5.1 Преобразование пищеварительной системы в филогенезе
Позвоночные, как и все животные, гетеротрофы и постоянно нуждаются в притоке органических веществ извне в виде пищи. Пища необходима для построения и возобновления клеток и тканей, поддержания постоянства их химического состава и для метаболических процессов. Кроме того, пища является источником энергии, затрачиваемой организмом на различные формы деятельности. Таким образом, питание – одна из важнейших составляющих общего обмена веществ и энергии организма с окружающей средой. Важная роль этой функции послужила причиной того, что общие принципы строения пищеварительной системы и физиологии питания сложились в эволюции животных очень рано. В частности, структура и функции ферментных систем, определяющих процессы пищеварения, практически однотипны у всех животных.
Глотка. Устройство ротового аппарата определяет захват пищи и ее передвижение в последующие отделы пищеварительной системы. У бесчерепных пищевые частицы с током воды поступают через предротовую воронку, а затем рот. За ротовой полостью у этих животных следует глотка, в которой совмещаются функции дыхания и проведения пищи: вода поступает сквозь жаберные щели наружу, а пища направляется в пищевод (рисунок 34).
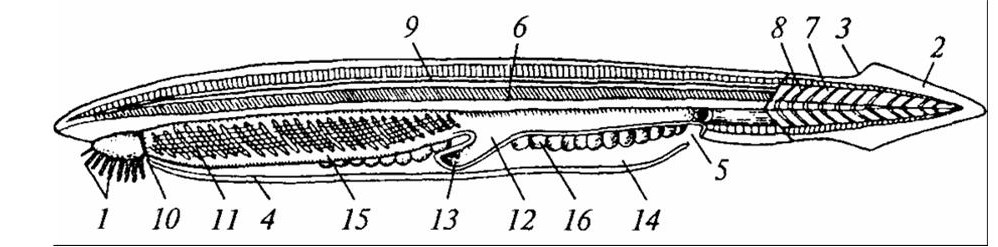
1 – предротовое отверстие, окруженное щупальцами; 2 – хвостовой плавник; 3 – спинной плавник; 4 – метаплевральная складка; 5 – атриальное отверстие; 6 – хорда; 7 – миомер (показан только в хвостовой области); 8 – миосепта; 9 – нервная трубка; 10 – парус; 11 – жаберные щели; 12 – кишка; 13 – печеночный вырост кишечника; 14 – околожаберная полость; 15 – эндостиль; 16 – половые железы.
Рисунок 34 – Продольный разрез тела ланцетника (схема)
Строение глотки, пронизанной жаберными щелями, позвоночные унаследовали от своих предков – фильтраторов. Как известно, у этих животных отделению пищи от тока воды способствует система слизистых каналов (эндостиль и др.). Круглоротые и рыбы, перешедшие к макрофагии, утратили эту систему. Остатки ее у этих групп существуют в иной функции: на основе эндостиля возникает эндокринная щитовидная железа, продуцирующая гормон тироксин.
Этот гормон, регулирующий процессы энергетического обмена и некоторые другие, появился в эволюции только с возникновением хордовых животных. Он формируется из белка тиреоглобулина путем последовательного иодирования аминокислоты тирозина. Эта реакция известна уже для эндостиля оболочников (правда, первые ее стадии – образование моно- и дииодтирозина – у них происходят в тунике). У бесчерепных ткань эндостиля также продуцирует тироксин (Т4) и его предшественник – трииодтирозин (Тз). У круглоротых эндостиль в измененном виде (так называемый гипобранхиальный мешок) сохраняется только у личинок, и в процессе их метаморфоза клетки его полностью замещаются тиреоидными фолликулами, характерными для ткани щитовидной железы. У круглоротых и у некоторых рыб щитовидная железа имеет дисперсный характер, тогда как у ряда рыб она уже хорошо выражена морфологически.
