Вездесущий симбиоз
Вездесущий симбиоз
Когда-то симбиоз считался сравнительно редким явлением — скорее курьезом, чем правилом. Когда в 60-е годы XIX века было обнаружено, что лишайники представляют собой симбиотические комплексы из грибов и водорослей, ученые страшно удивились (надо же, какие причуды бывают у матушки-природы!). С тех пор многое изменилось. Уже в начале XX века отдельные выдающиеся мыслители предполагали, что симбиоз и кооперация могут играть огромную роль в развитии жизни на Земле. Хотя «организмоцентрический» подход в биологии по-прежнему господствует, сегодня ученые ясно понимают, что симбиоз — это магистральный путь эволюции, без которого прогрессивное развитие жизни на Земле было бы крайне затруднено, если вообще возможно.
По-настоящему «автономный организм», сформировавшийся и живущий без всякого участия каких-либо симбионтов, в природе еще надо поискать. Большинство живых существ, населяющих планету, в действительности являются «сверхорганизмами» — сложными симбиотическими комплексами.
Человек не исключение. Каждая наша клетка получает необходимую ей энергию от митохондрий — потомков симбиотических бактерий. Многие из наших генов получены нами от вирусов, всевозможных «эгоистических» фрагментов ДНК и мобильных генетических элементов (см. главу «Наследуются ли приобретенные признаки?»). Наш метаболизм во многом определяется многочисленными микробами, составляющими кишечную флору. И даже если заглянуть внутрь любого из этих микробов, то и там мы найдем сожителей-симбионтов (плазмиды, фаги, транспозоны).
—————
Кишечная микрофлора превращает человека в «сверхорганизм». В молекулярно-биологических исследованиях главный залог успеха — это удачный выбор объекта и методики. Новые эффективные методы, разработанные для решения конкретных задач, могут вдруг оказаться весьма полезными и в совершенно других областях. Именно это и произошло в данном случае. Американские биологи нашли необычное применение новейшим методикам, разработанным для «прочтения» геномов различных организмов. Ученые применили эти методы к ДНК, экстрагированной из человеческих фекалий, с целью получения общей характеристики кишечной микрофлоры.
По имеющимся оценкам, в кишечнике взрослого человека присутствует более 1 кг микроорганизмов, относящихся к сотням различных видов. В точности их видовой состав неизвестен. Микробиологи знают «в лицо» лишь несколько десятков типичных представителей, которых можно вырастить на искусственных средах. Как выяснилось сравнительно недавно (и это открытие стало шоком для микробиологов), большинство существующих в природе микроорганизмов на искусственных средах не растет. Таких микробов называют некультивируемыми. В человеческом кишечнике они тоже, скорее всего, составляют большинство. Что-либо узнать об этих микробах удается лишь по нуклеотидным последовательностям ДНК в пробах, взятых из естественных сред.
Исследователи выделили ДНК из фекалий двух человек, не принимавших целый год перед этим никаких лекарств, и провели масштабную работу по секвенированию — определению нуклеотидной последовательности фрагментов ДНК. Отсеквенированные кусочки затем собирались в более длинные фрагменты на основе наличия перекрывающихся участков. В итоге получилось около 74 тысяч неповторяющихся кусков общей длиной свыше 78 млн пар нуклеотидов. Для сравнения — в геноме человека свыше 3 млрд пар нуклеотидов, в геноме одной бактерии обычно 2–5 млн п.н.
Разумеется, исследователи не рассчитывали получить полные геномы всех кишечных микробов. Для этого им пришлось бы приложить на несколько порядков больше усилий. Они лишь хотели получить общее представление о разнообразии микробного сообщества, его структуре и, главное, обмене веществ. А для этого полученной выборки геномных последовательностей оказалось вполне достаточно.
Такой радикальный подход к изучению сообществ — свалить всех в одну кучу, истолочь и отсеквенировать — получил даже специальное название — метагеномный анализ.
