Десять тысяч лет, три нуклеотида и защитная окраска
Десять тысяч лет, три нуклеотида и защитная окраска
Американские белоногие хомячки, называемые также оленьими мышами (Peromyscus maniculatus), в норме имеют темную окраску, однако представители этого вида, обитающие в районе с очень светлой почвой (Песчаные Холмы в штате Небраска), окрашены светлее своих сородичей. Хомячки с Песчаных Холмов стали хрестоматийным примером эволюции адаптивных признаков. Этому способствовали два обстоятельства. Во-первых, адаптивность (полезность) светлой окраски не вызывает сомнений: хищные птицы гораздо лучше видят на светлом фоне темного хомячка, чем светлого. Во-вторых, Песчаные Холмы — молодое геологическое образование: они сформировались после отступления ледника около 10 тыс. лет назад. Это дает основания думать, что мы имеем дело с адаптацией, появившейся — в эволюционном масштабе времени — совсем недавно. Однако для понимания механизмов адаптации важно выяснить ее генетические основы.
Известно, что окраска шерсти млекопитающих зависит от распределения двух пигментов: черно-коричневого эумеланина и рыжего феомеланина. Клетки меланоциты, отвечающие за окраску волоса, могут поочередно синтезировать то один, то другой пигмент по мере роста волоса. В результате волос получается неоднородно окрашенным. Часто кончик и основание волоса темные (эумеланиновые), а посередине имеется светлая (феомеланиновая) полоса. И вот любопытно, что же определяет разницу в окраске у темных и светлых хомячков? Если вглядеться попристальней, то окажется, что никаких специальных ухищрений и особой цветовой палитры не требуется. Всего лишь нужно в каждой шерстинке светлых хомячков расширить феомеланиновую полосу. И больше ничего! Элементарно! И вот американские генетики из Гарвардского и Калифорнийского университетов решили изучить, что же стоит за этим элементарным решением (Linnen et al., 2009).
Известно, что на окраску шерсти может влиять ген Agouti. Сигнальный белок, кодируемый этим геном, подает команду меланоцитам синтезировать феомеланин вместо эумеланина. Влияние мутаций Agouti на окраску шерсти подробно изучено на домовых мышах (Mus musculus). Оказалось, что мутанты с отключенным геном Agouti имеют черную окраску, а повышенная активность гена приводит к очень светлой окраске. Светлый аллель — доминантный по отношению к темному.
Чтобы проверить, действительно ли светлая окраска хомячков с Песчаных Холмов определяется доминантной мутацией, ученые скрещивали разномастных родителей. Все потомство от скрещивания получилось светлым, и эта гипотеза подтвердилась.
Дальнейшие эксперименты показали, что непосредственным результатом мутации является повышенная активность Agouti в первую неделю жизни хомячат, т. е. в тот период, когда у них отрастает шерсть. Максимальная активность Agouti приходится на четвертый день жизни как у темных, так и у светлых хомячков, однако абсолютная величина этой активности (измеряемая по количеству матричных РНК, считанных с гена) у светлых особей с Песчаных Холмов в несколько раз выше.
После была проведена весьма трудоемкая и кропотливая работа — секвенирование нуклеотидной последовательности Agouti у сотни хомячков из темных и светлых лабораторных линий, а также у диких зверьков из зоны смешения темной и светлой популяций. В результате было выявлено около 20 полиморфных сайтов, т. е. таких участков гена, которые не у всех особей одинаковы. Существуют эффективные методы статистического анализа нуклеотидных последовательностей, которые позволяют обнаруживать следы действия положительного отбора на те или иные участки генов. Иными словами, отличать изменения ДНК, которые поддерживались отбором (т. е. полезные, адаптивные), от нейтральных изменений, которые распространялись в популяции за счет дрейфа. Применив эти методы, ученые пришли к выводу, что ключевая мутация, в результате которой мыши приобрели светлую окраску, находится только в одном месте, а изменения в остальных 19 полиморфных сайтах тоже внесли свой вклад, но менее значительный.
Эта мутация находилась лишь в одном из 20 сайтов. Она состояла в выпадении трех нуклеотидов, кодирующих аминокислоту серин. Удивительно, что такой важный признак, как маскирующая окраска, может определяться всего тремя нуклеотидами! Какая поразительная несоразмерность: с одной стороны, три молекулы, их даже в микроскоп не видно, а с другой стороны — невидимость для врага, долгая жизнь, многочисленная семья и потомство… Когда сравниваются такие категории, то кажется, что три молекулы — это ничто, не могут они определить хомячьего счастья. Но эта несоразмерность — мнимая.
