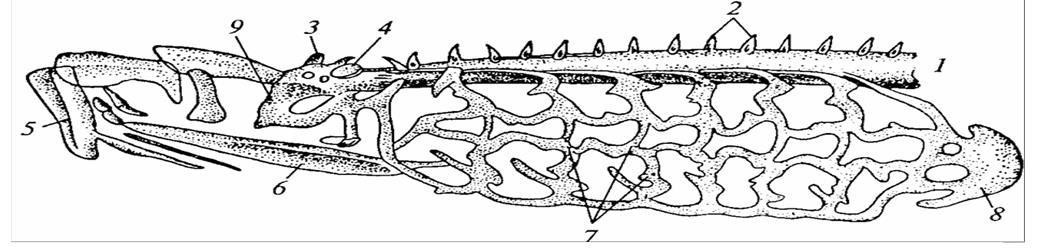
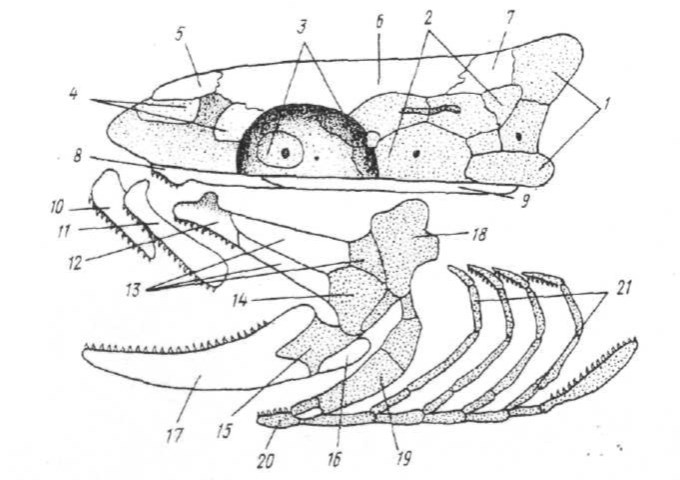
1 – хорда; 2 – зачатки верхних дуг позвонков; 3 – обонятельная капсула; 4 – слуховая капсула; 5 – скелет присасывательной воронки; 6 – подъязычный хрящ; 7 – жаберные дужки; 8 – околосердечный хрящ; 9 – подъязычная дуга.
Рисунок 12 – Череп и скелет жаберного аппарата миноги
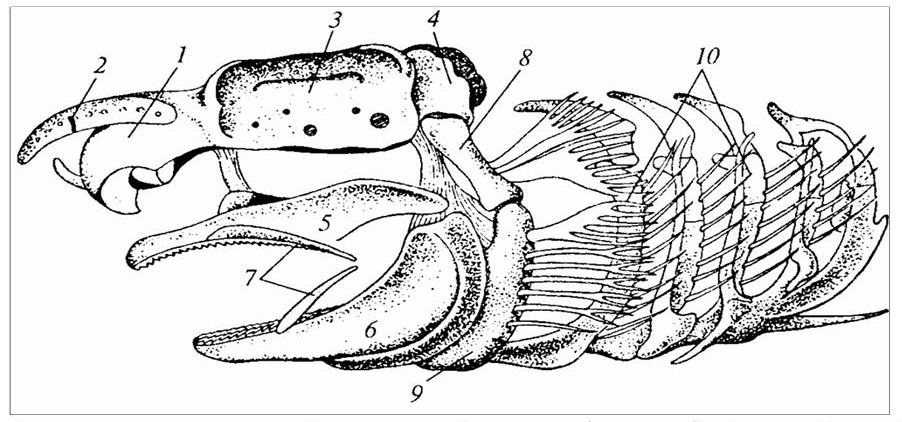
1 – носовая капсула; 2 – носовой вырост (рострум); 3 – орбита; 4 – затылочная капсула; 5 – нёбноквалратный хрящ (верхняя челюсть); 6 – меккелев хрящ (нижняя челюсть); 7 – губные хрящи; 8 – подвесок (гиомандибуляре); 9 – гиоид; 10 – жаберные дуги.
Рисунок 13 – Череп акулы
В мозговой коробке имеются многочисленные отверстия, через которые проходят черепные нервы и кровеносные сосуды. В срединное углубление крыши мозговой коробки открываются каналы из полостей внутреннего уха. Снизу в области коробки располагаются отверстия для сонных артерий, снабжающих кровью мозг, а сбоку другие отверстия для артериальных и венозных сосудов. Через хордальный канал в дне мозговой коробки у взрослых акул и зародышей других рыб вперед до гипофиза тянется хорда.
Мозговой череп костных рыб слагается из костей разного происхождения. Одни – хондральные – замещают хрящ в первичном хрящевом черепе, другие – покровные, формируются в соединительнотканном слое кожи. Исторически покровный костный скелет считают первичным. У костных рыб осевой череп частично или (у костистых) полностью окостеневает. Группы окостенений соответствуют отделам черепа хрящевых рыб: в затылочной области образуются затылочные кости, в слуховой – ушные, в стенках глазницы – клиновидные, в области обонятельных капсул – обонятельные кости. В крыше черепа располагаются парные носовые, лобные и теменные кости вторичного происхождения. Дно черепа представлено также вторичными костями: непарными сошником и парасфеноидом. Костный череп более легок и прочен (рисунок 14).
У современных амфибий заметна тенденция к относительному расширению и уплощению мозговой коробки и редукции ее окостенений. Мозговой череп амфибий значительной части остается хрящевым, что обусловлено слабым развитием как хондральных, так и накладных окостенений.
Осевой череп: 1 – затылочные кости, 2 – ушные, 3 – клиновидные, 4 обонятельные, 5 – носовые, 6 – лобные, 7 – теменные, 8 – сошник, 9 – парасфеноид; висцеральный череп: 10 – предчелюстные кости, 11 – верхнечелюстные, 12 – небные, 13 – крыловидные, 14 – квадратная, 15 – сочленовная, 16 – угловая, 17 – зубная, 18 – подвесок, 19 – гиоид, 20 – копула, 21– жаберные дуги.
Рисунок 14 – Череп костистой рыбы (схема)
У современных амфибий в сравнении с ранними тетраподами сильнее выражена редукция крыши черепа. В широком и плоском черепе современных бесхвостых и хвостатых амфибий сохранилась лишь малая доля покровной крыши предковых форм (рисунок 15).