—————
Метагеномный анализ применяется в тех случаях, когда требуется выяснить, какие микроорганизмы присутствуют в пробе. Суть метода в тотальном выделении из образца (например, из морской воды, почвы, тканей какого-нибудь животного или из содержимого кишечника) всех молекул ДНК, какие попадутся. ДНК секвенируют (определяют последовательности нуклеотидов) и по этим последовательностям выясняют, какие существа присутствуют в пробе. Важное достоинство такого подхода в том, что он позволяет обнаружить и охарактеризовать микробов, не поддающихся культивированию в лаборатории (а таких среди микробов большинство). Генетические базы данных сегодня уже достаточно представительны, чтобы по набору выделенных из пробы генов можно было определить, какие организмы присутствуют в пробе, даже если эти организмы до сих пор не были известны науке. Сравнивая найденные гены с известными, можно выяснить не только чьими родственниками являются эти существа, но и как они живут и чем дышат.
—————
На следующем этапе в отсеквенированных последовательностях искали гены с известными функциями и пытались определить, какие фрагменты принадлежат бактериям, а какие — археям. Для этого последовательности сравнивались с известными, то есть внесенными в базы данных генами бактерий и архей, а также с их полными геномами. Отдельно анализировались гены рибосомной РНК (16S), по которым традиционно проводят классификацию микробов.
В «явном виде» по генам рибосомной РНК удалось идентифицировать лишь 72 разновидности бактерий (из них 60 некультивируемых и 16 новых для науки) и один вид архей-метаногенов, однако авторы обосновали статистически, что, если бы работа по секвенированию ДНК из тех же самых проб была продолжена, число выявленных разновидностей микробов составило бы не менее 300.
Оказалось, что в кишечной микрофлоре резко повышена доля генов, имеющих отношение к метаболизму полисахаридов растительного происхождения, некоторых аминокислот и витаминов, а также к метаногенезу. На основе проведенного анализа авторы выделили наиболее важные метаболические функции, которые выполняют микробы в человеческом кишечнике. Это прежде всего переваривание растительных полисахаридов, которые не могут перевариваться ферментами, закодированными в геноме человека. С этими трудноусваиваемыми углеводами расправляются в основном бактерии-бродильщики, выделяющие в качестве конечных продуктов обмена низкомолекулярные органические кислоты. Однако то, что для бактерий-бродильщиков является отходами жизнедеятельности, для человека — вполне съедобные вещества, которые активно всасываются кишечным эпителием. По имеющимся оценкам, из этого необычного источника люди получают около 10% калорий (эта оценка справедлива для приверженцев типичной «европейской» диеты). Задумайтесь, что это значит: не бактерии питаются нашими отходами, как кто-то мог бы подумать, а как раз наоборот!
Кроме съедобных для человека веществ бактерии-бродильщики выделяют в качестве побочного продукта еще и молекулярный водород (H2), который вреден для них самих и препятствует их росту и жизнедеятельности. Чтобы процесс переваривания растительных полисахаридов шел эффективно, кто-то должен постоянно утилизировать образующийся водород. Именно этим и занимаются археи-метаногены. В ходе метаногенеза поглощаются водород и углекислый газ и выделяется метан.
В «совокупном геноме» кишечной флоры сильно повышено процентное содержание генов, связанных с синтезом незаменимых аминокислот и витаминов. Микробы сильно облегчают человеку жизнь, производя значительные количества этих необходимых нам веществ. Кроме того, кишечная флора располагает большим арсеналом ферментов для обезвреживания токсичных веществ, присутствующих в нашей повседневной пище, особенно растительной.
Микробные геномы, таким образом, служат важным дополнением к геному Homo sapiens. Хоть это и нехарактерно для публикуемых в наши дни на Западе научных работ, авторы в данном случае решились на философское обобщение. По их мнению, человека следует рассматривать как «сверхорганизм», чей обмен веществ обеспечивается совместной слаженной работой ферментов, закодированных не только в геноме Homo sapiens, но и в геномах сотен видов симбиотических микробов. Между прочим, доля человеческих генов в совокупном геноме этого «сверхорганизма» составляет не более 1%.
(Источник: Steven R. Gill et al. Metagenomic Analysis of the Human Distal Gut Microbiome // Science. 2006. V. 312. P. 1355–1359.)
