У бесполых организмов убыль выше прибыли
У бесполых организмов убыль выше прибыли
До сих пор мы вели разговор о преимуществах полового и бесполого размножений для особей одного и того же вида. Но в природе сравниваются по приспособленности не только особи одного вида между собой, но и особи разных видов, занимающие более или менее сходные условия. Здесь особенно важно успеть вперед конкурента — занять часть ареала, заполнить как можно большее пространство. В этой ситуации тот вид, который дает большее число потомков, должен бы получить территориальное преимущество и вытеснить медленного соперника. А как мы уже уяснили, быстрее размножаются бесполые и самооплодотворяющиеся организмы. Пусть они в итоге окажутся хуже приспособленными, но зато их раздельнополым конкурентам просто негде и некогда будет развернуться. Почему тогда бесполые организмы не вытеснили полностью раздельнополых? На этот вопрос помогают ответить примеры с растениями.
Семейство пасленовых — на редкость удобный объект для изучения эволюционных преимуществ полового и бесполого размножения, именно с ним и работали американские и британские биологи (Goldberg et al., 2010). Во-первых, пасленовые — очень разнообразная группа и к тому же прекрасно изученная. Во-вторых, у пасленовых имеется очень подходящий признак — самонесовместимость. Самонесовместимость, или отторжение родственной пыльцы, — свойство, предохраняющее растения от самооплодотворения, заставляющее их смешивать свои гены с генами других особей. Самонесовместимость интересна тем, что распространена среди пасленовых довольно хаотично. Во многих родах пасленовых одни виды имеют систему самонесовместимости, тогда как другие виды того же рода ее не имеют. При этом наличие или отсутствие самонесовместимости не коррелирует с другими признаками этих растений. Ученые задались вопросом, существует ли корреляция между самонесовместимостью и темпами диверсификации.
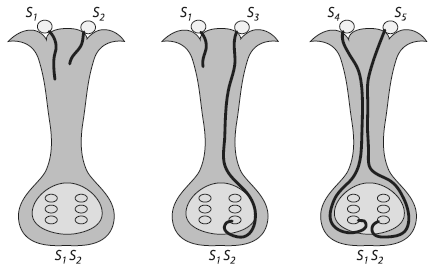
Схема механизма самонесовместимости у растений. Если аллельное состояние гена S («локуса несовместимости») совпадает у пыльцевого зерна и пестика, на чье рыльце оно попало, прорастание пыльцы блокируется, и оплодотворение не происходит. Геном пыльцы гаплоидный, взрослого растения — диплоидный, поэтому у каждого пыльцевого зерна обозначен один аллель S-локуса, а у пестика — два.
В семействе пасленовых около 2700 видов, из которых 41 % имеет систему самонесовместимости, 57 % ее не имеют, а 2 % видов — двудомные, т. е. имеющие отдельные мужские и женские растения, так что для них проблема самооплодотворения неактуальна. Авторы построили эволюционное дерево для 356 видов пасленовых, по которым есть молекулярные данные (дерево строилось по последовательностям двух ядерных генов и четырех пластидных) и для которых установлено наличие или отсутствие самонесовместимости.
Анализ получившегося дерева показал, что самонесовместимость унаследована пасленовыми от общего предка и с тех пор многократно утрачивалась в разных эволюционных линиях. Утратить эту систему легко, а восстановить — трудно, потому что это сложный молекулярный комплекс, состоящий из множества специализированных белков. В эволюции пасленовых, по-видимому, не было случаев восстановления самонесовместимости после ее утраты.
Понятно, почему самонесовместимость часто утрачивается. Переход к самооплодотворению дает немедленное преимущество в эффективности распространения своих генов. К тому же самооплодотворение выгодно, когда с доставкой пыльцы от других особей возникают трудности — например, из-за большой разреженности популяции. Непонятно другое: если этот признак часто утрачивается и почти никогда не восстанавливается, почему до сих пор сохранилось так много видов, обладающих системой самонесовместимости?
Расчеты по построенному филогенетическому дереву показали, что темп видообразования у видов, практикующих самоопыление, значительно выше, чем у самонесовместимых. Понятно, что высокая скорость видообразования у самоопыляющихся растений связана с тем, что у них возникшие полезные комбинации аллелей не «размываются». В итоге одно-единственное растение с подходящей комбинацией аллелей, попавшее в необычные условия, способно дать начало новому виду.
