Порядок из хаоса
Порядок из хаоса
Излюбленным аргументом креационистов является утверждение о невозможности самопроизвольного развития сложных структур из простых на основе случайных изменений. Действительно, здравый смысл вроде бы подсказывает, что, сколько бы мы ни ворошили лопатой кучу мусора, ничего полезного и принципиально нового из обрывков и обломков само собой не соберется. Креационисты очень любят приводить эту аналогию (она известна во многих вариантах). Часто в этом контексте поминают также второе начало термодинамики, согласно которому якобы невозможен самопроизвольный рост упорядоченности — сам собой нарастать может только хаос.
Все это, сказать по правде, полная ерунда. Подобные рассуждения изобилуют логическими ошибками: аналогии не соответствуют объясняемому феномену, а физические законы привлекаются не к месту, то есть делается попытка применить их за пределами установленной для них области применимости. Например, второе начало термодинамики справедливо только для изолированных систем, которые не получают энергии из внешней среды. Живые организмы, напротив, всегда получают энергию извне и, расходуя часть этой энергии на повышение упорядоченности внутри себя, создают при этом порядочный хаос снаружи. Суммарная неупорядоченость (энтропия) всей системы (организм + среда) при этом растет. Поэтому никакого противоречия между развитием жизни и вторым началом термодинамики попросту не существует. Возможность самоорганизации — самопроизвольного рождения порядка из хаоса — показана экспериментально и обоснована теоретически для самых разных типов открытых неравновесных систем.
Кроме того, говоря о «случайности», якобы лежащей в основе предполагаемых механизмов эволюции, креационисты занимаются откровенным передергиванием. Они спекулируют на многозначности и расплывчатости термина «случайность». В действительности эволюция основана не на случайностях, а на вполне строгих закономерностях (о которых мы еще будем говорить). Даже мутации, которые до недавнего времени действительно было принято считать случайными, на самом деле далеко не всегда таковы (см. главу «Управляемые мутации»). Дарвиновский механизм естественного отбора сам по себе вполне достаточен для того, чтобы придать эволюционному процессу упорядоченность. Пусть даже первичные изменения (мутации) происходят случайно — благодаря действию отбора запоминание системой произошедших изменений происходит уже не случайно, а строго закономерно. Это избирательное запоминание и производит новую информацию и новую сложность. Между прочим, математикам хорошо известна так называемая S-теорема Ю. Л. Климонтовича, представляющая собой строгое математическое доказательство того, что новая информация (или «отрицательная энтропия», упорядоченность) порождается, сочетанием случайного изменения состояния системы с последующим необходимым (избирательным) запоминанием результатов изменения. Иными словами, эволюционный механизм, предложенный Дарвином, является совершенно адекватным и достаточным объяснением самопроизвольного усложнения живых систем в ходе эволюции — как с точки зрения биологии, так и с точки зрения физики и математики[44].
Чтобы убедиться своими глазами в возможности самозарождения сложного из простого, налейте в ванну воды и выньте пробку слива (тем самым превратив закрытую систему в открытую). Вскоре хаотическое движение молекул воды под воздействием силы тяжести на ваших глазах породит довольно сложную, упорядоченную структуру — воронку-водоворот. Когда видишь это впервые в жизни (я помню свое впечатление из раннего детства), возникает ощущение чуда. Почему? Да просто наше мышление так устроено: оно специализировано для целеполагания, мы привыкли планировать свои действия, ориентируясь на ожидаемый результат. Поэтому нам и кажется, что если чего-то не предусмотришь, не проконтролируешь, то ничего хорошего и не выйдет — только разруха и хаос. Раз за разом нас подводит эта «инструментальная» направленность нашего мыслительного процесса. Так, в высокоразвитых сельскохозяйственных цивилизациях Мексиканского нагорья царило убеждение, что если пустить дело на самотек, то ни солнце утром не взойдет, ни лето в положенный срок не настанет, а потому необходимо регулярно подкармливать жертвенной кровью тех светлых богов, что в непрестанной героической борьбе с силами хаоса поддерживают привычный нам миропорядок. Ныне те боги, хочется верить, уже умерли с голоду, но на смене дня и ночи это, как видим, не отразилось.