—————
На симбиозе были основаны многие важнейшие ароморфозы (прогрессивные эволюционные преобразования). Мы уже познакомились с самым значительным из них — формированием эукариотической клетки, той основы, из которой в дальнейшем развились все высшие формы жизни (животные, растения, грибы и протисты[37]). В дальнейшем роль симбиоза в развитии жизни отнюдь не снижалась. Важнейшие функциональные блоки современной биосферы целиком и полностью держатся на симбиозе и симбиотических комплексах — «сверхорганизмах». Так, возможности высших растений — основных производителей органики и кислорода — были бы весьма ограничены без симбиоза с бактериями, способными переводить атмосферный азот в доступную для растений форму, и с почвенными грибами (микориза), без кооперации с насекомыми-опылителями и позвоночными — распространителями семян. Растительноядные животные — основные потребители производимой растениями органики — не могут эффективно переваривать растительную пищу без помощи разнообразных симбиотических бактерий и одноклеточных эукариот. Самые яркие и богатые жизнью морские экосистемы коралловых рифов невозможны без симбиоза коралловых полипов с одноклеточными водорослями — зооксантеллами. Сообщества различных экзотических, архаичных и экстремальных местообитаний (таких как наземные и подводные горячие источники, выходы метана и сероводорода, соленые лагуны, подземные воды и др.) тоже сплошь и рядом представляют собой сложные симбиотические комплексы микроорганизмов, в которых порой принимают участие и высшие организмы.
Одними из самых важных являются так называемые азотфиксирующие симбиозы — кооперация растений с микроорганизмами, способными переводить азот из атмосферы или захороненной в почве органики в доступную для растений форму (аммоний, NH4+). Основная часть биосферного азота содержится в атмосфере в химически инертной молекулярной форме (N2). Восстановление (фиксация) этого азота требует огромного количества энергии. Как мы знаем из главы «Планета микробов», на это способны лишь некоторые бактерии и архей, у которых есть специальные ферменты — нитрогеназы. Дополнительная сложность состоит в том, что нитрогеназы работают только в анаэробных (бескислородных) условиях. Все высшие (эукариотические) организмы, в том числе растения по определению аэробны, и в этом, возможно, главная причина того, что у высших организмов способность к фиксации азота не встречается. Много азота содержится также в почве в составе органических веществ, но и этот азот для растений недоступен, поскольку у них нет пищеварительных ферментов, необходимых для расщепления этой органики.
Два компонента азотфиксирующего симбиоза — это наземное растение (здесь годится любой тип растений) и какие-нибудь бактерии, способные фиксировать азот. В роли последних могут выступать цианобактерии, актинобактерии и альфапротеобактерии. Наиболее изучен симбиоз бобовых с клубеньковыми бактериями — ризобиями. Ризобии относятся к группе альфапротеобактерий. Ризобии, живущие в специализированных органах (клубеньках), снабжают растение аммонием, взамен получая весь комплекс элементов питания, в первую очередь углеводы, образуемые в ходе фотосинтеза. Между растительным и бактериальным компонентами симбиотического комплекса сложилась эффективная и гибкая система взаимной координации и регуляции. Например, специальные ферменты растений, работающие только в клубеньках, «заботятся» о том, чтобы концентрация кислорода в центральной части клубенька, где живут ризобии, была как можно ниже (и она там действительно ниже, чем в атмосфере, на 5–6 порядков). Биохимическая и генетическая интеграция симбиотического комплекса доходит даже до того, что активность некоторых растительных генов регулируется бактериальными белками!
Важную экологическую роль играет также симбиоз растений с азотфиксирующими цианобактериями. В отличие от ризобий цианобактерии сами способны к фотосинтезу, что несколько упрощает задачу снабжения азотфиксирующих симбионтов необходимой энергией. Симбиотический комплекс водного папоротника Azolla и цианобактерии Anabaena имеет большое сельскохозяйственное значение: заселение рисовых плантаций этим папоротником резко повышает урожайность риса. Неслучайно в некоторых районах Юго-Восточной Азии азоллу обожествляют.
В народнохозяйственных целях было бы очень полезно «научить» сами растительные клетки фиксировать атмосферный азот, точнее, их органеллы — пластиды. Теоретически это возможно, ведь многие «дикие» цианобактерии умеют фиксировать азот (мы ведь не забыли, что пластиды — потомки цианобактерий). Наверное, можно генно-инженерными методами создать пластиды с генами нитрогеназ, которые могли бы работать в темноте (например в корнях). Конечно, будет очень сложно добиться достаточно низкой концентрации кислорода в растительных клетках, но перспектива выглядит весьма заманчивой, ведь недостаток доступного азота — главный лимитирующий фактор, ограничивающий рост растений. Сняв это ограничение, можно было бы добиться колоссального увеличения урожайности.
