Нужны ли эмбрионам гены?
Нужны ли эмбрионам гены?
Имеется еще одно странное обстоятельство, затрудняющее понимание природы индивидуального развития животных. Оно состоит в том, что ранние этапы онтогенеза, как правило, идут вообще без участия генов, при полностью отключенном геноме.
Задумаемся, что это значит. Ведь принято считать, что индивидуальное развитие — это постепенная «реализация» той генетической информации, которая заключена в геноме оплодотворенной яйцеклетки (зиготы) и которая в итоге окончательно «воплощается» в строении взрослого организма. Все понимают, что путь от генотипа к фенотипу труден и извилист, но лишь немногие эволюционисты всерьез пытаются объяснить ключевые закономерности эволюции особенностями тех сложнейших процессов самоорганизации, которые составляют суть онтогенеза (уж слишком трудна задача). Поэтому обычно проблему пытаются упростить, сведя все к вопросу о том, каким образом те или иные изменения генотипа (например, мутации) могут отразиться на процессе развития зародыша.
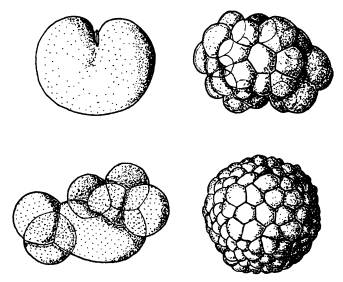
Делящаяся яйцеклетка (слева наверху), морула в виде комка клеток (слева внизу), бластула в виде полого шарика (справа внизу и вверху). На всех этих стадиях развития животных геном обычно не функционирует. На рисунке — развитие медузы Aequorea.
Геном традиционно рассматривается как активное начало (в нем все «закодировано», он «руководит» развитием). Развивающийся зародыш, напротив, считается чем-то вроде пассивного «результата» деятельности генома. Дело несколько осложняется тем, что сам геном в процессе онтогенеза явно не самодостаточен: в разных клетках эмбриона одни гены включаются, другие выключаются в строгой последовательности, определяемой, в частности, химическими сигналами, которыми обмениваются между собой клетки и ткани зародыша. Кто кем управляет, становится не совсем ясно. Некоторые теоретики по этому поводу даже заявляют, что геном — это не «программа развития зародыша», а скорее некий набор инструментов, которыми зародыш пользуется (или не пользуется) по своему усмотрению.
Для управления собственными генами зародыш использует множество разных механизмов: это и регуляция транскрипции (считывания генов) при помощи малых РНК и специальных регуляторных белков — транскрипционных факторов (Hox-гены, как мы помним, тоже кодируют транскрипционные факторы), и особые «эпигенетические» механизмы, о которых мы подробнее поговорим в главе 8.
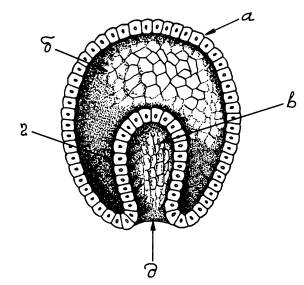
Типичная гаструла многоклеточного животного. а — эктодерма (наружный зародышевый листок), б — бластоцель (первичная полость тела), в — первичная кишка, г — энтодерма (внутренний зародышевый листок), д — бластопор (первичный рот).
Еще одно обстоятельство, которое делает сомнительным тезис о полной и однозначной обусловленности онтогенеза геномом зиготы, состоит в том, что, как уже было сказано выше, у подавляющего большинства животных на ранних стадиях развития геном вообще не функционирует. Он просто-напросто отключен, все гены молчат и матричные РНК («считываемые» с генов матрицы для синтеза белка) не производятся.
Зародыш между тем претерпевает сложные превращения. Яйцеклетка начинает дробиться, число клеток эмбриона растет в геометрической прогрессии: 2, 4, 8, 16, 32… Наконец формируется однослойный шар из клеток (бластула). Клетки, находящиеся на одном из полюсов бластулы, мигрируют внутрь, давая начало второму зародышевому листку (энтодерме), из которой позже разовьется кишечник. На этой стадии двуслойный зародыш называется гаструлой. Только на этом этапе у многих животных начинают наконец включаться гены, унаследованные от папы с мамой. У других это происходит чуть раньше — на стадии бластулы. И только млекопитающие — группа, уникальная во многих отношениях, — включают свои гены еще раньше (например, мышь делает это на стадии двух клеток)[64].