Аналогии с мусорной кучей и лопатой, или с землетрясением, которое может только разрушить здание, но никогда не соберет его из камней, или с мартышкой, которая, случайно нажимая на клавиши, никогда не напишет «Войну и мир», имеют примерно такое же отношение к эволюции, как ритуальная практика ацтеков — к вращению Земли вокруг солнца. Те эволюционные процессы, в результате которых рождается новая сложность, очень мало похожи на эти воображаемые ситуации. Если какое-то сходство и есть, то очень неполное, ограниченное отдельными признаками, поверхностное и ничего не говорящее о сути дела.
Мы уже упомянули вскользь об упорядочивающей и направляющей роли отбора и о неслучайности многих мутаций (наследственных изменений). Кроме того, «обрывки и обломки», из которых в процессе эволюции собирается новое, совсем не похожи на те разнородные предметы, которые можно обнаружить в мусорной куче.
Начать с того, что материал наследственности (ДНК), генетический код и базовая «технология» его прочтения и реализации у всех организмов одни и те же. Этот факт, между прочим, рассматривается учеными как решающее доказательство происхождения всего живого на нашей планете от единого общего предка (будь то предковый вид или предковое сообщество).
Жизненные задачи тоже у всех организмов сходны (выжить, добыть нужные вещества и энергию, вырасти, противостоять внешним разрушающим воздействиям, оставить потомство). Поэтому все имеющиеся в наличии на нашей планете фрагменты ДНК, во-первых, понятны для всего живого (могут быть более-менее адекватно прочтены и «интерпретированы» любой живой клеткой[45]), во-вторых, все они являются фрагментами устройств, сходных по своим функциям. Это резко повышает вероятность того, что случайное комбинирование фрагментов (например, в ходе горизонтального обмена генами, о котором рассказано в главе «Наследуются ли приобретенные признаки?») может породить что-то новое и осмысленное.
Была раньше такая хорошая игрушка — радиоконструктор (что-то вроде нынешнего «Лего», только из конденсаторов, ламп, катушек и прочих радиодеталей). Простые работающие устройства — такие, например, как детекторный приемник — из этого конструктора собирались с достаточно высокой вероятностью путем абсолютно случайного, произвольного соединения деталек. Этот пример показывает, что если «детальки» хороши, то даже случайное их комбинирование вполне может породить что-то полезное и жизнеспособное. А в «эволюционном конструкторе» детальки, несомненно, самого высшего сорта — отшлифованные естественным отбором, проверенные и испытанные многими поколениями живых «испытателей».
Кроме того, геном организма или отдельный ген — это далеко не «Война и мир», а нечто гораздо более помехоустойчивое. Если мы заменим случайным образом букву, слово или предложение в талантливом романе, то наверняка хоть чуть-чуть, но испортим произведение. Если мы заменим в гене нуклеотид или в белке аминокислоту, с очень большой вероятностью не произойдет абсолютно ничего плохого (более того, есть даже вполне реальный шанс, что изменение окажется полезным, но об этом чуть позже).
Белковая молекула обычно состоит из нескольких сотен аминокислот, но только очень немногие из них действительно необходимы для того, чтобы белок исправно выполнял свою функцию. Если речь идет о ферменте, то для его работы абсолютно необходимы в основном те аминокислоты, которые составляют так называемый «активный центр». Активный центр — это то место белковой молекулы, которое, собственно, и катализирует реакцию. Кроме того, могут оказаться важными и некоторые аминокислоты, определяющие пространственную конфигурацию молекулы — то, в какую фигуру она самопроизвольно «свернется» после того, как будет синтезирована. Например, в построении пространственной структуры фермента участвуют цистеины — это аминокислоты, содержащие серу, которые образуют дисульфидные мостики (-S-S-), скрепляющие между собой различные витки, лопасти и спирали белковой молекулы.
Такие принципиально важные аминокислоты составляют лишь малую часть белка. Более того, это даже не конкретные наборы аминокислот, которые должны находиться в строго определенных местах белковой молекулы, а довольно расплывчатые «рисунки», «паттерны», или, как их официально называют, мотивы.