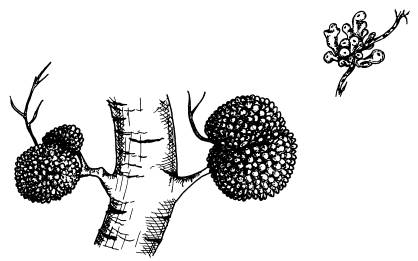
Клубеньки с азотфиксирующими бактериями встречаются не только у бобовых, но и у некоторых других растений. На рисунке — клубеньки на корнях ольхи.
Огромную роль в биосфере играют также симбиозы автотрофов с гетеротрофами — кооперация организмов, синтезирующих органику из углекислого газа, с потребителями готовой органики. В роли первых выступают фотосинтезирующие организмы (растения, одноклеточные эукариоты, цианобактерии) или бактерии-хемоавтотрофы, использующие для фиксации CO2 энергию окисления неорганических веществ (например, сероводорода или метана). В роли вторых выступают животные или грибы. Широко распространены симбиозы с участием грибов — микоризы и лишайники. В случае микоризы грибной компонент получает от растения-хозяина углеводы (глюкозу, фруктозу), а сам берет на себя функцию корневых волосков (которые на микоризных корнях часто не развиваются) и вдобавок снабжает хозяина азотом и фосфором, которые гриб добывает, разлагая почвенную органику. Лишайники иногда называют «микоризой наоборот», поскольку в этих симбиотических комплексах гриб выступает в роли хозяина, а фотоситезирующие организмы (одноклеточные водоросли или цианобактерии) — в роли симбионта. Однако система биохимической интеграции у лишайников и микориз во многом сходна. Наибольшего совершенства эта система достигает у трехкомпонентных лишайников, в состав которых входят помимо гриба-хозяина специализирующиеся на фотосинтезе зеленые водоросли и специализирующиеся на азотфиксации цианобактерии.
—————
Выход растений на сушу — результат симбиоза. Первые наземные растения появились в конце силурийского периода (более 400 млн лет назад). Недавно палеонтологи обнаружили, что уже самые первые наземные растения жили в симбиозе с грибами: у них была самая настоящая микориза. У этих растений еще не было настоящих корней — вместо них имелись так называемые ризоиды, не способные самостоятельно всасывать что-либо из почвы и служившие только для закрепления в грунте, а также, как теперь выяснилось, для обеспечения симбиоза с почвенными грибами. По-видимому, без этого симбиоза растения вообще не смогли бы выйти на сушу. Симбиоз растений с азотфиксирующими бактериями возник позже, причем для этого растениям оказалось достаточно лишь немного изменить те генетические системы, которые сложились у них ранее для взаимодействия с микоризными грибами.
—————
Симбиоз с автотрофами открывает большие возможности для многих водных животных, особенно малоподвижных (кишечнополостных, губок, асцидий, некоторых червей и моллюсков). Такие симбиотические комплексы представляют собой «сверхорганизмы», сочетающие признаки растений и животных (яркий пример — коралловые полипы). Автотрофы не только снабжают хозяина органикой, полученной в результате фото- или хемосинтеза, но и в ряде случаев помогают ему избавляться от конечных продуктов азотного обмена (например, мочевой кислоты или мочевины), которые служат для симбионтов ценным источником азота.
Чрезвычайно широко распространены также симбиозы животных с микробами, помогающими усваивать растительную пищу. Потребление органики, производимой растениями в ходе фотосинтеза, — главная «экологическая роль» животных в биосфере, однако, как это ни парадоксально, сами по себе животные практически не способны справляться с этой задачей. Подавляющее большинство растительноядных животных попросту лишены ферментов для расщепления растительных полимеров (главным из которых является целлюлоза). Поэтому практически все животные-фитофаги — это на самом деле симбиотические комплексы из животного-хозяина и разнообразных бактерий, грибов или простейших (причем в последнем случае симбиотические простейшие зачастую сами имеют бактериальных симбионтов). Скорее всего, растительноядность изначально была симбиотическим феноменом. Роль симбионтов не сводится к расщеплению растительных полимеров: они могут также утилизировать азотные шлаки хозяина и синтезировать многие вещества, необходимые хозяину, но отсутствующие в растительной пище. Микробное сообщество, обитающее в пищеварительном тракте термитов, обладает даже способностью к азотфиксации, что позволяет этим насекомым питаться такими несъедобными вещами, как химически чистая целлюлоза. Некоторые биохимические процессы в таких симбиотических системах оказываются весьма причудливым образом распределены между хозяином и симбионтом. Например, комплекс «тли — бактерия Buchnera» синтезирует важнейшее вещество кофермент А совместными усилиями: сначала бактерия синтезирует из пирувата пантотеновую кислоту (чего не может насекомое), а затем тля синтезирует из пантотеновой кислоты кофермент А (чего не может бактерия). Конечным продуктом пользуются они вместе.