—————
Следы естественного отбора
Влияние естественного отбора на генофонд популяции трудно наблюдать в природе из-за медленности процесса. Непосредственно наблюдать отбор, действовавший на популяцию в прошлом, и вовсе невозможно без машины времени. К счастью, это и не обязательно, потому что отбор, нацеленный на тот или иной локус (участок молекулы ДНК), оставляет в геномах легкоразличимые следы. Мы можем их увидеть, потому что разрешающая способность молекулярного метода исследования теперь фантастическая — один нуклеотид. Еще 15–20 лет назад это казалось чудом, волшебством. Геномы изучали с помощью скрещиваний мутантов, составляли генные карты, разрешение такого метода было не выше частей хромосом или групп генов, но уж никак не одного нуклеотида. Это все равно что заменить 20-кратную лупу на электронный микроскоп. И если уж эволюционные механизмы работают с наследственным материалом — последовательностями ДНК и РНК, то при такой разрешающей способности молекулярного метода мы это увидим.
Следы работы эволюционных механизмов бывают двух типов. Каждая аминокислота кодируется или одним, или, чаще, несколькими определенными тройками (триплетами) нуклеотидов. Если аминокислота кодируется одним-единственным триплетом, то замена любого нуклеотида в триплете приведет к замене аминокислоты; если она кодируется несколькими триплетами, то замена одного нуклеотида может привести либо к замене аминокислоты, либо к замене триплета другим из возможного набора, а аминокислота останется той же. Замена нуклеотида без замены аминокислоты называется синонимичной, а если аминокислота меняется, то получим несинонимичную, или значимую, замену. Если, сравнивая какой-то ген у двух организмов, мы видим, что среди наблюдаемых различий преобладают значимые замены, это указывает на действие положительного отбора, который поддерживал полезные мутации в этом гене. Если, наоборот, резко преобладают синонимичные замены, а значимых нет или очень мало, — это след отрицательного отбора, который отбраковывал все значимые мутации, потому что они оказывались вредными. Промежуточное соотношение указывает на нейтральную эволюцию. Таким образом, отношение синонимичных и несинонимичных замен свидетельствует о направлении отбора, и это первый тип «следов», оставляемых в геноме естественным отбором.
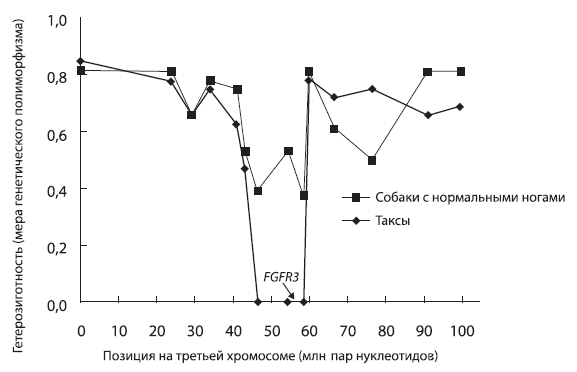
Пример «селективного выметания». У такс по сравнению с обычными собаками резко понижен генетический полиморфизм на участке третьей хромосомы, включающем ген FGFR3. Это указывает на то, что отбор, осуществлявшийся собаководами, выводившими новую породу коротконогих охотников на лис (было это менее 100 собачьих поколений назад), благоприятствовал какой-то мутации на этом участке хромосомы. В условиях отбора на «таксовость» некая мутация, произошедшая в этом участке, оказалась для собак «полезной», поскольку она обеспечивала успешное размножение, тогда как щенки, не имевшие этой мутации, отбраковывались селекционерами. Что же это за ген — FGFR3? Имеет ли он какое-то отношение именно к таксам? Оказывается, имеет. Белок, кодируемый этим геном, называется рецептором фактора роста фибробластов. Мутации в нем приводят к разнообразным нарушениям развития скелета, в том числе к укорочению конечностей. По рисунку из Pollinger et al., 2005.