В мозговой коробке амфибий развиваются следующие хондральные кости. В затылочной области есть две боковые затылочные кости. На них заметны два затылочных мыщелка. Места, соответствующие основной и верхней затылочным костям рыб, у амфибий остаются хрящевыми. В области слуховой капсулы формируется одна небольшая переднеушная кость, большая часть капсулы остается хрящевой. В передней части глазницы мозговой череп сужается. Здесь у бесхвостых амфибий развивается клинообонятельная кость; у хвостатых эта кость парная. Обонятельная капсула остается хрящевой.
Покровных костей также немного. Крышу черепа составляют теменные и лобные кости, которые у бесхвостых срастаются в лобно-теменные кости. Кпереди от них находятся носовые кости, у безногих они срастаются с предчелюстными костями. По бокам задней части черепа расположены чешуйчатые кости, особенно сильно развитые у безногих. Дно черепа подстилает большой крестообразный парасфеноид, а впереди от него лежат парные сошниковые кости. Снизу на черепе видны отверстия для выхода нервов (зрительного и тройничного). На поверхности крыши слабо заметными желобками проступают трубчатые ходы боковой линии. У древних тетрапод поверхность крыши была структурирована гребнями и ямками, к которым прикреплялась кожа.
1 – межчелюстная кость; 2 – сошник; 3 – носовая полость; 4 – носовая кость; 5 – клинообонятельная (поясок); 6 – лобная; 7 – теменная; 8 – переднеушная; 9 – чешуйчатая; 10 – боковая затылочная; 11 – стремячко; 12 – квадратная; 13 – сочленовная; 14 – гиоид; 15 – подъязычная пластинка; 16 – угловая; 17 – зубная; 18 – нёбная; 19 – верхнечелюстная; 20 – квадратно-скуловая; 21 – крыловидная; 22 – парасфеноид; 23 – хоаны.
Рисунок 15 – Схема строения черепа земноводного (А – сбоку; Б – снизу; хрящевые элементы показаны точками, хондральные кости – черным цветом, покровные – светлым)
Общей особенностью мозгового черепа рептилий является почти полное окостенение первичного хрящевого черепа и развитие большого числа кожных костей, формирующих крышу, бока и дно черепа.
Затылочная область мозгового черепа состоит их четырех костей хондрального происхождения: верхнезатылочной, основной затылочной и двух боковых затылочных (рисунок 16). Все эти кости окаймляют затылочное отверстие, книзу от которого лежит единственный затылочный мыщелок. В его образовании участвуют обе боковые затылочные и основная затылочная кости.
Спереди от основной затылочной кости лежит основная клиновидная кость, принимающая участие в образовании дна черепа. У некоторых рептилий – (змей, крокодилов – есть дополнительные боковые клиновидные кости. Парасфеноид мал и расположен у переднего конца основной клиновидной кости. Сошниковые кости редуцированы.
В области слуховых капсул развиваются три ушные кости. Одна из них – переднеушная – пожизненно остается самостоятельной, дне другие срастаются: заднеушная – с боковой затылочной костью, а верхнеушная – с верхнезатылочной костью.
В обонятельной области хондральные окостенения не развиваются и она остается хрящевой.
1 – теменные кости (1' – отверстие для теменного глаза); 2 – лобные; 3 – предлобные; 4 – надглазничные; 5 – заглазничные; 6 – носовые; 7 – верхнечелюстные; 8 – межчелюстные; 9 – сошники; 10 – нёбные; 11 – хоаны; 12 – крыловидные кости (12' – зубы на крыловидных костях); 13 – квадратные; 14 – поперечные; 15 – основная затылочная; 16 – затылочный мыщелок; 17 – основная клиновидная; 18 – остаток парасфеноила; 19 – скуловая; 20 – слезная; 21 – столбчатая (надкрыловидная); 22 – чешуйчатая; 23 – надписочная; 24 – зубная; 25 – сочленовная; 26 – угловая;27 – надугловая; 28 – венечная кость.
Рисунок 16 – Череп ящерицы (А – сбоку; Б – сверху; В – снизу)
Крыша черепа образована парными накладными костями: носовыми, предлобными, лобными, заднелобными, теменными и непарной межтеменной. Последняя имеет в середине отверстие для теменного органа. Это отверстие есть у ящериц и гаттерий, однако в черепе змей, черепах и крокодилов оно отсутствует.
Бока черепа слагают многочисленные накладные кости: непарная межчелюстная, парные верхнечелюстные, надглазничные, ску-ювые, чешуйчатые. Под крышей черепа, ниже височной кости, у ящериц расположена большая впадина – боковая височная яма.
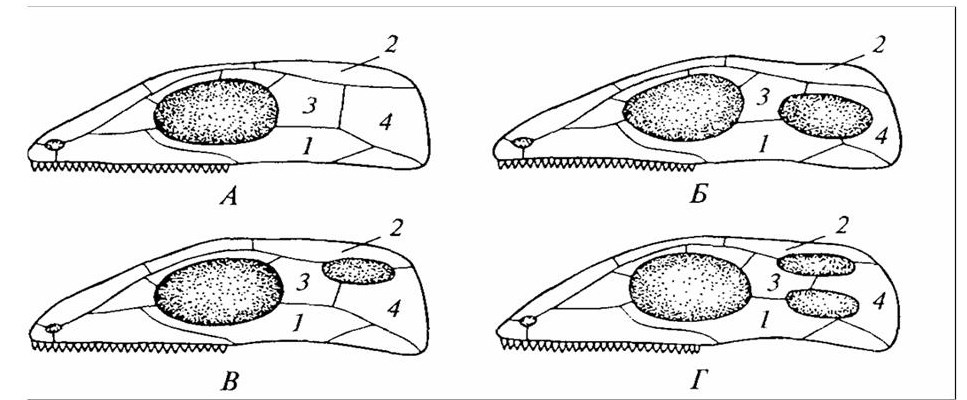
А – анапсидный тип (котилозавры, черепахи); Б – синапсидиый тип (звсроподобные рептилии); В – эвриапсидный тип (вымершие плезиозавры и др.); Г – диапсидный тип (клювоголовые; архозавры, а также ящерицы и змеи, утратившие одну или обе височные дуги); 1 – скуловая кость; 2 – теменная; 3 – заглазничная; 4 – чешуйчатая.