Функция разделения воды и пищи у водных позвоночных решается на иной, нежели у низших хордовых, основе. У круглоротых ток жидкой пищи и воды разделяется благодаря дифференциации глотки на пищевод и дыхательную трубку (миноги) или путем перекрытия внутренних жаберных отверстий во время питания (миксины). У рыб выносу пищевых частиц наружу препятствует система расположенных на жаберных дугах выростов – жаберных тычинок (рисунок 32). Длина и форма жаберных тычинок соответствуют составу пищи: у планктоноядных форм очень многочисленные и длинные тычинки образуют густой цедильный аппарат; у видов, питающихся более крупной добычей, они короткие и толстые, а подчас и совсем не развиты: «решетку», задерживающую пищу, образуют сами жаберные дуги.
Интересный пример экологического варьирования цедильного аппарата демонстрирует гигантская акула – Cetorhinus maximus: летом она питается планктоном и имеет густой цедильный аппарат из тонких и длинных тычинок; зимой, когда этот вид переходит на питание более крупными бентическими организмами, жаберные тычинки отпадают.
Кишечный тракт. За глоткой следует пищевод – растяжимая трубка, по которой пища перемещается в желудок. Этот отдел кишечника имеет лишь транспортную функцию; пищеварительных процессов здесь не происходит.
Эволюция пищеварительной системы, связанная с повышением эффективности переваривания и, усвоения пищи, морфологически выражается в степени дифференцирования пищеварительного тракта на морфофункциональные отделы и в возрастании общей длины кишечника, что косвенно соответствует увеличению поверхности железистого эпителия и всасывающей части кишечника. Разделение пищеварительного канала позвоночных на отделы представлено на рисунке 35. У круглоротых идущая от пищевода кишка не дифференцирована на отделы и не образует изгибов.
Некоторое расширение ее внутренней поверхности достигается с помощью продольной складки кишечного эпителия спинной стороны кишки. Уже у круглоротых появляется довольно крупная печень, лежащая непосредственно позади сердца. В печени имеется желчный пузырь, проток которого открывается в передней части кишки. Кишка проходит по верхнему краю печени, а затем опускается на брюшную сторону и тянется назад, открываясь наружу анальным отверстием.
В обоих классах рыб кишечник дифференцируется на отделы. Переваривание пищи начинается в желудке: в его стенках расположены железы, продуцирующие пищеварительные ферменты и соляную кислоту (ферменты активны в кислой среде).
У хрящевых рыб желудок U-образно изогнут; начальная, более крупная часть его называется кардиальной; а часть, следующая за изгибом, пилорической. Позади этой части находится очень короткая тонкая кишка, в просвет которой открываются протоки поджелудочной железы, а также желчный проток, идущий из крупной трехлопастной печени. В тонком кишечнике продолжается переваривание пищи с участием ферментов, вырабатываемых в стенках кишки и в поджелудочной железе.
Морфологически выраженная поджелудочная железа появляется лишь у хрящевых рыб. Это сложная железа, работающая и как экзокринная (продукция пищеварительных ферментов), и как эндокринная (продукция инсулина, глюкагона и некоторых других гормонов). Инсулин и глюкагон – гормоны – антагонисты, регулирующие уровень сахара в крови: инсулин способствует резервированию углеводов в виде гликогена мобилизации глюкозы путем распада гликогена, а глюкагон – мобилизации глюкозы путем распада гликогена в печени. Эндокринная функция железы связана с особыми клетками островков Лангерганса. У круглоротых ткань поджелудочной железы разбросана островками вдоль кишечника и внешне не выражена, хотя функциональная ее роль проявляется как в пищеварении, так и в продукции гормонов (прежде всего инсулина).