Как удается эмбриону развиваться без всякого «генетического контроля» вплоть до стадии гаструлы? Почему гены зародыша так долго остаются выключенными? Какие механизмы обеспечивают отключение генов в зиготе, а затем их своевременное включение[65]?
Ответ на первый вопрос более или менее ясен. Яйцеклетка содержит большое количество матричных РНК, унаследованных от материнского организма. Эти мРНК считываются с материнских генов заблаговременно, в процессе созревания яйцеклетки. Именно они обеспечивают синтез белков, необходимых для ранних стадий онтогенеза. В определенный момент материнские мРНК начинают уничтожаться. Это происходит как раз тогда, когда зародыш начинает сам производить мРНК, то есть включает свои гены. Этот довольно быстрый процесс замены зародышем материнских мРНК на свои собственные называется maternal-zygotic transition (MZT).
Менее ясен вопрос о том, что движет процессом MZT. Предполагается три возможных механизма.
1. По мере роста числа клеток, из которых состоит зародыш, начинает сказываться нехватка тех веществ (что бы они из себя ни представляли), которые не позволяют генам зародыша включиться. Ранние стадии эмбриогенеза животных не случайно называют «дроблением»: зигота именно дробится, клетки эмбриона после каждого деления становятся все мельче, поскольку между клеточными делениями отсутствует стадия роста клеток. Общее количество цитоплазмы не растет, тогда как количество клеточных ядер, а следовательно и ДНК, увеличивается в геометрической прогрессии. Если предположить, что яйцеклетка заранее запаслась какими-то ингибиторами транскрипции, то количество этих гипотетических ингибиторов, приходящихся на каждую клетку, должно быстро убывать, и в конце концов их остается так мало, что они уже не могут сдерживать транскрипцию.
2. Не исключено, что в зиготе изначально имеет место целенаправленное блокирование некоторых ключевых генов, работа которых инициирует транскрипцию. Так, показано, что искусственное введение в эмбрион мощных активаторов транскрипции (транскрипционных факторов «широкого спектра действия») может вызвать преждевременное частичное включение эмбрионального генома.
3. Наконец, сама по себе быстрая череда клеточных делений может мешать транскрипции. Ведь каждому делению должно предшествовать удвоение ДНК (репликация). В ходе дробления репликация должна происходить по сути дела непрерывно. Между тем известно, что репликация может мешать транскрипции, а во время клеточного деления (митоза) могут происходить обрыв и уничтожение тех мРНК, синтез которых еще не закончился. Может быть, клетки эмбриона просто физически не успевают транскрибировать свои гены? Искусственное замедление процесса дробления действительно может вызвать преждевременное включение эмбрионального генома. Кстати, у млекопитающих дробление протекает сравнительно медленно, не этим ли объясняется раннее включение генов зародыша?
Впрочем, ни одна из этих теорий не объясняет всей совокупности имеющихся фактов. Например, они не могут объяснить, почему эмбриональные гены включаются не все сразу, а постепенно, в строго определенном порядке.
Вторым существенным аспектом MZT является уничтожение материнских мРНК. Тут, как выяснилось, все предусмотрено заранее: материнские мРНК помечены особой последовательностью нуклеотидов, расположенной на нетранслируемом (то есть не кодирующем белок) «хвостике» этих молекул. Среди первых генов, которые эмбрион включает в процессе MZT, находятся гены особых белков и маленьких РНК, которые распознают эту последовательность, прикрепляются к ней и тем самым инициируют уничтожение материнских мРНК.
До полного понимания всех этих процессов науке еще очень далеко, но дело движется. Когда основные механизмы генной регуляции в ходе раннего онтогенеза будут расшифрованы, биологи смогут вплотную заняться следующим принципиальным вопросом, а именно — зачем все это надо? Почему животные не доверяют своему геному контроль над ранними стадиями развития, а потом в какой-то момент вдруг «переключаются» с материнских мРНК на свои собственные?
Между прочим, эта проблема имеет не только теоретическое, но и практическое значение. Трудности, с которыми сталкиваются работы по клонированию животных, во многом определяются тем, что мы еще слишком мало знаем о работе генов в ходе раннего развития. Клонирование животных осуществляется путем пересадки ядра из соматической (неполовой) клетки одного животного в яйцеклетку другого. Однако для нормального развития необходимо, чтобы гены зародыша поначалу помалкивали, а они в донорском ядре вполне активны. Цитоплазма яйцеклетки должна каким-то образом «перепрограммировать» ядро, выключить геном, а потом в нужный момент снова включить его. Пока мы не знаем, как ей в этом помочь, больших успехов в клонировании ожидать трудно.