Рисунок 17 – Схемы расположения височных ям в черепах разных типов рептилий
Многие важные изменения в крыше черепа рептилий связаны с развитием височных впадин (окон, ям). Для ранних рептилий (котилозавров, черепах) характерна сплошная крыша без впадин. Этот примитивный тип называли анапсидным (рисунок 17). Черепа современных черепах в сравнении с древними изменились: у многих видов теменные кости часто образуют глубокие выемки. Иногда вся височная область превращается во впадину, и элементы крыши от глазницы до заднего края черепа отсутствуют.
У большинства рептилий в процессе эволюции в области виска и щеки развились одна или две впадины. Этот признак сыграл большую роль в классификации древних рептилий. Нижняя височная впадина свойственна синапсидным (звероподобные), верхняя – параноидным и эвриапсидным (ихтиозавры, плезиозавры). Наконец, в черепе диапсидных образовались две впадины: одна в верхней части крыши, другая – низко, в области щеки. Горизонтальную костную дугу между ними образуют заг-лазничная и чешуйчатая кости. Диапсидными были многочисленные динозавры, широко распространенные в мезозойскую эру. От ранних диапсидных рептилий произошли чешуйчатые (в частности, ящерицы). Считают, что современные виды нижнюю впадину утратили и у них сохранилась только одна боковая височная впадина.
Мозговой череп птиц по общей схеме строения весьма близок к черепу рептилий (рисунок 18). Затылочный отдел оформлен обычными четырьмя затылочными костями (основной, двумя боковыми и верхней). Затылочный мыщелок, как и у рептилий, один. Слуховая капсула закладывается, как у рептилий, из трех ушных костей, которые у взрослых птиц сливаются в одну.
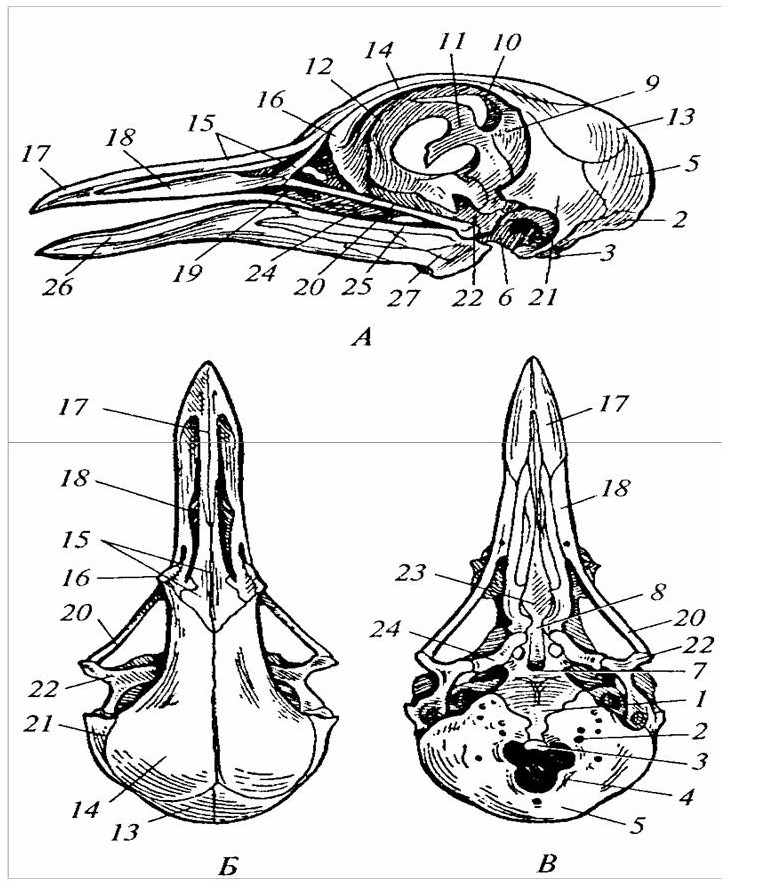
1 – основная затылочная кость; 2 – боковая затылочная; 3 – затылочный мыщелок; 4 – большое затылочное отверстие; 5 – верхняя затылочная кость; 6 – ушная; 7 – основная клиновидная; 8 – переднеклиновидная; 9 – крылоклиновидная; 10 – глазоклиновидная; 11– межглазничная перегородка; 12 – средняя обонятельная кость; 13 – теменная; 14 – лобная; 15 – носовая; 16 – слезная; 17 – межчелюстная; 18 – верхнечелюстная; 19 – скуловая; 20 – квадратноскуловая; 21 – чешуйчатая; 22 – квадратная; 23 – сошник; 24 – крыловидная; 25 – сочленовная; 26 – зубная; 27 – угловая.
Рисунок 18 – Строение черепа голубя (А – сбоку; Б – сверху; В – снизу)
Дно черепа представлено основной клиновидной и переднеклино-видной костями, а также нёбными и крыловидными костями. Крышу черепа формируют парные носовые, лобные, теменные и чешуйчатые кости В связи с тонкостью костей, ранним и прочным их срастанием череп птицы значительно облегчен. Швы между костями зарастают рано и незаметны. Объем мозговой коробки и размеры глазниц существенно увеличены. Глазницы как бы сжали щечный отдел исходно диапсидного черепа, а перекладины позади глазниц и между двумя височными впадинами исчезли. Увеличение мозговой коробки произошло за счет разрастания парных элементов крыши – лобных, теменных, чешуйчатых и комплекса боковых клиновидных костей.
Череп млекопитающих (рисунок 19) также характеризуется относительно более крупной в сравнении с рептилиями мозговой коробкой, что связано, как и у птиц, с большими размерами головного мозга. Швы между костными комплексами у млекопитающих зарастают сравнительно поздно, что обеспечивает возможность увеличения объема головного мозга по мере роста животного. У молодых зверей мозговая коробка по сравнению с лицевой частью обычно развита относительно сильнее, чем у взрослых. Число отдельных костей в черепе млекопитающего меньше, чем у ранее охарактеризованных групп позвоночных. Это обусловливается срастанием ряда костей мозгового черепа между собой: так, в единый комплекс срастаются основная, боковые и верхняя затылочные кости; сращение ушных костей приводит к образованию единой каменистой кости.