Разжиженная пищевая масса из тонкой кишки попадает в толстую, где заканчиваются процессы пищеварения, и происходит всасывание питательных веществ и воды. Толстая кишка хрящевых рыб имеет своеобразное строение: ее внутренние стенки образуют спиральный клапан, существенно увеличивающий поверхность контакта кишечного эпителия и пищевой массы. Этим компенсируется небольшая длина кишечника. Кроме того, спиральный клапан увеличивает время прохождения пищи через кишечник и таким образом – эффективность усвоения пищи. Толстый кишечник переходит в короткую прямую кишку, вырост которой у хрящевых рыб образует ректальную железу – орган солевого обмена. Прямая кишка впадает в клоаку – расширенный, задний отдел кишечной трубки, в который открываются анальное отверстие и отверстия мочевых в половых протоков.
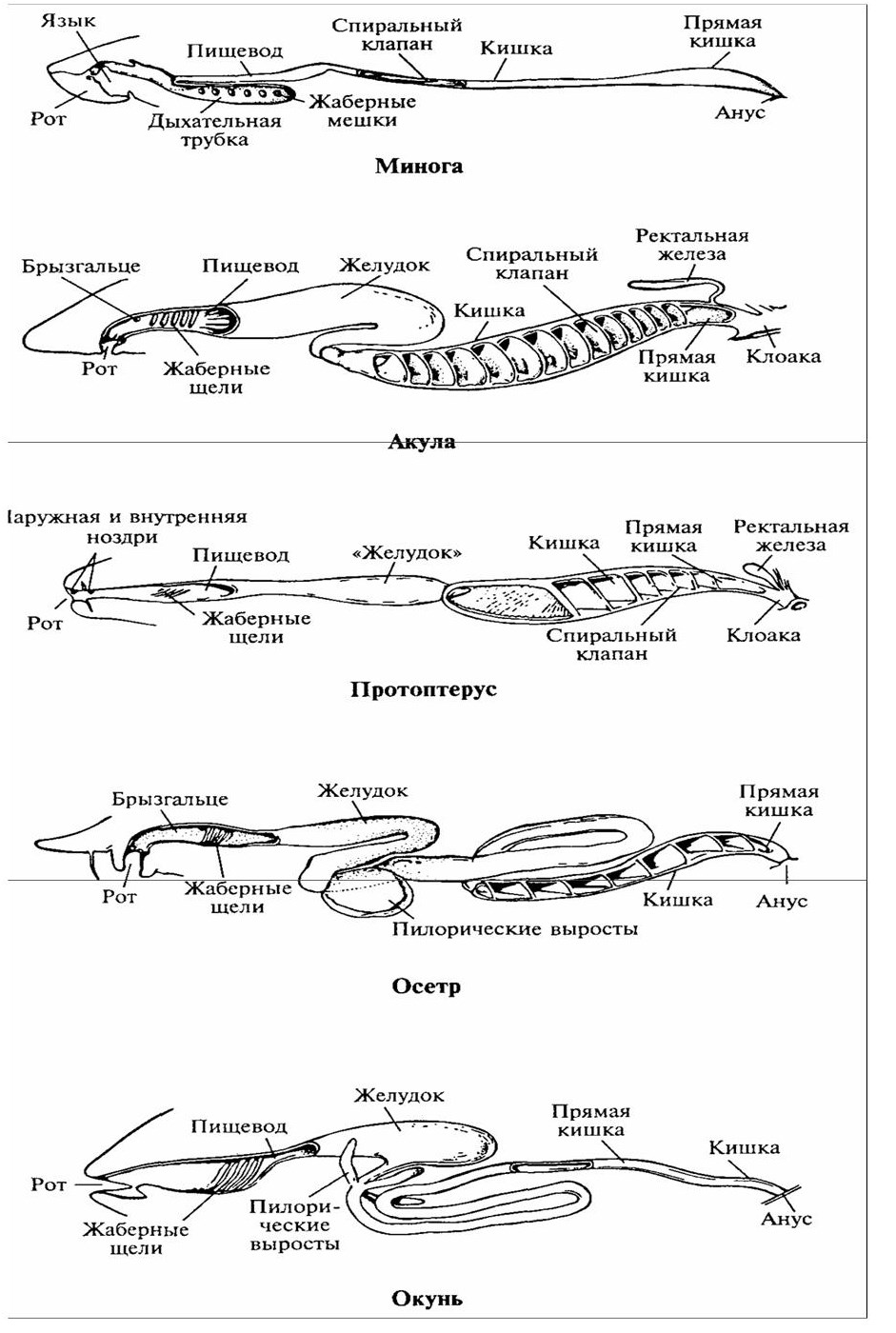
Рисунок 35 – Схема строения пищеварительной системы разных животных (по А.Ромеру, Т.Парсонсу, 1992)
Сходно подразделение пищеварительной системы на отделы у костных рыб. Желудок у некоторых “мирных” костистых рыб может и не выделяться морфологически, но выражен функционально. Позади желудка, на границе с тонкой кишкой, у многих видов имеется несколько замкнутых выростов – пилорические придатки. Здесь тоже происходит переваривание белков, и всасывание. Тонкий кишечник длинный, образует петли, благодаря которым увеличивается его общая поверхность. Первая петля тонкой кишки формирует двенадцатиперстную кишку, в которую впадают протоки поджелудочной железы и желчные протоки, поэтому функционально этот отдел отличается от остальной части тонкой кишки. Толстый кишечник у костистых рыб морфологически почти не выделяется, но у низших костных рыб, как и у акул, имеется спиральный клапан. Клоаки у костных рыб (кроме двоякодышащих) нет, и кишечник открывается анальным отверстием прямо наружу.
Подразделение кишечного тракта на пищевод, желудок, кишечник (с двенадцатиперстной кишкой в его начале, толстую и прямую кишки) сохраняется у всех позвоночных животных, лишь в деталях строения.
