Сообщество превращается в организм
Сообщество превращается в организм
«Планета микробов» жила и успешно развивалась в течение миллиарда или более лет, и за это время ее микроскопические обитатели добились немалых успехов. Высшим достижением этого этапа эволюции стали сложные микробные сообщества — бактериальные маты, о которых говорилось в предыдущей главе.
По уровню целостности бактериальный мат приближается к настоящему организму. Приближается, но все-таки не достигает этого уровня. В какой-то момент эволюция микробных сообществ словно наткнулась на невидимую преграду — и вдруг оказалось, что для дальнейшего эволюционного прогресса не хватает чего-то очень важного. Забегая вперед, скажу, что прокариоты так и не смогли преодолеть этот рубеж и дать начало настоящим многоклеточным организмам.
Все развитие прокариотной биосферы строилось на основе одного и того же базового «кирпичика», породившего бесчисленное множество собственных модификаций. Этим кирпичиком была прокариотическая клетка. При всех бесспорных преимуществах этой универсальной живой единицы, при всем ее умении приспосабливаться почти к любым условиям и извлекать энергию чуть ли не из любого химического процесса в строении прокариотической клетки есть ряд непреодолимых ограничений.
Главное из них связано с отсутствием клеточного ядра. Наследственный материал прокариот (обычно это единственная кольцевая молекула ДНК — кольцевая хромосома) находится прямо в цитоплазме, то есть, образно говоря, в бурлящем биохимическом котле, где происходит обмен веществ и осуществляются тысячи химических реакций. В такой неспокойной обстановке очень трудно развить сложные и эффективные молекулярные механизмы регуляции работы генов. У эукариот функционирование генома регулируется сотнями и тысячами специализированных белков, а также особыми регуляторными РНК и другими молекулами. Весь этот управляющий аппарат находится в ядре клетки, и ядерная оболочка надежно защищает его от бурной биохимической деятельности цитоплазмы. Тонкая регуляция работы генов обеспечила эукариотам качественно иной уровень пластичности. Самое главное, она позволила клетке радикально менять свои свойства, структуру и облик, не изменяя при этом сам геном, а только усиливая или ослабляя работу разных генов. Именно эта пластичность позволила эукариотам в конце концов стать многоклеточными в строгом смысле этого слова. Ведь в настоящем многоклеточном организме не просто много клеток, а много разных типов клеток (покровные, мышечные, нервные, половые и т. д.) Однако геном у них у всех один и тот же!
Разумеется, механизмы регуляции работы генов есть и у прокариот, но они проще и работают менее эффективно. Некоторые «высшие» прокариоты, такие как цианобактерии, могут даже иногда менять строение своих клеток, не меняя генома (пример — образование гетероцист у нитчатых цианобактерий, о чем говорилось в главе «Планета микробов»). Но эти зачаточные способности к адаптивным модификациям клеток не идут ни в какое сравнение с тем, что наблюдается у эукариот.
Второй важный конструктивный «недостаток» прокариотической клетки состоит в отсутствии окруженных мембранами органелл. Или, как говорят биологи, в отсутствии компартментализации внутренней среды клетки: цитоплазма прокариот не подразделена на отсеки — «компартменты». А ведь далеко не все биохимические процессы, которые могли бы оказаться полезными клетке, можно осуществлять в едином «общем котле», каковым является цитоплазма прокариот. Представьте себе, как усложнилась бы работа химика, если бы в его распоряжении имелась лишь одна-единственная пробирка! Прокариоты, конечно, пытались по-своему преодолеть этот недостаток. Если присмотреться к прокариотической клетке повнимательнее, можно заметить, что в распоряжении бактерий на самом деле не одна «пробирка», а две. В роли второй выступает так называемое периплазматическое пространство, то есть область снаружи от клеточной мембраны (здесь обычно расположено толстое, рыхлое, сложно устроенное многослойное образование — «клеточная стенка»). В толще клеточной стенки могут протекать химические процессы, не совместимые с теми, которые идут в цитоплазме. Но и двух пробирок все-таки мало для хорошей химической лаборатории! Эукариоты в этом отношении дадут прокариотам сто очков вперед. Внутренняя среда эукариотической клетки подразделена двойными и одинарными мембранами на множество разнообразных отсеков-«компартментов» (ядро, митохондрии, пластиды, эндоплазматическая сеть и т. д.).