—————
Белки с одинаковой функцией могут сильно различаться по структуре. Вот аминокислотная последовательность фермента фруктозо-бисфосфат альдолазы жгутиконосца Euglena gracilis. Это один из ферментов гликолиза — важного биохимического процесса, в ходе которого, как мы помним, клетка тратит часть энергии, заключенной в молекуле глюкозы, для синтеза двух молекул АТФ без использования кислорода. Каждая буква соответствует определенной аминокислоте (P — пролин, D — аспарагиновая кислота, F — фенилаланин, K — лизин и т. д.):
PDFPKDLKGV LDGNQVRTLF DFAQKKGFAI PAVNCTSSST VNVVLERARD THNPVIIQVS QGGAAFYCGK GVKDEKLIAS VDGSVALAHH VRAVAHTMAP VVVHSDHCAK KLLPWFDGML DADGEIFCEH GVPLFSSHML DLSEENDEED IGTCVKYFTR MAKLNLWLEM EIGMTGGVED GVDNSGVAND KLYTSSEQVF AVHKALGASS PNFSIAAAFG NVHGVYKPGN VKLQPNLLKE HQDYARKQLS SSEDHPLYLW FHGPSGSTDA EIHEAVRNGV VKMNLDTDMQ WAYWDGLRQF EAKKHDYLQG QIGNPEGPDK PNKNYYDPRK WIREAELGML ARVKVAFKAV ELPGGLKEFI GIP[46].
А вот тот же самый фермент бактерии Mycoplasma pneumoniae:
MLVNIKQMLQ HAKQHHYAVP HININNYEWA KAVLTAAQQA KSPIIVSTSE GALKYISGHQ VVVPMVKGLV DALKITVPVA LHLDHGSYEG CKAALQAGFS SIMFDGSHLP FQENFTKSKE LIELAKQTNA SVELEVGTLG GEEDGIVGQG ELANIEECKQ IATLKPDALA AGIGNIHGLY PDNWKGLNYE LIEAIAKATN LPLVLHGGSG IPEADVKKAI GLGISKLNIN TECQLAFAKA IREYVEAKKD LDTHNKGYDP RKLLKSPTQA IVDCCLEKMQ LCGSTNKA[47].
На первый взгляд заметить сходство между этими белками очень непросто. На второй, впрочем, тоже. А ведь это, по молекулярно-генетическим меркам, весьма похожие друг на друга, близкородственные белки! У них есть длинный сходный участок (от 16-й до 333-й аминокислоты первой молекулы и от 5-й до 265-й аминокислоты второй), в пределах которого совпадает целых 25% аминокислот. Кроме того, некоторые другие аминокислоты в двух белках хотя и разные, но близкие по своим химическим свойствам.
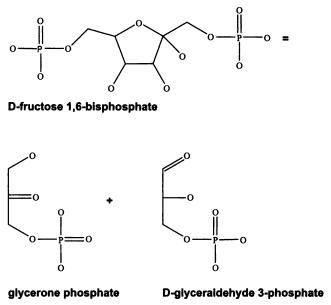
Реакция, которую катализирует фермент фруктозо-бисфосфат альдолазы.
Для того чтобы обнаружить это сходство и оценить его количественно, можно воспользоваться специальной программой BLAST, свободно доступной в Интернете по адресу http://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi. Taм надо просто вставить две аминокислотные последовательности в два окошка и нажать кнопку.
Такой уровень сходства при сравнении белков представителей разных надцарств живой природы (мы сравнивали эукариот с бактериями) считается очень высоким. В данном случае, возможно, этот результат объясняется тем, что предки эвглены сравнительно недавно приобрели соответствующий ген путем горизонтального переноса от каких-то бактерий. У высших эукариот фруктозо-бисфосфат альдолазы этого семейства вообще не встречаются. У нас с вами, например, они совсем другие.
Читатель может попробовать самостоятельно сравнить между собой фруктозо-бисфосфат альдолазы различных организмов (найти их можно, например, в базе данных PFAM http://pfam.sanger.ac.uk/, осуществив поиск по ключевым словам «fructose bisphosphate aldolase»). Любой желающий может легко убедиться, что если сравнить один и тот же (по функции) фермент, взятый у человека и, допустим, у кишечной палочки Escherichia coli, то программа сравнения, скорее всего, выдаст безрадостное «no significant similarity found» — «значимого сходства не обнаружено». Это, впрочем, не значит, что никакого сходства действительно нет — оно есть (это те самые неопределенные «мотивы», о которых шла речь выше), но для его выявления необходимы более мощные аналитические средства, чем программа BLAST.