Однако темп вымирания у них еще выше. Почему они чаще вымирают, тоже понятно: ведь у них из-за самоопыления должны быстрее накапливаться вредные мутации и реже фиксироваться мутации полезные. Так что итоговое число самоопыляющихся видов не повышается. Несмотря на то что совокупность самонесовместимых видов убывает за счет превращения их в самоопыляющиеся, число самонесовместимых видов не снижается. Этому препятствует то обстоятельство, что самонесовместимые виды реже вымирают, передавая видам-потомкам самонесовместимость по наследству. Темп вымирания у них не превышает темп появления новых видов. В результате такого соотношения вымирания и появления и даже с учетом перехода некоторых в группу бесполых штрейкбрехеров число самонесовместимых видов остается на постоянном уровне (порядка 30–40 %).
Исследование показало, что естественный отбор может действовать не только на уровне генов и особей, но и на уровне целых видов. Такой отбор способен длительно сохранять сложный признак, который в каждой отдельно взятой эволюционной линии имеет тенденцию исчезать и почти никогда не появляется вновь. Но нужно помнить, что медлительный и малоэффективный межвидовой отбор может лишь поддерживать такой признак, но не может его создать. Подобным творческим потенциалом обладает только отбор на более низких уровнях: генов, особей или, в некоторых особых случаях, групп особей[47].
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
ЧЕМ ВЫШЕ ЛЕЗЕТ ОБЕЗЬЯНА, ТЕМ ЛУЧШЕ ВИДЕН ГОЛЫЙ ЗАД
ЧЕМ ВЫШЕ ЛЕЗЕТ ОБЕЗЬЯНА, ТЕМ ЛУЧШЕ ВИДЕН ГОЛЫЙ ЗАД Тут уместно сказать, что у человека запретна вся сфера зада, а также многое с ней связанное, например публичная дефекация, чего нет у животных. Зад, промежность и свои половые органы человек издревле прятал под
У какого телескопа выше разрешающая способность – радио– или оптического?
У какого телескопа выше разрешающая способность – радио– или оптического? Недостатком радиотелескопов долгое время была их низкая разрешающая способность, достигавшая даже у больших радиотелескопов лишь нескольких минут дуги. Проблема была решена посредством
Какая птица летает выше всех?
Какая птица летает выше всех? Рекордсменами по высоте полета среди птиц являются грифы, одного из которых наблюдали на высоте 11 275 метров над уровнем
В интересах прибыли: отказ от производства натуральных продуктов и замена их опасными субстанциями
В интересах прибыли: отказ от производства натуральных продуктов и замена их опасными субстанциями Препарат производства Hoffman-La Roche, широко используемый в борьбе против малярии под названием «лариам», пережил трудные времена.Однако санитарные власти Швейцарии сделали
11.4. Взаимоотношения организмов в природе
11.4. Взаимоотношения организмов в природе Популяционные взаимодействия определяются взаимоотношениями между отдельными особями популяций. Совокупность таких взаимоотношений и составляют действующие на популяцию биологические
2.2. Адаптации организмов
2.2. Адаптации организмов Приспособления организмов к среде носят название адаптации. Под адаптациями понимаются любые изменения в структуре и функциях организмов, повышающие их шансы на выживание.Способность к адаптациям – одно из основных свойств жизни вообще, так как
7.3. Отношения организмов в биоценозах
7.3. Отношения организмов в биоценозах Основу возникновения и существования биоценозов представляют отношения организмов, их связи, в которые они вступают друг с другом, населяя один и тот же биотоп. Эти связи определяют основные условия жизни видов в сообществе,
Агробизнес делает прибыли на страхе перед птичьим гриппом
Агробизнес делает прибыли на страхе перед птичьим гриппом Мало того, что министр обороны Рамсфелд оказался прямым благотворителем правительств США, Великобритании и других, пополняя их запасы своим «Тамифлю», паника перед птичьим гриппом также пошла в дело, чтобы
Чем выше лезет обезьяна, тем лучше виден голый зад
Чем выше лезет обезьяна, тем лучше виден голый зад Тут уместно сказать, что у человека запретна вся сфера зада, а также многое с ней связанное, например, публичная дефекация, чего нет у животных. Зад, промежность и половые органы человек издревле прятал под набедренными
Выше вы говорили о ревности. Поясните подробнее, чем будет отличаться реакция на измену у НР и ВР мужчины
Выше вы говорили о ревности. Поясните подробнее, чем будет отличаться реакция на измену у НР и ВР мужчины Если говорить об инстинктивно-обусловленных действиях, то любой мужчина в такой ситуации захочет вернуть свою монополию на обладание женщиной. Не этой – так другой.
4. Размножение организмов
4. Размножение организмов Организмы обладают важным биологическим свойством — способностью размножаться, т. е. воспроизводить потомство. Благодаря этой способности поддерживается и продолжается существование всех форм живых существ. Через размножение же