Крыловидная кость срастается с основной клиновидной костью, а глазоклиновидная кость срастается с передней клиновидной костью. Бывают случаи образования и более сложных комплексов. Единая затылочная кость несет два мыщелка для сочленения с атлантом.
Крыша черепа образована парными носовыми, лобными, теменными и непарной межтеменной костями. Такое строение крыши характерно, к примеру, для черепа собаки. Весьма изменчиво и черепе различных млекопитающих расположение ноздрей. У форм с гибким рылом (тапиры) или с хоботом (слоны, сирены) наружное отверстие ноздрей бывает сдвинуто по крыше черепа назад. У китообразных такое «дыхало» находится на вершине черепа, так как носовая область сместилась назад, а затылочные кости продвинулись вперед.
Бока черепной коробки образованы чешуйчатыми костями, от них наружу и вперед отходят скуловые отростки. Последние соединяются со скуловой костью, которая, в свою очередь, спереди сочленена со скуловым отростком верхнечелюстной кости. В итоге образуется весьма характерная для млекопитающих скуловая дуга.
Парасфеноид в дне мозгового черепа отсутствует. Дно образуют поразному сочетающиеся основная клиновидная, крылоклиновидная и крыловидная кости. В отдельных случаях они срастаются в единую клиновидную кость. Переднеклиновидные и глазокли-новидные кости тоже могут образовывать перед ней единую структуру. У приматов, грызунов и хищников под ними развивается срединная обонятельная кость, разделяющая две носовые полости. В пределах носовых полостей развиваются костные или хрящевые завитки, либо раковины. Они увеличивают поверхность обонятельных ходов.
Слуховые капсулы окостеневают несколькими центрами и в конечном счете в дне черепа формируется каменистая кость. Кроме того, здесь располагается свойственная только млекопитающим покровная барабанная кость. Она окружает среднее ухо. В слуховой барабан входит наружный слуховой проход.
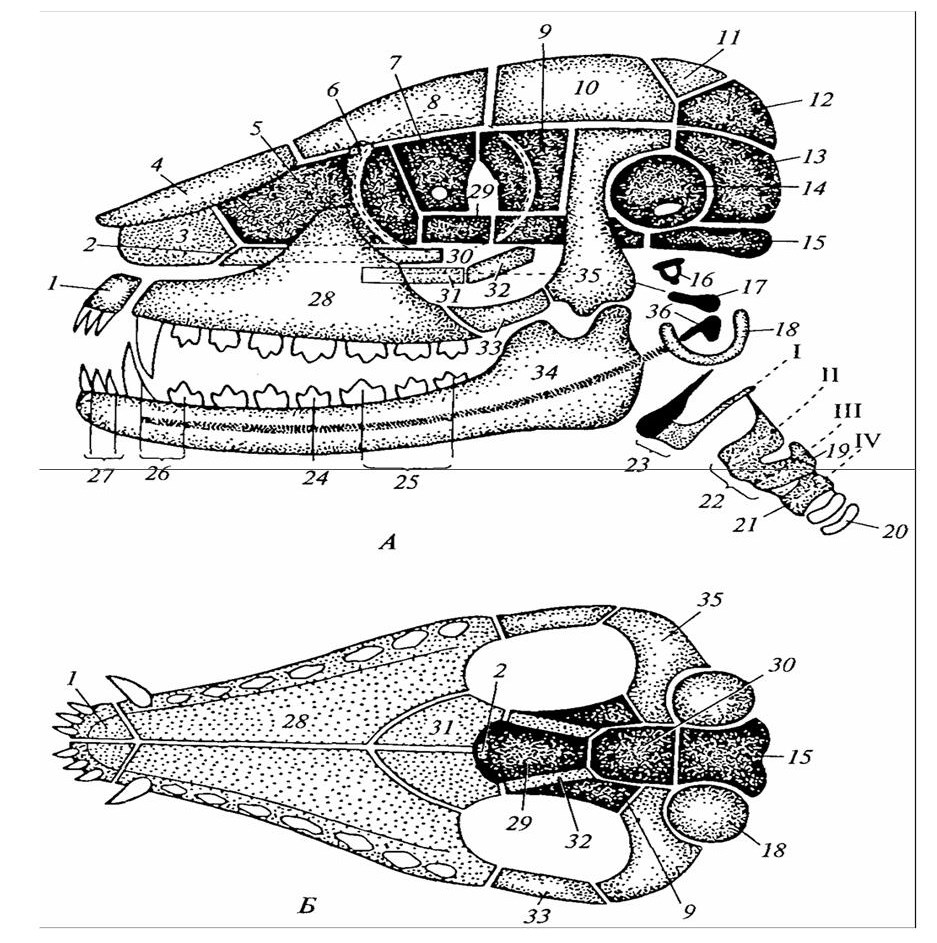
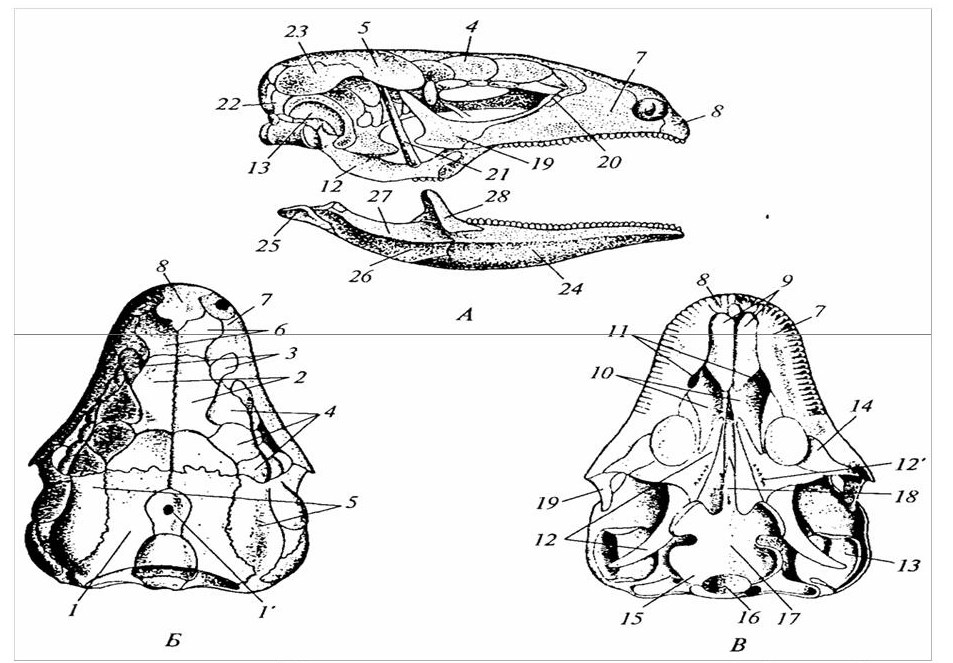
1 – межчелюстная кость; 2 – сошник; 3 – носовая перегородка; 4 – носовая кость; 5 – решетчатая; 6 – слезная; 7 – глазоклиновидная; 8 – лобная; 9 – крылоклиновидная; 10 – теменная; 11 – межтеменная; 12, 13, 15 – затылочные; 14 – каменистая; 16 – стремя; 17 – наковальня; 18 – барабанная; 19 – черпаловидный хрящ гортани; 20 – кольца трахеи; 21 – перстневидный хрящ гортани; 22 – щитовидный хрящ гортани; I – IV – остатки висцеральных дуг; 23 – рожки подъязычной кости; 24 – 27 – разные типы зубов; 28 – верхнечелюстная; 29, 30 – передняя и основная клиновидные; 31 – нёбная; 32 – крыловидная; 33 – скуловая; 34 – зубная; 35 – чешуйчатая; 36 – молоточек.