Именно из-за этих ограничений прокариоты с самого начала так сильно тяготели к симбиозу. В микробном сообществе разные виды микробов в функциональном отношении соответствуют разным отсекам эукариотической клетки. Сообща микробы могут достичь гораздо большего, чем поодиночке.
Однако как бы ни были тесны связи в симбиотическом содружестве микробов, сообщество не становится настоящим целостным организмом, потому что оно не может размножаться как единое целое (подробнее об этом см. врезку «Появлению многоклеточности мешают обманщики» в главе «Рождение сложности»). Каждый из микробов сохраняет способность «сбежать» из сообщества и перейти к самостоятельной жизни, если где-то сложатся подходящие для этого условия; каждый размножается сам по себе и имеет свой собственный индивидуальный геном, который и передает своим потомкам. Поэтому естественный отбор продолжает действовать на уровне отдельных клеток, а не на уровне сообщества. Любая мутация, повышающая жизнеспособность данного конкретного микроба, будет поддержана отбором даже в том случае, если это вредно для сообщества как целого. Чаще всего вред для сообщества будет означать и вред для данного микроба, но, увы, не всегда — иначе не было бы на свете паразитов и обманщиков. Поэтому эволюция прокариот в основе своей остается эгоистической.
Чтобы преодолеть эти ограничения, прокариотическим клеткам нужно было сделать еще один шаг — вполне естественный и логичный — в сторону дальнейшего усиления интеграции, сплоченности сообщества. Они должны были по-настоящему слиться в единый организм, отказаться от своей клеточной индивидуальности и объединить свои персональные хромосомы в один большой общий геном.
Именно это и случилось в начале протерозойского эона (вероятно, около 2,0–2,2 млрд лет назад)[27]. Сообщество прокариот, слившееся в единый организм — эукариотическую клетку, — стало новым базовым «строительным блоком», усовершенствованным «кирпичиком» в том великом конструкторе, из которого эволюция по сей день продолжает собирать новые формы жизни.
—————
Воскрешенные белки рассказывают о климате древнейших эпох.[28] Для реконструкции древнейших этапов эволюции очень важно хотя бы примерно представлять, в каких условиях происходили эти события. Одним из самых спорных является вопрос о том, каким был климат на планете в течение архейского и протерозойского эонов.
Как правило, о климате столь отдаленных эпох судят по изотопному составу углерода, кислорода, кремния, серы и других элементов в осадочных породах земной коры. Эти данные порой допускают неоднозначное толкование. Иногда предположения о протерозойских температурах базируются на присутствии в породах ископаемых остатков тех или иных бактерий, но и эти данные считаются не вполне надежными и обычно вызывают массу возражений. Как и в случае с абсолютными геохронологическими датировками[29], для повышения надежности и точности палеоклиматических реконструкций огромное значение имеет привлечение разных источников данных. Если каждый метод в отдельности не очень надежен, нужно использовать сразу несколько независимых методов и смотреть, сходятся ли результаты. Вплоть до самого последнего времени палеоклиматические реконструкции, относящиеся к архею и протерозою, казались весьма сомнительными. Однако в феврале 2008 года в журнале Nature появилась статья американских исследователей, которая сильно изменила ситуацию к лучшему.
Ученые из Фонда прикладной молекулярной эволюции, факультета биохимии и молекулярной биологии Флоридского университета и компании «ДНК 2.0» (DNA 2.0 Inc) провели многоступенчатое исследование, в результате которого удалось получить весьма правдоподобную реконструкцию температуры земной поверхности в архейские и протерозойские времена, то есть 3,5–0,5 млрд лет назад.
Ученые подошли к проблеме с совершенно новой и неожиданной стороны. Их идея состояла в том, чтобы восстановить белки древнейших бактерий и проверить, к каким температурам эти белки лучше всего приспособлены — благо современная молекулярная биология уже способна выполнить такие удивительные маневры, как реконструирование исчезнувших белков. Тогда диапазон температур, в которых реконструированные белки будут устойчивы, как раз и покажет температурные условия, к которым были приспособлены бактерии.