Тем не менее эти столь разные белки примерно с одинаковой эффективностью выполняют одну и ту же работу, то есть катализируют одну и ту же биохимическую реакцию.
—————
Конечно, не все различия между разными белками, выполняющими одну и ту же функцию, случайны и бессмысленны. Некоторые из них обеспечивают тонкую подстройку фермента под конкретные особенности той или иной клетки. Однако, как показывает генно-инженерная практика, довольно часто эти белки оказываются вполне взаимозаменимыми. Это значит, что если заменить у какого-либо организма его собственный ген на чужой, кодирующий фермент, тот же по функции, но иной по структуре, то такой генно-модифицированный организм с большой вероятностью окажется вполне жизнеспособным.
Из этого видно, что «подобрать» белок с нужной функцией методом случайного перебора аминокислот в принципе не так уж сложно. Для этого вовсе не требуется правильно угадать все несколько сотен аминокислот, составляющих молекулу белка. Достаточно подобрать лишь некий довольно расплывчатый «мотив».
Эксперименты показывают, что такая технология отлично работает. Среди коротких белковых молекул, искусственно синтезированных путем случайного комбинирования аминокислот, удается отыскать белки практически с любой каталитической активностью. Конечно, эффективность таких катализаторов ниже, чем у настоящих ферментов, но это неважно. Главное, чтобы отбору было за что «зацепиться», и он со временем доведет функцию до совершенства. Эта технология годится не только для белков, но и для функциональных РНК. Именно таким способом исследователи ищут и находят новые рибозимы — РНК с ферментативными свойствами (см. главу «Происхождение жизни»).
Отобрав подходящие молекулы, можно затем раз за разом вносить в них небольшие случайные изменения и отбирать лучшие из получившихся вариантов. Это называют «методом искусственной эволюции». Само его существование и широкое практическое применение является отличным доказательством работоспособности дарвиновской модели.
Однако изготовить высокоэффективный белковый фермент с изначально заданными свойствами методом искусственной эволюции — дело довольно долгое и трудоемкое. Поэтому ученые в последние годы прикладывают большие усилия, чтобы не только выявить, но и понять связи между последовательностью аминокислот и функцией белка. Если эта цель будет достигнута, ферменты с заданными свойствами можно будет проектировать, то есть просто вычислять на компьютере искомую последовательность аминокислот. Это сэкономит огромные средства и позволит осуществить настоящую революцию в химической промышленности. Ведь ферменты — чрезвычайно эффективные катализаторы, способные в тысячи раз ускорить синтез чуть ли не любых химических соединений.
Вообще белки — удивительные вещества, заключающие в себе немало парадоксов. Как мы уже знаем, многие аминокислоты, входящие в состав белковой молекулы, могут быть заменены или удалены без заметного изменения свойств белка. С другой стороны, замена всего одной-двух аминокислот в активном центре белковой молекулы может привести к радикальному изменению свойств фермента. В большинстве случаев, конечно, фермент от этого портится, снижает производительность или вовсе становится нефункциональным. В других случаях это может привести к модификации или даже полной смене функции, то есть к появлению нового фермента.
—————
Разнообразие белков началось с немногих исходных «мотивов». Некоторые ученые предполагают, что на самых ранних этапах становления жизни — возможно, еще в РНК-мире — методом «слепого поиска» было найдено несколько сотен базовых аминокислотных мотивов, соответствующих основным функциям белков. Изначально эти функции были реализованы на основе очень коротких белковых молекул (пептидов), причем эффективность функционирования была поначалу низкой. Но все равно это был большой шаг вперед по сравнению с неуклюжими рибозимами. Эти первичные «мотивы», оформленные в виде коротких пептидов, в дальнейшем легли в основу всех ныне существующих надсемейств белков.
—————
Конечно, разумное просчитанное проектирование может значительно ускорить создание новых белков, на которое эволюции требуются миллионы лет. Иногда, впрочем, белки с новыми функциями возникают в природных популяциях живых организмов гораздо быстрее — буквально за считанные годы или даже месяцы.