Рисунок 19 – Череп млекопитающего (А – сбоку; Б – снизу; хондральные кости темные, покровные – светлые)
Весьма примечательна изменчивость общего облика черепа разных млекопитающих. К примеру, у мелких млекопитающих его дорсальная поверхность выглядит вздутой из-за относительно большого мозга. У крупных видов на облике мозговой коробки отражается развитие височных и шейных мышц, в результате чего на поверхности черепа образуются гребни (например, сагиттальный гребень у хищных, выйный гребень у копытных). Высокая куполообразная крыша черепа слона объясняется в основном не увеличением мозга, а развитием широкой поверхности для прикрепления шейных мышц и связок, поддерживающих тяжелую голову с бивнями и хоботом.
Поверхность черепа сверху и снизу несет множество отверстий – мест выхода нервов, кровеносных сосудов, каналов (в том числе подглазничный канал, слезно-носовой канал, резцовое отверстие и т.д.).
Висцеральный череп. Висцеральный (или лицевой) череп закладывается в стенках передней части кишечной трубки. Функция его – опора ротового и жаберного аппарата. Как уже говорилось, жаберные щели у низших хордовых прежде всего выполняли функцию фильтрации, т. е. участвовали в процессе питания. С появлением жабр как органов дыхания их скелетная опора (жаберные дуги) также участвовала в фильтрующей функции. Древние круглоротые, вероятно, были микрофагами, питаясь мелкими органическими частицами, взвешенными в воде или входящими в состав отстоя. Рыбы перешли к макрофагии; этот переход сопровождался укреплением скелетной основы жаберного и ротового аппаратов. Видимо, на этой основе и возникли принципиальные отличия в схеме строения висцерального аппарата между круглоротыми, с одной стороны, и обоими классами рыб (а также всеми наземными позвоночными) – с другой. Они заключаются в том, что у круглоротых висцеральный аппарат состоит из нерасчлененных элементов, поддерживающих ротовую воронку, язык и стенки глотки в области межжаберных перегородок. Висцеральный череп такого строения несет чисто опорную функцию. У всех рыб висцеральный аппарат также располагается в межжаберных перегородках, но в каждой из них закладываются парные висцеральные дуги, состоящие из нескольких (исходно – четырех) элементов, подвижно сочлененных друг с другом. Это открывает возможность участия висцерального скелета не только в опорной функции, но и в активных движениях, связанных с захватом пищи (передние висцеральные дуги преобразуются в подвижные челюсти) и дыханием (жаберные дуги).
Эти отличия дают основание для деления всех позвоночных на два раздела: Бесчелюстные (Agnatha), к которым относятся ныне живущие круглоротые и два вымерших класса, и Челюстноротые (Gnathostomata), включающие всех остальных позвоночных. Такая неравномерность объясняется тем, что активное участие висцерального аппарата в актах питания и дыхания давало челюстноротым несомненное преимущество в прогрессивной эволюции перед придонными или плавающими микрофагами-фильтраторами с неподвижной ротовой воронкой и малоактивными дыхательными движениями, какими были ныне вымершие классы бесчелюстных. В эволюционном «соревновании» с более активными челюстноротыми смогли сохраниться лишь формы, использовавшие сосущий тип ротового аппарата для специфического способа питания: круглоротые (миноги и миксины) присасываются предротовой воронкой к телу рыб и, разрушая роговыми «зубами» языка покровы жертвы, поршневидными движениями языка высасывают жидкость тканей и кровь. Миксины при этом глубоко вгрызаются в тело жертвы, перетирая с помощью роговых «зубов» языка все мягкие ткани.
Учитывая указанные принципиальные отличия, рассмотрим строение висцерального скелета круглоротых и рыб. У круглоротых (рисунок 12) он представлен хрящами, поддерживающими ротовую воронку и язык, а также ажурной хрящевой жаберной решеткой, поддерживающей стенки тела в месте расположения жаберных отверстий. Один из непарных хрящей предротовой воронки (задний верхний хрящ) прирастает к мозговой коробке; это единственная структура, связывающая висцеральный и осевой отделы черепа.
Челюстноротые, как уже говорилось, имеют висцеральный череп, состоящий из серии висцеральных дуг, расположенных в межжаберных перегородках. Передняя пара их функционирует в качестве подвижных челюстей. У хрящевых рыб (рисунок 13) челюстная дуга представлена двумя хрящами: нёбноквадратным, играющим роль верхней челюсти, и меккелевым, образующим нижнюю челюсть. Челюсти вооружены коническими, загнутыми назад зубами, происходящими от покрывающей тело плакоидной чешуи и состоящими из дентина, одетого снаружи чехликом из прочной эмали.