В качестве белков-индикаторов решено было восстановить ферменты, участвующие в последовательном присоединении аминокислот к синтезируемой молекуле белка (этот процесс называется элонгацией и представляет собой основной этап трансляции). Ферменты этого этапа трансляции называются факторами элонгации. Они должны всегда присутствовать в клетке в большом количестве, пока клетка жива, и всегда должны быть наилучшим образом приспособлены к окружающей среде, иначе белки в клетке будут синтезироваться слишком медленно. Исследователи остановили свой выбор на одном из трех основных факторов элонгации, присутствующих у прокариот, — EF-Tu (elongation factor thermo-unstable).
Исходя из строения факторов элонгации и, соответственно, кодирующих их генов у разных групп современных бактерий, а также у хлоропластов и митохондрий были реконструированы наиболее вероятные предковые гены факторов элонгации, соответствующие основным узлам (точкам ветвления) эволюционного древа бактерий. Эта стандартная статистическая задача сейчас легко решается: банки данных с расшифрованными генными последовательностями, в том числе и гены белков — факторов элонгации, находятся в открытом доступе. Для реконструкций были использованы две альтернативные схемы эволюции бактерий, так что генные последовательности предковых белков получились немного разные. Затем эти гипотетические белки изготовили в лаборатории: собрали кодирующие их гены, внедрили их в клетки кишечной палочки, и бактерия вынужденно синтезировала необходимые ученым белки. После чего можно было спокойно изучать термостойкость этих воскрешенных белков.
Выяснилось, что, чем раньше разошлись эволюционные ветви, то есть чем раньше жил общий предок соответствующей группы бактерий, тем более термостойким был его фактор элонгации. Самые древние общие предки, жившие в раннеархейские времена (3,5 млрд лет назад), были приспособлены к температуре около 60–70 °C. Самые молодые, жившие в конце протерозоя (550 млн лет назад), предпочитали гораздо более прохладный климат — 37–35 °C.
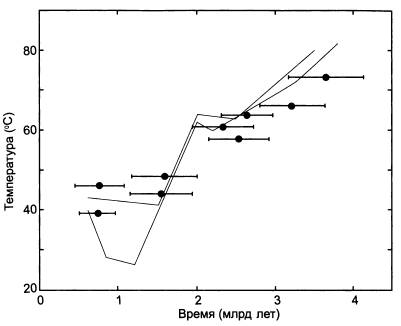
Снижение воссозданных температур плавления факторов элонгации по мере приближения к современности. По горизонтальной оси — время в млрд лет назад. Горизонтальные отрезки — это предполагаемое время существования общего предка той или иной группы бактерий по молекулярным часам с диапазоном возможных ошибок. Температура плавления экспериментальных белков определена без ошибок (для современных бактерий показано, что они лучше всего растут при температуре примерно на 2 градуса ниже, чем температура плавления белка EF-Tu, характерного для данной бактерии). Серыми линиями показан тренд снижения температуры океанов по изотопам кислорода (светлая и темная линии отражают оценки, полученные разными авторами на основе анализа разных горных пород).
Это означает, что на заре земной жизни бактерии жили примерно в таких же условиях, какие сейчас существуют в горячих источниках, если к этому прибавить ультрафиолет и отнять кислород (пока в атмосфере не было кислорода, не было и озонового слоя, задерживающего ультрафиолетовое излучение).
Что касается цианобактерий, то они, как выяснилось, изначально жили при температуре около 64 °C. Примерно к таким же температурам приспособлены и современные цианобактериальные маты, живущие в горячих источниках. Общий предок всех митохондрий, судя по свойствам воскрешенных белков, жил при температуре 51–53 °C; общий предок всех бактерий — вообще при 64,8-73,3 °C.
Самое главное, что полученные результаты почти полностью совпали с теми графиками, которые были получены ранее по изотопам кислорода и кремния. Совпадение результатов, полученных различными методами и на основе различных данных, всегда обнадеживает — это, пожалуй, единственный критерий правдоподобия подобных реконструкций.
(Источник: Eric A. Gaucher, Sridhar Govindarajan, Omjoy K. Ganesh. Palaeotemperature trend for Precambrian life inferred from resurrected proteins // Nature. 2008. V. 451. P. 704–707.)