Второй тип следов выявляется при изучении уровня полиморфизма (разнообразия) нуклеотидных последовательностей в популяции. Если мы отсеквенируем геномы множества представителей какого-нибудь вида и сравним их между собой, то увидим, что уровень полиморфизма, т. е. индивидуальных различий между геномами, распределен по геному неравномерно. Где-то полиморфизм выше — это те участки генома, в которых большинство мутаций оказываются нейтральными, поэтому полиморфизм свободно накапливается. Скорее всего, это просто не очень нужные участки, «генетический балласт». Где-то генетическое разнообразие ниже — это важные участки генома, в которых большинство мутаций оказываются вредными и удаляются очищающим отбором. Ну а в некоторых местах — и это самое интересное — мы увидим резкое, чуть не до нуля, снижение полиморфизма. В таких участках нет (или очень мало) не только значимых, но и синонимичных различий. Как правило, это означает, что здесь поработала «метла» положительного отбора. Это явление так и называют — selective sweep, т. е. «выметание посредством отбора». В середине участка с резко пониженным полиморфизмом обычно сидит какая-то полезная мутация. Она возникла не очень давно у какой-то особи, а потом быстро распространилась под действием положительного отбора. Вместе с мутацией автоматически распространялись и прилегающие к ней участки ДНК. Это явление называют сцепленным наследованием. До появления полезной мутации уровень полиморфизма в данном участке хромосомы был, скорее всего, примерно таким же, как на соседних участках. Но, когда отбор начинает распространять полезную мутацию, он автоматически распространяет и ее окружение со всеми индивидуальными (и в основном нейтральными) особенностями, присущими вовлеченному в сценарий участку ДНК. Повезет тем нейтральным аллелям, которые находились рядом с полезной мутацией. Остальные нейтральные вариации исчезнут из генофонда, а выживут те, что были у счастливого обладателя первого экземпляра мутантного гена. Как будто все варианты, кроме одного, из данного фрагмента генома «выметаются».
Со временем следы метлы стираются за счет накопления новых нейтральных мутаций. Таким образом, глубокие ямы на графике распределения полиморфизма указывают на относительно недавние случаи действия положительного отбора.
—————
Как в первую неделю жизни хомячат работает мутантный сигнальный белок Agouti — пока неизвестно. Зато удалось показать, что данная мутация, по всей видимости, появилась и начала распространяться в популяции совсем недавно — позже, чем отступил ледник и сформировались Песчаные Холмы с их светлым грунтом. В пользу этого свидетельствуют результаты статистических тестов. В частности, оказалось, что «светлые» варианты гена Agouti (в которых отсутствуют вышеупомянутые три нуклеотида) меньше варьируют по остальным полиморфным сайтам, чем «темные» варианты. Уровень полиморфизма в окрестностях полезной мутации относительно низок. Это типичный пример «выметания посредством отбора», и этого не должно было бы наблюдаться, если бы данная мутация (выпадение трех нуклеотидов) существовала в популяции в качестве нейтральной задолго до того, как появились Песчаные Холмы и она стала полезной.
Исследование показало, что быстрое формирование новых адаптаций может происходить за счет новых мутаций, которые появляются уже после того, как в них «возникла потребность». В тот момент, когда условия среды (а значит, и направление отбора, действующего на популяцию) вдруг меняются, в популяции может не оказаться подходящих генетических вариантов, которые были до сих пор нейтральными, а теперь стали полезными. Скорее всего, светлая окраска была не нейтральной, а однозначно вредной для хомячков, пока они жили в районах с темной почвой. Но, когда образовались Песчаные Холмы — подходящая для жизни хомячков территория со светлой почвой, — ситуация сразу изменилась, и изредка появляющиеся в популяции светлоокрашенные мутанты, которых отбор до сих пор безжалостно отсеивал, получили свой шанс.
—————
Отбору все равно
Есть другие случаи адаптивных изменений окраски, где механизм адаптации на молекулярном уровне хорошо изучен. Один из таких примеров — скальные щетинистые прыгуны (Chaetodipus intermedius), грызуны, обитающие в пустынях юго-запада США, где участки светлого грунта чередуются с черными лавовыми полями. Как читатели уже догадались, на светлых участках преобладают светлоокрашенные животные, на лавовых полях — темноокрашенные. Показано, что отбор в данном случае осуществляется хищными птицами, в том числе совами, которые лучше видят на темном фоне светлую добычу, а на светлом — темную. Самое интересное, что у обитателей разных лавовых полей отбор зафиксировал разные мутации, приводящие к одному и тому же фенотипическому эффекту — темной шерсти. На одном из лавовых полей все черные грызуны несут мутацию в гене меланокортинового рецептора MC1R (это один из генов, регулирующих синтез черного пигмента эумеланина)[26]. У прыгунов, обитающих на лавовых полях, удаленных на 700 км от первого, черная окраска определяется мутациями в других генах (Majerus, Mundy, 2003).