Следующая висцеральная дуга – подъязычная – включает два парных хряща: подвесок (hyomandibulare) и гиоид (hyoideum); гиоиды правой и левой подъязычных дуг объединяются непарным хрящом–копулой. Функция подъязычной дуги – подвижное сочленение висцерального аппарата к осевому черепу: подвесок своим верхним концом с помощью сустава соединен со слуховым отделом мозговой коробки, а нижним – с обоими элементами челюсти в области челюстного сустава. Такой тип связи висцерального и осевого отделов черепа называется гиостилией (рисунок 20) и характерен для большинства хрящевых рыб. У некоторых примитивных акул имеется дополнительная связь в виде отростка в передней части небноквадратного хряща, причленяющегося к дну мозгового черепа. Такой тип связи носит название амфистилии. У химер (подкласс Holocephali) верхняя челюсть полностью срастается с дном осевого черепа – возникает аутостилия.
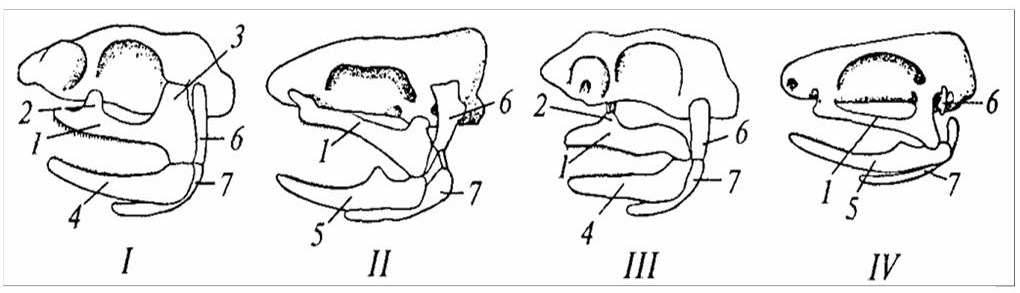
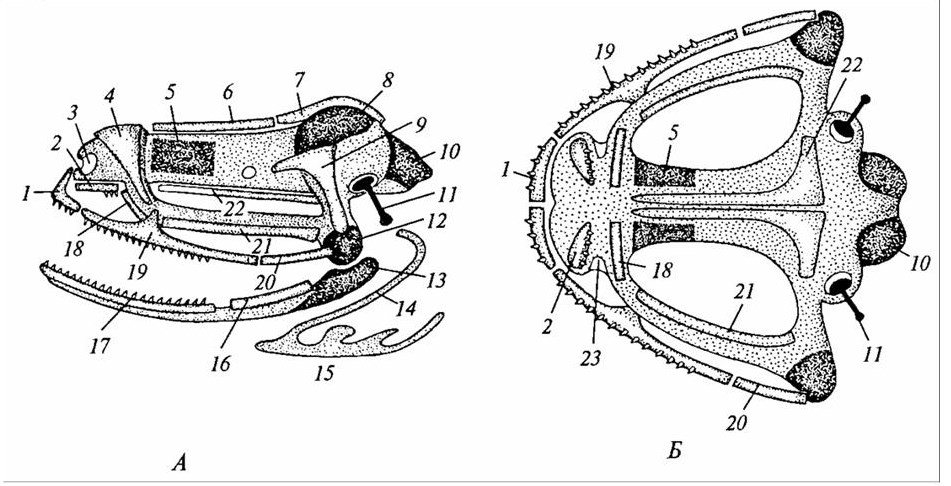
I – амфистилический череп древней акулы; II – амфистилический череп костного ганоила; III – гиостилический череп современной акулы; IV – аутостилический череп наземного позвоночного (лягушка); / – нёбноквадратный хрящ; 2, 3 – выросты нёбноквадратного хряща; 4 – меккелев хрящ; 5 – нижняя челюсть; 6 – гиомандибуляре; 7 – гиоид.
Рисунок 20 – Типы причленения висцеральных дуг к мозговому черепу
Подъязычная дуга ограничивает сзади остаток жаберной щели, которая сохраняется у хрящевых рыб в виде брызгальца, соединяющего, как и все жаберные щели, полость глотки с внешней средой. У скатов, жаберные щели смещены на брюшную сторону, расположенное на спинной стороне брызгальце, играет важную роль в дыхании.
Позади подъязычной дуги расположены однотипно устроенные жаберные дуги. У большинства хрящевых рыб их пять – соответственно числу жаберных щелей: первая расположена позади подъязычной дуги, вторая – позади 1-й жаберной и т. д. У некоторых акул число жаберных шелей может достигать 6 – 7. Каждая жаберная дуга состоит из четырех подвижно сочлененных парных элементов, объединенных снизу непарной копулой. На жаберных дугах, кроме последней, расположены органы дыхания – жабры.
У костных рыб (рисунок 14) сохраняется тот же принцип строения висцерального черепа, но висцеральные дуги составлены не хрящевой, а костной тканью. В челюстной дуге образуется большое число первичных и вторичных костей. В верхней челюсти первичные кости представлены задней крыловидной и квадратной, а в нижней – сочленованной; квадратная и сочленованная кости образуют челюстной сустав. Вторичные кости в верхней челюсти представлены предчелюстной, челюстной, небной и двумя крыловидными, а в нижней – большой зубной костью и маленькой угловой. Предчелюстная, челюстная и зубная кости формируют вторичные челюсти, снабженные зубами; кроме того, зубы имеются на небной кости.
Череп костных рыб – типично гиостилический; соединение висцерального черепа с осевым осуществляется с помощью подъязычной дуги. Эта дуга образована первичными костями, соответствующими ее отделам у хрящевых рыб: гиомандибуляре (подвесок), гиоид и непарная копула. Кроме того, к подвеску прикрепляются кости, образующие жаберную крышку, – новое по сравнению с хрящевыми рыбами образование, обеспечивающее интенсификацию дыхательных движений. Эти кости – вторичные по происхождению.
Жаберные дуги (их у костистых рыб четыре) построены по тому же плану, что и у хрящевых рыб; их элементы составлены первичными костями. Пятая жаберная дуга редуцирована и не несет жабр.