Здесь нужно различать две принципиально разные ситуации. Иногда новые белки возникают благодаря особым механизмам, специально для этого существующим в клетке. Таковы, например, образование новых антител в ответ на проникновение в организм новой инфекции или целенаправленное изменение поверхностных белков у некоторых патогенных бактерий, имеющее целью обмануть иммунную систему хозяина. Эти особые механизмы направленного изменения генома сами являются результатом длительной эволюции и будут рассмотрены нами отдельно в главе «Управляемые мутации». Но новые белки иногда могут быстро возникать и по обычной, классической схеме — за счет случайных мутаций и отбора. Самый известный пример такого рода — поразительно быстрая выработка бактериями устойчивости к антибиотикам. Как выяснилось, бактерии предпочитают не пассивную, а активную защиту: они не пытаются «обойти» или видоизменить тот биохимический процесс, по которому «бьет» новый антибиотик, а вырабатывают новый фермент, способный активно атаковать и обезвреживать лекарственное средство.
Хотя возникновение подобного фермента требует не одной мутации, а нескольких, классическая эволюционная модель вполне способна объяснить это явление. В слаженном появлении целого комплекса необходимых мутаций нет ничего противоречащего простейшей схеме «единичные случайные мутации плюс отбор». Кроме того, на примере развития устойчивости к антибиотикам можно понять, как работает один из механизмов, ограничивающих число допустимых эволюционных траекторий движения к заданной цели. Одним из следствий подобных ограничений могут быть параллелизмы — независимое возникновение сходных признаков в разных эволюционных линиях.
—————
Параллелизмом, или параллельной эволюцией, называют независимое появление похожих признаков у разных организмов. Параллелизмы очень широко распространены в природе, и биологи-теоретики уделяют много внимания этому явлению. В нем отчетливо проявляется закономерный (неслучайный) характер эволюции. Одной из основных причин параллелизмов является ограниченность возможных путей эволюционного преобразования организма. Сформулированный Н. И. Вавиловым «Закон гомологических рядов в наследственной изменчивости» (http://evoibiol.ru/vavilov.htm) позволил связать это явление с закономерностями внутривидовой изменчивости. У близких видов встречаются одинаковые вариации — например, у большинства злаков есть остистые и безостые формы. Палеонтология дает много ярких примеров схожести путей эволюции в разных группах (мы поговорим об этом подробно в главе «Направленность эволюции»).

Яркий пример параллельной эволюции: сумчатый саблезубый тигр Thylacosmilus из плиоцена Южной Америки (вверху) и «обычный» саблезубый тигр Smilodon из плейстоцена Северной Америки.
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 9. Общественный порядок без любви
Глава 9. Общественный порядок без любви … и в сердце вечный хлад Гёте Противопоставление анонимной стаи и личной связи, которым мы закончили предыдущую главу, означает лишь, что эти два механизма социального поведения в весьма значительной степени исключают друг друга;
У павианов главное — порядок
У павианов главное — порядок Многие виды стадных животных пришли к сходному решению проблемы полов: во избежание бесконечных стычек и драк самец изгоняет из стада ставших половозрелыми самцов, включая и собственных сыновей. Плодовитость стада от этого не страдает, так
Порядок
Порядок Закон, который я довольно расплывчато называю порядком или правильностью, очень важен в искусстве и дизайне, особенно в дизайне. Опять же, этот принцип так очевиден, что трудно говорить о нем так, чтобы это не звучало банально, но обсуждение зрительной эстетики
4. Навыки и порядок их отработки
4. Навыки и порядок их отработки Понятие о навыках. При дрессировке собаку приучают выполнять определенные действия. Каждое из действий, отрабатываемое у собаки при дрессировке, обозначается как навык. Навыки дрессировки весьма разнообразны и различны по своему
Порядок рождения и манипулирование родительским вкладом у человека
Порядок рождения и манипулирование родительским вкладом у человека В исследованиях в области эволюционной антропологии в последние годы большое внимание уделяется вопросу о роли раннего социального опыта в формировании особенностей поведения человека и его
Глава 7. Порядок и хаос в обществе.
Глава 7. Порядок и хаос в обществе. Самая главная и наиболее эффективная [сила], гарантирующая длительное сохранение… власти — это не насилие во всех его формах, используемое власти предержащими для управления подчиненными, а все виды уступок, на которые идут