—————
Разгадан механизм регенерации конечностей. «Генетическая программа» индивидуального развития очень тесно связана с другой «программой», на основе которой происходит регенерация — заживление ран и отращивание утраченных частей тела. Изучая регенерацию, ученые одновременно проникают в тайны онтогенеза и наоборот.
Биологи из Испании и США недавно провели серию генно-инженерных экспериментов, в результате которых им удалось показать, что регенерация конечностей у позвоночных идет под управлением тех же регуляторных белков, которые руководят развитием конечностей у эмбриона. Белки, о которых идет речь, образуют два сигнально-регуляторных каскада, которые называются Wnt/beta-catenin (о роли белка Wnt в развитии червя Caenorhabditis elegans говорилось выше в сюжете «Развитие червей начинается с хвоста») и BMP (по названию ключевого участника каскада — белка BMP, bone morphogenetic protein).
Для первого эксперимента ученые сконструировали специальный вирус, в геном которого был встроен ген белка, блокирующего работу Wnt-каскада. Введение вируса аксолотлю снизило способность к регенерации. У аксолотля в норме отрезанные конечности восстанавливаются полностью, однако у зараженных искусственным вирусом аксолотлей вместо лапы вырастала лишь заостренная культя без пальцев.
Этот эксперимент показал, что нормальная работа Wnt-каскада — необходимое условие регенерации, но является ли она также и достаточным условием? Чтобы проверить это, ученые поставили эксперимент на шпорцевой лягушке, у которой в отличие от аксолотля способность к регенерации есть только у головастиков, а с возрастом эта способность теряется. Был сконструирован еще один вирус, производящий белок бета-катенин — один из ключевых участников Wnt-каскада. Введение этого вируса улучшало способность шпорцевых лягушек к регенерации конечностей на тех стадиях развития, когда эта способность уже ослаблена. Однако у взрослых лягушек, полностью потерявших способность к регенерации, этого не наблюдалось. Следовательно, для регенерации необходимо что-то еще, кроме Wnt-каскада.
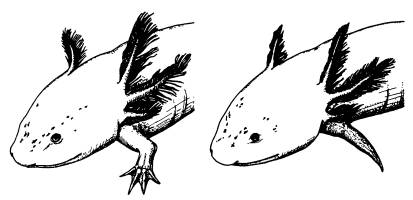
Нормально регенирировавмая после ампутации конечность аксолотля (слева) и недоразвитая конечность у особи, которой был введен Axin1 — ингибитор Wnt-каскада.
Третьим подопытным объектом стала рыбка данио (Danio rerio). В норме эта рыбка способна заново отращивать утраченные плавники. Ранее было показано, что различные ингибиторы Wnt-каскада снижают эту способность (так же как у аксолотля и шпорцевой лягушки). Чтобы проверить, способно ли активирование Wnt-каскада усилить способность к регенерации у данио, исследователи использовали рыбок, потерявших эту способность в результате мутации. Введение таким рыбкам бета-катенинового вируса привело к частичному «исправлению» данного дефекта. На данио исследователям удалось также показать, что для успешной регенерации помимо Wnt-каскада должен нормально работать и сигнально-регуляторный каскад BMP.
Самое интересное, что оба каскада (Wnt и BMP) руководят и развитием конечностей у эмбрионов. На клеточно-тканевом уровне оба процесса — регенерация и развитие конечности в эмбриогенезе — тоже очень сходны. Все эти данные указывают на то, что восстановление утраченных конечностей осуществляется на основе той же самой программы развития, которая руководит формированием конечностей у эмбриона. И эту программу, по-видимому, можно искусственно «включить» даже у тех животных, которые в норме вообще не способны восстанавливать утраченные конечности. Ведь программа эмбрионального развития есть у всех!
Ученым удалось подтвердить это предположение в опытах на курином эмбрионе. Удаление особого многослойного эпителия, расположенного на верхушке зачатка крыла, приводит к недоразвитию крыла. Однако оказалось, что, если активировать у цыпленка ген бета-катенина, крыло после такой ампутации развивается гораздо лучше.