Висцеральный череп амфибий в сравнении с рыбами изменен значительно. Личинки амфибий (и некоторые взрослые) имеют жабры, соответственно у них развиваются жаберные дуги. Количество жаберных дуг уменьшается до четырех; часто они не имеют расчленения на элементы и являются цельными.
У амфибий, прошедших метаморфоз, висцеральные дуги преобразуются в скелетные элементы, характерные для наземных позвоночных. Они входят в состав подъязычного аппарата, служащего опорой языку, в состав скелета гортани и кольцевидных хрящей трахеи, а также участвуют в совершенствовании слухового аппарата. Подъязычный аппарат располагается у лягушки в дне ротовой полости между ветвями нижней челюсти и представляет собой хрящевую пластину с рожками. Ее передние рожки, загибаясь вверх, охватывают кишечную трубку и прикрепляются к слуховым капсулам.
Функцию верхних челюстей у земноводных выполняет, как и у костистых рыб, костная дуга, состоящая из покровных межчелюстных (предчелюстных), верхнечелюстных и квадратных костей.
Элементы первичного челюстного комплекса, образованного исбноквадратным хрящом, принимают участие в формировании та мозгового черепа: нёбная кость прилегает к сошнику, а квадратная кость – к чешуйчатой кости. В нижней челюсти меккелев хрящ прикрыт снаружи зубной и угловой костями.
Череп амфибий аутостилический: челюстная дуга прирастает непосредственно к мозговому черепу (рисунок 15). Подъязычная дуга не принимает участия в прикреплении челюстного аппарата к черепу. Верхний элемент этой дуги – гиомандибуляре – прекращен в маленькую кость – стремя, которая, как подвесок у рыб, верхним концом упирается в слуховую капсулу. Она лежит в преобразованной из брызгальца полости и выполняет роль слуховой косточки.
Висцеральный скелет рептилий (ящериц) в значительной степени близок к амфибиям. Верхние челюсти образованы межчелюстными и верхнечелюстными костями, несущими зубы.
Нёбноквадратный хрящ подвергается дальнейшим превращениям. Хондральная квадратная кость своим верхним отделом связана с мозговой коробкой; снизу к ней причленяется нижняя челюсть. Остальная часть нёбноквадратного хряща покрывается кожными костями – в передней части нёбными, а в серединной – крыловидными. Все эти кости участвуют в укреплении дна черепа. Появляются новые покровные поперечные кости, соединяющие крыловидные кости с верхнечелюстными, а у ящериц и гаттерий еще и хондральные столбчатые кости, соединяющие крыловидные кости с теменными.
Нижняя челюсть, гомологичная меккелеву хрящу, состоит из хондральной сочленовной кости, которая причленяется к квадратной, и ряда накладных костей, облегающих меккелев хрящ: зубной, угловой, надугловой, венечной, пластинчатой. Верхний отдел подъязычной дуги, гомологичный гиомандибуляре, представлен (как и у земноводных) слуховой косточкой – стременем. Остальные элементы висцерального скелета образуют подъязычный аппарат, состоящий из хрящевой пластинки (гомолог копулы) и трех пар рожков (гомологи гиоидов и жаберных дуг).
Весьма своеобразно строение черепа у змей. Для них характерно подвижное соединение элементов челюстного аппарата и связанных с ними костей: нёбных, крыловидных и чешуйчатых. Эта особенность позволяет змее заглатывать пищу целиком, даже довольно крупных животных. Механизм заглатывания достигается попеременным движением левой и правой половин нижней челюсти. При наблюдении за кормящейся змеей создается впечатление, что она как бы наползает на свою добычу. У ядовитых змей подвижность костей облегчает перемещение ядовитых зубов (рисунок 21).
1 – ядовитый зуб; 2 – верхнечелюстная кость; 3 – поперечная ко сть; 4 – крыловидная кость; 5 – чешуйчатая кость; 6 – квадратная кость.
Рисунок 21 – Череп гремучей змеи с закрытой (А) и раскрытой (Б) пастью
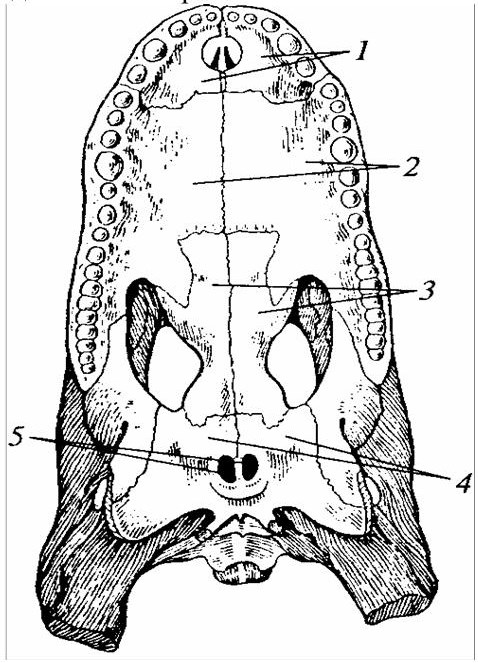
У крокодилов зубы расположены на межчелюстных, верхнечелюстных и зубных костях и сидят в альвеолах, как у млекопитающих. В связи с разрастанием нёбных отростков межчелюстных и верхнечелюстных, а также нёбных костей у крокодилов развивается костный помост – вторичное костное нёбо. Оно разделяет полость рта на верхний отдел, носоглоточный ход, и нижний отдел, собственно ротовую полость (рисунок 22). Таким образом, внутренние отверстия ноздрей (хоаны) открываются не непосредственно в ротовую полость, а в ее заднюю часть, поэтому пища, находящаяся во рту, не мешает поступлению воздуха к органам дыхания. У черепах нёбо прирастает к мозговому черепу. Квадратная кость массивная и на большом протяжении контактирует с ушным отделом, повышая прочность задней части черепа.