—————
Способность тлей приспосабливаться к колебаниям температуры зависит от симбиотических бактерий. Тли питаются исключительно соками растений. Жить на этой скудной диете им позволяет удачный симбиоз с бактериями Buchnera. Симбионты получают от хозяев кров и пропитание, а в обмен синтезируют для них аминокислоты, витамины и другие вещества, напрочь отсутствующие в той чуть сладенькой водичке, которая составляет единственную пищу тлей.
То, что тли представляют собой в действительности симбиотические «сверхорганизмы», придает довольно своеобразный характер их экологии и эволюции. Например, у тлей вполне возможно наследование благоприобретенных признаков «по Ламарку». Ведь за жизнь одной тли в ее клетках сменяется несколько поколений симбионтов-бухнер, при этом генофонд бактерий может измениться в результате мутаций и отбора, и тогда тля передаст своим потомкам симбионтов, генетически отличных от тех, что она сама получила от матери (см. главу «Наследуются ли приобретенные признаки?»).
Недавно сотрудники Факультета экологии и эволюционной биологии Аризонского университета в Тусоне (США) обнаружили, что генетические особенности симбионтов могут сильнейшим образом сказываться на жизнеспособности и экологических характеристиках всего симбиотического комплекса.
Симбиозу тлей с бухнерами уже более 100 миллионов лет. За это время бухнера подверглась генетической деградации — это почти неизбежная судьба всех внутриклеточных симбионтов (конечным этапом деградации, как мы знаем, может стать превращение бактерии в клеточную органеллу). Бухнера потеряла способность к рекомбинации (обмену генами с другими бактериями), ее геном сильно сократился, упростился и потерял пластичность из-за утраты мобильных генетических элементов и повторов. Это снизило приспособительные способности бактерии. Как любой организм, привыкший к относительно неизменным благоприятным условиям, бухнера стала ужасно изнеженным и капризным существом. А это, в свою очередь, накладывает ограничения на весь «сверхорганизм». Например, бухнеры не любят жары — и это сильно мешает тлям распространяться в тех регионах, где высока вероятность перегрева. Между прочим, ограничения такого рода характерны и для многих других сверхорганизмов: мелкие наземные позвоночные не могли как следует освоить растительную диету, пока не выработали теплокровность (необходимая для переваривания клетчатки кишечная флора не выносит охлаждения); вспомним также, какие усилия прикладывают для поддержания постоянной температуры в своих жилищах термиты.

Компоненты симбиотического сверхорганизма: тля (справа) и бактерии Buchnera, живущие в специализированных клетках хозяина — бактериоцитах.
Главная проблема, с которой организм сталкивается при перегреве, — это возможность денатурации (необратимого изменения структуры) некоторых белков. Обычно клетка в ответ на повышение температуры включает ряд генов (их называют генами теплового шока), кодирующих специальные белки — шапероны, чья функция состоит втом, что они «насильно» придают другим белкам правильную конфигурацию (см. сюжет «Когда вредных мутаций много, они не так вредны» в главе «Управляемые мутации»). Гены теплового шока (в количестве пяти штук) есть и у бухнеры.
Любопытно, что некоторые гены, которые у других организмов обычно активируются при перегреве, у бухнеры всегда работают с повышенной активностью. Дело в том, что неправильное сворачивание белков может происходить не только из-за высоких температур, но и из-за вредных мутаций в генах, кодирующих эти белки. Бухнера, лишенная способности к рекомбинации, обречена на постоянное накопление мутационного груза, а шапероны до некоторой степени могут сгладить негативные эффекты мутаций, сворачивая правильным образом не совсем правильные белки.