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
2.4. Влияние вирусов на организм человека
2.4. Влияние вирусов на организм человека Большую группу паразитов человека, животных и растений образуют вирусы. Они могут вызывать ряд тяжелых заболеваний, таких как натуральная и ветряная оспа, полиомиелит и др. Вирусы изучаются специальной наукой –
В начале было сообщество?
В начале было сообщество? Многие биологи полагают, что все разнообразие жизни на нашей планете происходит от единственного исходного вида — «универсального предка» по имени Лука (об этом прародителе всего живого на Земле мы уже упомянули в начале главы). Другие, в том
ГЛАВА 3 СООБЩЕСТВО ТЕРМИТОВ
ГЛАВА 3 СООБЩЕСТВО ТЕРМИТОВ ТЕРМИНЫ-ОРАКУЛЫ Насекомые, живущие единой общиной, — муравьи, осы, пчелы и термиты — всегда вызывали у людей удивление. Это нашло отражение в многочисленных мифах, легендах и преданиях. В Европе особенно таинственным казалось поведение пчел,
ГЛАВА 3. СООБЩЕСТВО ТЕРМИТОВ
ГЛАВА 3. СООБЩЕСТВО ТЕРМИТОВ Возможность провести исследование термитных колоний появилась у меня в августе 1998 г., когда я три недели провел в Бразилии, работая в одном экологическом институте, расположенном во влажном тропическом лесу на побережье Атлантического
Плавник превращается в лапу
Плавник превращается в лапу Как мы уже говорили, растения совершили «подвиг»: они вышли из воды и заселили сушу. Это были псилофиты — странные и слабые растеньица, не выше четверти метра, немного похожие на плауны. Выполнив эту величайшую из задач природы, псилофиты
10. СООБЩЕСТВО ВЛАДЕЮЩИХ ЯЗЫКОМ ШИМПАНЗЕ
10. СООБЩЕСТВО ВЛАДЕЮЩИХ ЯЗЫКОМ ШИМПАНЗЕ Ключевые свойства – Уошо, Люси и Элли Двойственность – да Продуктивность – да Произвольность – да Взаимозаменяемость – да Специализация – да Перемещаемость – да Культурная преемственность – да Вот точка зрения Футса на
21. Организм и среда. Экологические факторы
21. Организм и среда. Экологические факторы Вспомните!Что изучает наука экология?Какие экологические факторы вам известны?Как организм состоит из отдельных клеток, которые в сумме создают некое единство, обладающее новыми качествами, так и биосфера состоит из своих
СООБЩЕСТВО
СООБЩЕСТВО Сообщество — это совокупность видов, обитающих в определенном месте. Иногда отдельное местообитание имеет вполне определенные границы, как, например, пруд, иногда же нет — в степи одни виды сменяют другие по мере изменения градиента влажности. Конечно, любые
СООБЩЕСТВО: СТРУКТУРА
СООБЩЕСТВО: СТРУКТУРА От каких процессов зависят численность и разнообразие видов в том или ином сообществе? Какие процессы определяют структуру сообщества? До какой степени структура сообщества определяется локальными процессами, происходящими внутри сообщества
СООБЩЕСТВО: ФОРМИРОВАНИЕ
СООБЩЕСТВО: ФОРМИРОВАНИЕ Как из всех присутствующих в данном географическом регионе видов формируется сообщество? Является ли сообщество просто «сборищем» всех возможных для данного типа местности видов или его образование подчиняется каким-то правилам комбинации
СООБЩЕСТВО: ЧЕРЕДОВАНИЕ СТАБИЛЬНЫХ СОСТОЯНИЙ
СООБЩЕСТВО: ЧЕРЕДОВАНИЕ СТАБИЛЬНЫХ СОСТОЯНИЙ В последние годы увеличилось число случаев «цветения» озер — усиленного размножения в них водорослей. Причиной такого «цветения» служит повышение в них концентрации биогенных элементов, главным образом азота и фосфора.
Глава 3. Организм
Глава 3. Организм ТЕМЫ• Организм – единое целое. Многообразие организмов• Обмен веществ и превращение энергии• Размножение• Индивидуальное развитие (онтогенез)• Наследственность и изменчивость• Основы селекции. БиотехнологияМысленно поднимаясь по лестнице