В висцеральном черепе птиц много общего с черепом рептилий. Верхняя челюсть включает парные межчелюстные и верхнечелюстные кости. К последним причленяются палочковидные скуловые и квадратноскуловые кости, которые, в свою очередь, соединяются с квадратными костями. В итоге образуется весьма характерная для птиц нижняя височная дуга, отграничивающая снизу глазницу и височную яму. Нижняя челюсть состоит из гомологичной меккелеву хрящу сочленовной кости и из кожного происхождения зубной, пластинчатой, угловой и венечной костей. Вместе с верхней челюстью она образует характерный для птиц клюв. Челюсти, как правило, сильно вытянуты, их кости срастаются между собой. Возникает весьма совершенный, прочный хватательный аппарат (рисунок 18). Слуховая косточка, стремечко, одна, как и у рептилий.
1 – межчелюстные кости; 2 – верхнечелюстные кости; 3 – нёбные кости; 4 – крыловидные кости; 5 – хоаны.
Рисунок 22 – Вторичное костное у нёбо у крокодила
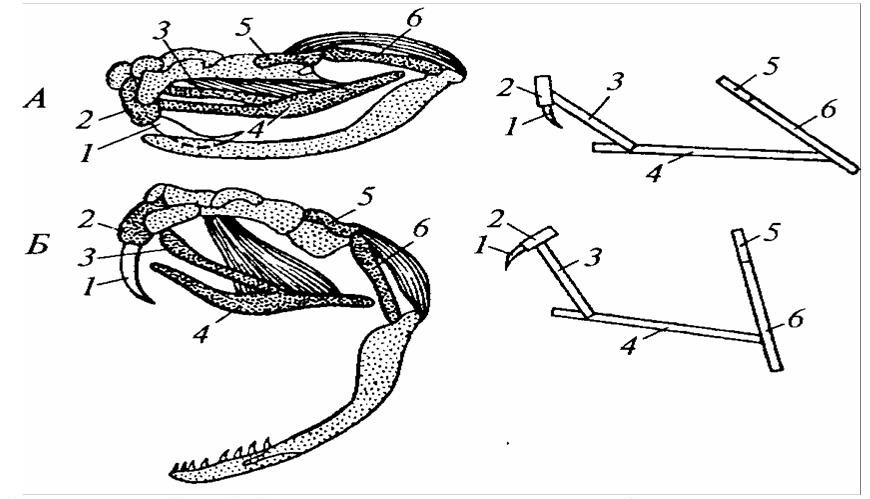
Костный подъязычный аппарат имеет вид удлиненной пластинки с очень длинными рожками, гомологичными первой паре жаберных дуг. Подъязычная пластинка участвует у птиц в движении языка при захвате пищи, а также при звукообразовании. Специалисты отмечают, что, например, у токующего глухаря во время второго колена призывной песни, когда птица глубоко втягивает язык в гортань, загнутые рожки сдвигают набухшую кровью кожу и соединительнотканную пленку на голове, отчего глухарь временно глохнет (рисунок 23).
Строение висцерального черепа млекопитающих в сравнении с рептилиями существенно меняется. Верхние челюсти состоят из парных межчелюстных и верхнечелюстных костей. Для всех млекопитающих характерно развитие вторичного костного нёба, образованного нёбными отростками межчелюстных и верхнечелюстных костей и нёбными костями. В связи с его образованием хоаны открываются не между верхнечелюстными костями, как у других наземных позвоночных, а как у крокодилов и черепах – позади нёбных костей. Такое строение нёба предотвращает закупорку хоан (т.е. перерыв дыхания) в то время, когда пищевой комок задерживается в ротовой полости при обработке пищи во рту и при глотании. Крыловидные кости уменьшаются и прирастают к дну черепа.
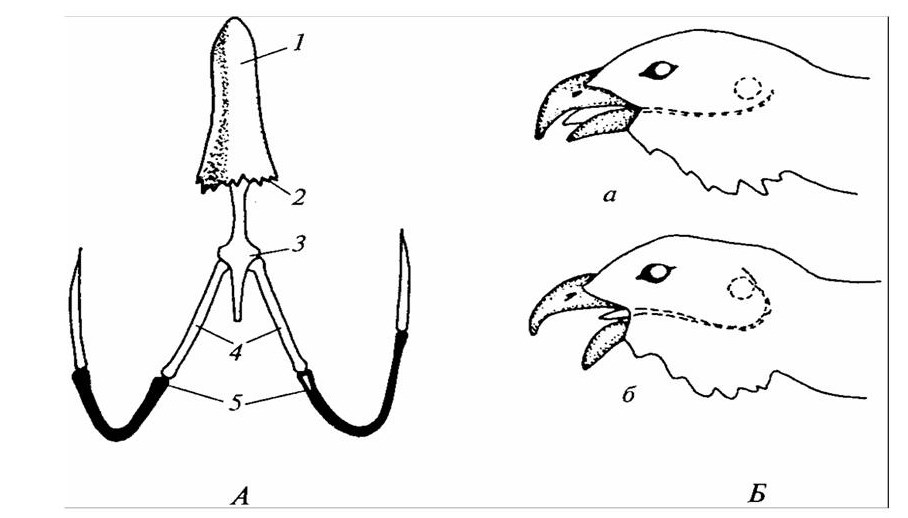
1 – язык; 2 – ороговевшие шипики языка; 3 – костная основа языка; 4 – передние рожки; 5 – задние рожки; а – обычное положение языка; б – язык втянут в глотку.
Рисунок 23 – Подъязычный аппарат глухаря (А) и его положение относительно слухового прохода (Б)
Нижняя челюсть образована только парными зубными костями, которые несут два отростка – венечный и угловой. Нижние челюсти причленяются непосредственно к чешуйчатым костям. Наиболее серьезным преобразованием становится превращение сочленовной кости в слуховую косточку – молоточек, а квадратной – в наковальню. Вместе со стременем они лежат в полости среднего уха и участвуют в звукопередаче. Наружная стенка последней, а также часть наружного слухового прохода окружены барабанной костью, гомологичной, по-видимому, угловой кости нижней челюсти прочих позвоночных. Таким образом, у млекопитающих наблюдается дальнейшее превращение части висцерального аппарата в слуховой аппарат среднего уха. Остатки подъязычного аппарата располагаются у млекопитающих в нижней части глотки и входят в состав части гортани. Функциональное значение скелета подъязычного аппарата определяется его ролью в движениях языка, связанных с захватыванием, обработкой в ротовой полости и проглатыванием пищи, с движениями глотки и гортани во время глотания, при воспроизведении звуков и т.д.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.