Ученые взяли три лабораторные линии гороховой тли Acyrthosiphon pisum и измерили активность генов бухнеры[38] при нормальной температуре и при перегреве. Оказалось, что у бухнер одной из трех линий при тепловом шоке не активируется ген ibpA, который кодирует небольшой «теплозащитный» белок, имеющийся не только у бактерий, но и практически у всех живых организмов.
Поскольку линия тлей, где обнаружилась эта аномалия, была произведена всего пять лет назад от «нормальной» линии, в которой ibpA активируется при перегреве, ученые предположили, что причиной является мутация, возникшая менее пяти лет назад.
Геном бухнеры прочтен, и найти мутацию было нетрудно. Исследователи отсеквенировали (определили нуклеотидную последовательность) регуляторной области гена ibpA и обнаружили, что у бактерий с аномальной реакцией на перегрев «потерялся» один нуклеотид (аденин) в том месте, где к хромосоме присоединяется регуляторный белок y32, который как раз и включает ген ibpA (и другие гены теплового шока) в стрессовой ситуации.
В дальнейшем были проверены многие другие лабораторные линии тлей этого вида, и в одной из них обнаружился второй случай независимого возникновения той же самой мутации. Зная историю изучаемых линий, авторы установили, что один раз мутация произошла в 2001 году, второй — в 2005-м. Естественно, ученые захотели выяснить, встречается ли эта мутация в природных популяциях. Оказалось, что она довольно обычна у тлей из прохладных штатов Нью-Йорк и Висконсин (в некоторых популяциях мутантные симбионты присутствуют у 20% тлей), но не встречается в штатах Аризона и Юта, где бывает по-настоящему жарко.
Эксперименты подтвердили, что у всех мутантов ген ibpA не активируется при перегреве, а у всех линий без мутации — активируется. Остальные гены теплового шока активировались совершенно одинаково как у мутантных, так и у нормальных бухнер.
Исследователи решили выяснить, каким образом мутация, произошедшая у бактериального симбионта, отражается на жизнеспособности и репродуктивном потенциале симбиотического сверхорганизма. Тлей с мутантными и нормальными бухнерами выращивали в разных температурных условиях: при постоянной температуре 15 и 20 °C, а часть насекомых в возрасте двух дней после вылупления из яйца подвергали четырехчасовому нагреванию до 35,5 °C. Выяснилось, что после перегрева тли с мутантными симбионтами почти полностью утрачивают способность к размножению, тогда как тли с нормальными симбионтами размножаются вполне успешно. Однако при постоянной температуре 15 или 20 °C заметным преимуществом обладают тли с мутантными симбионтами. Они раньше начинают размножаться и в среднем производят больше потомков.
Дополнительные эксперименты позволили установить, почему кратковременный перегрев так губительно сказывается на тлях с мутантными бухнерами: повышение температуры до 35 °C или выше приводит к массовой гибели бактериальных симбионтов. После этого остается всего лишь около 1000 бактерий на одно насекомое, тогда как норма составляет примерно миллион.
Таким образом, мутантные бухнеры дают преимущество тлям при постоянной невысокой температуре среды, а «нормальные» выгодны в том случае, если существует опасность перегрева. По-видимому, в зависимости от климатических условий местности и даже от времени года направление отбора в природных популяциях тлей может меняться: иногда преимущество получают тли с обычными, а иногда — с мутантными симбионтами. В результате оба генетических варианта сохраняются в природных популяциях, и ни один из них не может полностью вытеснить другой.
(Источник: H. E. Dunbar, A. C. C. Wilson, N. R. Ferguson, N. A. Moran. Aphid Thermal Tolerance Is Governed by a Point Mutation in Bacterial Symbionts // PLoS Biol. 2007. 5(5): e96.)
—————
Совместное существование позволяет каждому из компонентов симбиотического комплекса отказаться от тех биохимических функций, которые лучше удаются партнеру, и сосредоточиться на том, что лучше получается у него самого. Например, клубеньковые бактерии занимаются почти исключительно фиксацией азота, переложив заботу обо всем остальном на растение-хозяина. Часто один из партнеров специализируется на поставке в систему азота, а другой — углерода. По мере развития симбиотических отношений симбионт может полностью утратить самостоятельность и превратиться в неотъемлемую часть своего хозяина (впрочем, надо признать, что и хозяин при этом теряет самостоятельность и перестает быть «автономным организмом»).