Исследователи сделали из всего этого осторожный вывод, что сигнально-регуляторные каскады, управляющие развитием зародыша, вероятно, стали тем «сырым материалом», на основе которого у некоторых животных развилась способность к регенерации. Не исключено, что дальнейшее изучение этих регуляторных систем позволит в будущем «включать» регенерацию у взрослых животных, полностью лишенных способности к восстановлению утраченных частей тела, в том числе и у человека.
(Источник: Yasuhiko Kawakami et al. Wnt/beta-catenin signaling regulates vertebrate limb regeneration // Genes & Development. 2006. V. 20. P. 3232–3237.)
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Для чего нужны ангелы
Для чего нужны ангелы Увлекшись объяснениями минерализации скелета, мы забыли, что события, произошедшие в начале кембрийского периода, нельзя сводить только к обретению скелета.Последовательные изменения в размерах вендских и кембрийских планктонных
Для чего человеку нужны ненужные (рудиментарные) органы
Для чего человеку нужны ненужные (рудиментарные) органы Разумеется, никто никогда не сомневался в том, что человек не тождествен с человекообразными обезьянами и что он отличается от них многими особенностями скелета и других органов. Но эти отличия не настолько
Что такое протеины и зачем они нужны организму?
Что такое протеины и зачем они нужны организму? Протеины, чаще называемые белками, представляют собой высокомолекулярные органические соединения, построенные из аминокислот. При образовании белковой молекулы аминокислоты соединяются в длинные пептидные нити, которые
Что такое углеводы, зачем они нужны организму и в каких продуктах содержатся?
Что такое углеводы, зачем они нужны организму и в каких продуктах содержатся? Углеводы (сахара) – обширная группа природных соединений, химическая структура которых часто отвечает общей формуле Cm(H2O)n (то есть углерод плюс вода, отсюда название). Углеводы являются
Зачем нужны два пола?
Зачем нужны два пола? В процессе эволюции мужские и женские организмы играют разные роли. Проблему неравноценности полов подробно исследовал российский ученый В.А. Геодакян, который показал целесообразность этого явления для существования вида. На всех стадиях полового
Для видообразования нужны популяции, а не отдельные особи
Для видообразования нужны популяции, а не отдельные особи Селекция — вот путь, которым, по мнению большинства ученых, волки были превращены в собак. Однако эволюционирует не отдельная особь, а популяция, поэтому для видообразования нужно было, чтобы на протяжении
Доминантные гены
Доминантные гены Присутствие доминантного гена всегда явно и внешне проявляется соответствующим признаком. Поэтому доминантные гены, несущие нежелательный признак, представляют для селекционера значительно меньшую опасность, чем рецессивные, так как их присутствие
Глава 19. ДЛЯ ЧЕГО ЖЕ ОНИ НУЖНЫ?
Глава 19. ДЛЯ ЧЕГО ЖЕ ОНИ НУЖНЫ? Теперь, после того, как мы подробно выяснили, почему присутствие ферментов в ящерице и их отсутствие в камне так важно, разумно поинтересоваться: какие же именно химические реакции катализируют ферменты, если уж эти реакции являются
Лемминги нужны всем!
Лемминги нужны всем! Почти над всей тундрой и над всеми островами и морями, которые севернее ее, в декабре-январе — полная полярная ночь.Месяцами не видно солнца. Над заснеженной равниной, над белыми холмами, над глыбами полярных льдов безграничным кажется царство
1 От генетики к эпигенетике: почему генам нужны переключатели
1 От генетики к эпигенетике: почему генам нужны переключатели Книга жизни До конца срока на посту президента США Уильяму Джефферсону — или просто Биллу — Клинтону остается еще несколько месяцев. Он входит в церемониальный зал Белого дома. Размеренным шагом приближается
Гены жаберных дуг
Гены жаберных дуг В течение первых нескольких недель после зачатия в клетках жаберных дуг зародыша и во всех тканях, из которых впоследствии образуется наш мозг, последовательно включаются и выключаются целые батареи генов. В соответствии с инструкциями, записанными в
Гены
Гены Открытие Арендта подводит нас еще к одному вопросу. Одно дело, что у глаз разных животных есть общие части, но как получилось, что такие непохожие друг на друга глаза, как у червей, мух и мышей, родственны друг другу? Чтобы ответить на этот вопрос, обратимся к