4.2 Типы окостенений и их биологическое значение. Локомоция
Как уже указывалось, скелет костных рыб частично или полностью состоит из костной ткани. Значение этого преобразования заключается в том, что костный скелет обеспечивает эффективное выполнение опорных функций при меньших массе и объеме. Это определяется большей, чем у хряща, прочностью и сложной внутренней структурой костей. Кроме того, при формировании костного скелета образуются новые структуры, способствующие расширению ряда функций Закладка костных элементов скелета происходит двумя путями. Костная ткань может формироваться непосредственно в исходных хрящевых закладках, постепенно полностью замещая хрящ. Так возникают первичные или замещающие кости. Кости такого типа полностью принимают на себя функции хрящевых предшественников. Второй путь формирования костей – образование очагов окостенений в соединительнотканном слое кожи (кориуме), вне связи с внутренним хрящевым скелетом. Возникшие таким путем кости называются вторичными или покровными. Они или сохраняются как самостоятельные образования например, «жучки» в коже осетровых рыб) или накладываются на хрящевую основу внутреннего скелета (например, покровные кости черепа тех же осетровых). В последнем случае хрящевая основа может полностью резорбироваться и соответствующие части скелета оказываются составленными исключительно из вторичных костей, смыкающихся с первичными. Это происходит со скелетом черепа, парных конечностей и их поясов у костистых рыб.
Появление в эволюции вторичных окостенений имеет важное значение. Возникая вне хрящевого скелета, они могут формировать новые его элементы и приобретать различный внешний вид, дополняя и расширяя функции, свойственные скелету предковых форм. Напомним, что именно вторичными костями образована часть плечевого пояса костистых рыб, прочно фиксирующая его на неподвижном осевом черепе. Это интенсифицирует опорную функцию пояса, способствует более энергичной и маневренной деятельности грудных плавников и расширяет разнообразие их функций. Из вторичных костей состоит жаберная крышка костных рыб – орган, появляющийся только в этом классе и определяющий формирование более энергичного и эффективного механизма жаберного дыхания.
Не менее важна и роль вторичных челюстей. Массивные, нерасчлененные хрящевые челюсти акуловых рыб жестко связаны с функцией открывания и закрывания рта и потому ограничены в собственно хватающей функции. Отсюда – малая степень морфоэкологического разнообразия этой группы: это преимущественно или активно плавающие хищники (акулы), или придонные формы, челюсти и зубы которых приспособлены к раздавливанию панцирей-иглокожих, раковин моллюсков (скаты) и т. п. У костистых рыб функция челюстного сустава сохраняется за первичными костями, а собственно хватательная роль переходит к комплексу костей вторичной челюсти. У разных видов они приобретают разнообразную форму и способствуют расширению способов захвата пищи, а отсюда – и дифференциации большого числа жизненных форм. Поэтому именно костистые рыбы представляют собой наиболее многочисленную, разнообразную и широко распространенную группу в современной фауне водных позвоночных.
Выше описанное строение опорно-двигательного аппарата и обтекаемая форма тела обеспечивают эффективную локомоцию в плотной среде. Боковые движения хвоста, сочетаясь с сопротивлением среды, препятствующим отклонениям головного конца в стороны, формируют силу; направленную вперед (рисунок 32). Такой способ локомоции очень экономичен. Современные исследования гидродинамики показывают, что коэффициент полезного действия при таком типе движения почти в 1,5 раза превосходит эффективность винтовых судовых движителей.
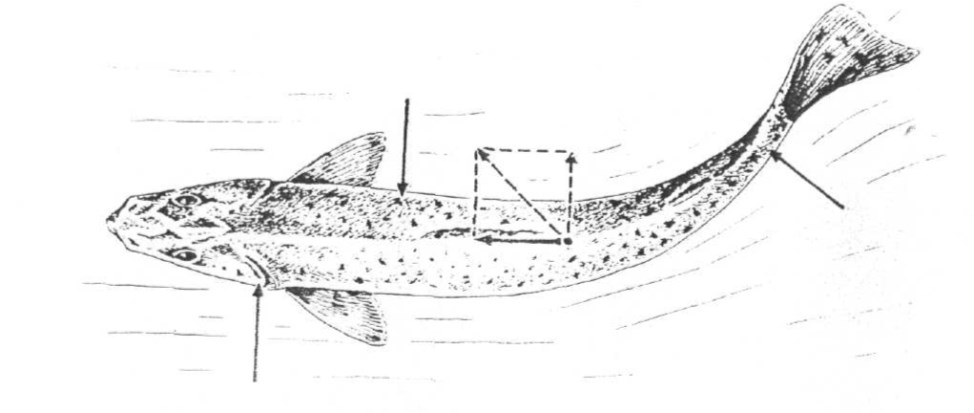
Стрелками обозначены направления сил, в сумме формирующих поступательное движение.
Рисунок 32 – Схема локомоции рыб
Таким образом, в эволюции опорно-двигательного и висцерального аппарата водных позвоночных можно отметить два важных этапа: формирование единого принципа локомоции в плотной водной среде на основе внутреннего осевого скелета и метамерной мускулатуры и дивергенция по принципу организации висцерального скелета на бесчелюстных и челюстноротых. При этом принцип организации челюстноротых открыл возможность прогрессивной эволюции, связанной с повышением энергетического метаболизма, увеличением биологической активности и расширением экологического разнообразия, тогда как эволюция бесчелюстных шла по пути специализации, в результате чего сформировалась боковая ветвь в развитии позвоночных. В пределах челюстноротых прогресс в значительной степени связан с формированием костного скелета.
У рептилий и хвостатых амфибий конечности расставлены в стороны от тела. Животные перемещаются, прижимая тело к земле, используя одновременно возможности хвоста. Такая позиция требует значительного расхода энергии на поддержание тела и движение. В истории наземных позвоночных соотношение тела и конечностей решалось различными путями. Экономичность движения была достигнута перемещением обеих пар конечностей под тело и изменением ориентации локтевого и коленного суставов.
Попытка приподнять тело над землей при четырехногой локомоции свойственна некоторым крокодилам, черепахам, ящерицам. Известно, что некоторые из них могут быстро бегать. Ящерица-василиск, например, встает на задние конечности при беге.
Бипедальная локомоция, при которой задние ноги удлинены, а колени вывернуты вперед под тело, известна уже у некоторых динозавров.Позднее это стало основой движения в наземной среде и привело у птиц и некоторых млекопитающих к бипедальной (двуногой) локомоции. Другой путь – утрата конечностей, как это свойственно змеям (и безногим амфибиям).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.