Грань между мутуалистическими (взаимовыгодными) и антагонистическими симбиозами — довольно условная и зыбкая. Например, многие растительно-грибные симбиозы в ходе эволюции могли долго «балансировать» на грани мутуализма и антагонизма, причем преобладающие потоки питательных веществ могли неоднократно менять свое направление. Непосредственный переход паразитизма в мутуализм — сранительно редкое явление (пример — «защитные симбиозы» растений со спорыньевыми грибами, в которых исходно паразитический гриб стал защищать растение от растительноядных животных путем синтеза токсичных веществ). В большинстве случаев симбиотические системы развиваются из фрагментов сообществ (экосистем). Например, симбиозы, основанные на биохимической кооперации, в большинстве случаев развиваются из кооперативных объединений свободноживущих организмов, соместно утилизирующих какой-то ресурс, или из фрагментов «трофической пирамиды» (симбиозы производителей органики с ее потребителями). Особый и весьма удивительный случай связи между трофической цепью и биохимическим симбиозом представляет морской моллюск Elysia viridis, питающийся водорослями. Этот моллюск ухитряется переселять пластиды съеденных водорослей в свои собственные клетки и долгое время сохранять их там живыми, приобретая таким образом способность к фотосинтезу. Настоящий гибрид животного и растения![39]
В трех заключительных разделах этой главы я расскажу о трех в высшей степени необычных симбиотических системах, которые ученым удалось недавно «расшифровать».
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Симбиоз насекомых и микробов
Симбиоз насекомых и микробов В теле насекомых, как правило, находятся бактерии, дрожжевые и другие микроскопические грибы. Они живут в клетках особого органа, получившего название мицетом. Там они синтезируют витамины, а возможно, и другие жизненно важные вещества,
Глава 3. Великий симбиоз
Глава 3. Великий симбиоз Эволюция «с человеческим лицом» Типичным примером примитивизации дарвиновских идей является широко распространенное по сей день мнение о том, что в основе эволюции лежат исключительно эгоизм, конкуренция и безжалостная «борьба за
Горизонтальный перенос генов и симбиоз
Горизонтальный перенос генов и симбиоз Дупликация и перестановки участков ДНК одного и того же генома открывают большие эволюционные возможности. Еще больше их дает комбинирование фрагментов разных, неродственных геномов[76]. Обмен генами между неродственными
Симбиоз на службе у гигиены
Симбиоз на службе у гигиены В зверином царстве взаимные услуги — дело обычное. Если вы думаете, что животные только охотятся друг за другом и сильные пожирают слабых в бесконечной и жестокой борьбе за существование, то вы глубоко заблуждаетесь. Борьба борьбой — она,
Часть первая Симбиоз, или содружество соседей
Часть первая Симбиоз, или содружество соседей
Симбиоз на земле и под водой
Симбиоз на земле и под водой Открытие симбиозаГеродот приехал в Египет в 450 году до нашей эры. На Меридовом озере он видел шлюзы, которые регулировали уровень великой реки. Гулял по аллее сфинксов, спускался в усыпальницу священных быков, блуждал по Лабиринту.Потом
Лучезарный симбиоз
Лучезарный симбиоз Открытие ОзориоХуанито разрубил акулу на куски. Потом каждый кусок стал резать на мелкие дольки.Старик брал эти дольки и тер их о живот черной рыбы: три раза вправо, три влево. Потом переворачивал кусок и проводил другой его стороной по жесткой чешуе.
Симбиоз в цветке
Симбиоз в цветке Булки только для муравьевСимбиоз животных с растениями не ограничивается лишь союзом грибов и насекомых. Многие высшие растения — травы и деревья — путем естественного отбора приобрели особые приспособления, предназначенные специально для
СИМБИОЗ
СИМБИОЗ Рифтия (Riftia), трубчатый червь длиной около метра, является важным членом сообщества гидротермальных источников, расположенных на срединно-океанических хребтах, в тех местах, где дно океана покрыто трещинами. У рифтии нет ни рта, ни пищеварительной системы. Как же