Отбору все равно, мутацией какого гена вызвано изменение окраски. Никто не проектирует эволюционное изменение, все происходит само собой. Если возникает мутация, фенотипический эффект которой здесь и сейчас повышает эффективность размножения (приспособленность) ее носителей, то оные носители, эффективно размножаясь, передают мутацию по наследству своим потомкам. А это автоматически ведет к росту частоты встречаемости данной мутации в генофонде. Вот и все. Как видите, можно точно описать дарвиновский эволюционный механизм, не используя слово «отбор» (дабы не вводить читателя лишний раз в искушение, ведь в привычных метафорических формулировках типа «отбор поддерживает» или «отбору все равно» легко усмотреть то, чего там на самом деле нет, — приписывание отбору свойств разумного деятеля, имеющего какие-то цели и планы).
—————
Перед нами классический пример адаптации животных к внезапно изменившимся природным условиям. Чтобы приобрести полезный признак, хомячкам потребовалось изменить всего один ген, а если еще точнее — всего лишь три нуклеотида в нем. И в результате мутации хомячки обрели защитную окраску. Эта мутация, как и в случае с культурным рисом, стала полезной после изменения условий среды и направленности отбора, а до того она была для организмов вредной. В геноме остались следы работы отбора, и мы умеем их находить и анализировать. И мы видим, что все случилось не по взмаху волшебной палочки, а здесь действовал мелочный и постепенный положительный отбор.
Но полезная мутация может быть связана не только с дефектом того или иного потерявшего актуальность гена, как в случае с рисом, или с изменением активности гена на определенной стадии развития организма, как в случае с хомячками. Очень своевременной может стать и замена аминокислоты в активном центре фермента, в результате которой фермент приобретает способность работать с новыми веществами. Следующий пример — как раз из этой серии.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Окраска и украшения
Окраска и украшения Окраска этих крошечных птичек чрезвычайно пестра и красива. Основным тоном, так же как у попугаев, можно признать зеленый; но в то время как попугаям свойственна зелень шелковистого отлива, зелень колибри всегда металлически блестяща. Зеленый цвет
В крови плавают десятки тысяч различных антител, но они имеют общий план строения.
В крови плавают десятки тысяч различных антител, но они имеют общий план строения. — Как устроены молекулы антител? Это, наверное, белки, которые циркулируют в крови. — Да, это самые крупные белковые молекулы кровяной сыворотки — гамма–глобулины. Поскольку все
Окраска рыб
Окраска рыб Окраска рыб очень разнообразна. В дальневосточных водах обитает мелкая (8–10 сантиметров[1]), похожая на снетка лапша-рыба с бесцветным, совершенно прозрачным телом: через тонкую кожицу просвечивают внутренности. Около морского берега, где так часто вода
Глава 6 Десять лишних заповедей
Глава 6 Десять лишних заповедей Две вещи наполняют мою душу всегда новым и все более сильным изумлением и благоговением, чем чаще и продолжительнее я размышляю о них: это звездное небо надо мной и моральный закон во мне. Иммануил Кант Можем ли мы не чувствовать боли, когда
ДЕСЯТЬ ЛЕТ СПУСТЯ
ДЕСЯТЬ ЛЕТ СПУСТЯ НЕ ТОЛЬКО на следующий год, но и через два, три, четыре года ничто не выдает присутствия термитов в их подземном убежище. Оно все еще незаметно извне. Зарывшиеся на глубину нескольких сантиметров основатели гнезда теперь уже не одиноки. Их окружают первые
Глава 1. Десять самых вредных заблуждений
Глава 1. Десять самых вредных заблуждений Заблуждение No1. Силовой спорт = анаболические стероиды Прежде чем мы познакомимся с принципами эффективных тренировок, я хочу рассмотреть наиболее вредные заблуждения, касающиеся силовых видов спорта.Первое заблуждение,
Глава 2. Десять принципов эффективного тренинга
Глава 2. Десять принципов эффективного тренинга Без труда не вытащишь и рыбку из пруда, а без пруда не вытащишь рыбку и с трудом.Я больше не буду перечислять заблуждения, которые обрушиваются на голову начинающего атлета со страницы красочных журналов, а просто изложу
Сто тысяч долларов за шубу!
Сто тысяч долларов за шубу! «Ванна из золота или шуба?» — нелегкий этот вопрос, возможно, задавали себе немногие, но зато очень богатые женщины. И шуба из шиншиллы, и ванна из чистого золота стоят одинаково дорого. Многие миллионерши купаются в золоте, но шиншилловые шубки
Сорок тысяч полевок и мышей
Сорок тысяч полевок и мышей Нет, это не ругательство. 40 тысяч полевок и мышей насчитали счетчики лесной переписи населения на одном квадратном километре по-летнему зеленого массива. Цифра внушительная. Невольно вообразишь, что, отправившись на тот самый квадратный