У земноводных в связи с их адаптацией к водной и воздушной средам обитания и развитием органов дыхания происходит значительная перестройка головной кишки, которая приводит к обособлению ротовой и носовой полостей, редукции жаберного аппарата и развитию органов газообмена – легких. Средняя кишка удлиняется, вентральная брыжейка редуцируется и образуются петли кишечника, подвешенные на дорсальной брыжейке. Задняя кишка представлена клоакой и небольшим дорсальным выпячиванием – слепой кишкой.
У пресмыкающихся жаберный аппарат можно проследить лишь на ранних стадиях эмбриогенеза. У них происходит образование вторичного твердого, а у высших пресмыкающихся (крокодилы) – и мягкого неба; в желудке выделяется мышечная часть, получившая наибольшее развитие у птиц; слепая кишка достигает значительных размеров.
У млекопитающих отделы пищеварительного тракта достигают наибольшей степени развитости. Для них характерны полное разделение ротовой полости от носовой; дифференциация и специализация жевательного аппарата и особенно зубной системы; подразделение средней кишки на двенадцатиперстную, тощую и подвздошную, а толстой, полностью отделившейся от мочеполового аппарата, на ободочную, слепую и прямую кишки.
В зависимости от адаптации к определенным условиям существования и типа питания (травоядные, плотоядные, всеядные, насекомоядные, плодоядные и т. д.) различные отделы и органы пищеварительного аппарата приобретают характерные видовые различия. Так, у растительноядных пища, состоящая из клетчатки, требует более длительного времени для переваривания, что привело к значительному увеличению толстого отдела кишечника с его характерными расширениями (лошадь, зайцеобразные) или к увеличению количества камер желудка от двух (хомяки), трех (верблюды) до четырех (крупный и мелкий рогатый скот, олени, лоси) и более (некоторые китообразные). У плотоядных (собака, кошка), пища которых состоит из кормов животного происхождения, пищеварительный тракт значительно короче. Всеядные (свинья) занимают между травоядными и плотоядными промежуточное положение (таблица 2).
Таблица 2 – Длина отделов кишечника у домашних млекопитающих
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.