Новые белки из старого конструктора
Новые белки из старого конструктора
Большинство белковых молекул состоит из нескольких функциональных блоков (доменов). Например, белки, участвующие в передаче сигналов, часто содержат домены двух типов — регуляторные и каталитические. Первые реагируют на поступающие сигналы и определяют условия, при которых белок-передатчик «сработает». Вторые передают сигнал следующему участнику каскада (например, присоединяют фосфатную группу к другому белку, переводя его в активное состояние). Каждый такой белок действует как оператор условного перехода «если… то», причем регуляторный домен отвечает за «если», а каталитический — за «то».
Ученые давно предполагали, что перетасовка фрагментов белковых молекул может быть важным источником эволюционных новшеств (Ратнер, 1993). На это указывают данные сравнительной генетики, а недавно начали появляться и прямые экспериментальные подтверждения. Одно из них получили биологи из Калифорнийского университета в Сан-Франциско, взявшись изучать сигнальный каскад, регулирующий половое поведение дрожжей (Peisajovich et al., 2010).
Дрожжи, как уже говорилось в главе 3, делятся на два «пола»: а и ?. Эксперименты проводились с полом а. Сигналом к спариванию для этих клеток служит альфа-фактор — феромон, выделяемый полом ? (см. рисунок). Феромон взаимодействует с рецептором Ste2, который находится на поверхности клетки а. В результате комплекс из трех белков, прикрепленный к рецептору с внутренней стороны мембраны, распадается. Освободившийся белок Ste4 взаимодействует с белком Ste5, в результате чего Ste5 с прикрепленными к нему протеинкиназами[75] Ste11, Ste7 и Fus3 транспортируется к клеточной мембране. Здесь благодаря посредничеству белка Ste50 устанавливается связь между Ste11 и Cdc42. Последний белок входит в состав комплекса из трех белков, один из которых — киназа Ste20 — активирует белок Ste11, прикрепляя к нему фосфатную группу. Ste11 в свою очередь фосфорилирует киназу Ste7, а она активирует киназу Fus3. Активированный белок Fus3 отправляется в ядро, где он активирует несколько транскрипционных факторов, которые в свою очередь включают комплекс генов, необходимых для спаривания. В клетке приостанавливаются процессы, связанные с ростом и делением, меняется морфология клетки, и кончается все тем, что клетка а сливается со своим половым партнером — клеткой ?.
Эксперименты проводились с 11 белками, которые на рисунке обозначены серыми овалами. Шесть из них состоят более чем из одного домена. Гены многодоменных белков разрезали на части, а фрагменты перекомбинировались случайным образом. В итоге были получены гены 66 новых белков. Эти гены затем поодиночке вставляли в дрожжевые клетки. При этом исходный сигнальный каскад оставляли без изменений, лишь добавляя к нему новых потенциальных участников. Все искусственные гены были соединены с одним и тем же регуляторным участком (промотором), что обеспечивало одинаковый (невысокий) уровень активности внедренных генов.
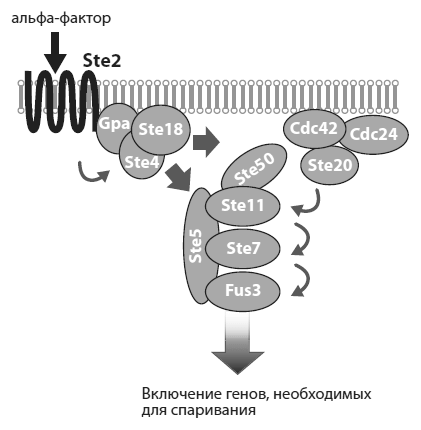
Схема сигнального каскада, запускающего программу «брачного поведения» у дрожжей. Из Peisajovich et al., 2010.
Чтобы оценить эффективность работы сигнального каскада у 66 генно-модифицированных штаммов дрожжей, в их геномы был добавлен ген зеленого флуоресцирующего белка, соединенный с промотором, который реагирует на один из транскрипционных факторов, активируемых белком Fus3. В результате по силе свечения можно было определить силу реакции сигнального каскада на добавление в среду альфа-фактора. Регистрировались два параметра: «базовая» сила свечения, которая наблюдается до добавления альфа-фактора, и скорость, с которой свечение усиливается после добавления феромона.
Оказалось, что в десяти случаях из 66 добавление нового белка изменило поведение сигнального каскада. У одних штаммов изменился базовый уровень активности, у других — интенсивность реакции на феромон.
Но эти изменения теоретически могли быть вызваны не перекомбинированием доменов в добавленном белке, а просто тем, что какого-то домена в клетке стало больше. Чтобы исключить эту возможность, провели контрольные эксперименты. В клетки вставляли дополнительные копии целых генов и их усеченных, но работающих вариантов (кодирующих только один из доменов многодоменного белка). Эти манипуляции, однако, не повлияли на работу сигнального каскада. Значит, полученные в эксперименте новые признаки связаны именно с новыми свойствами белков, образованных путем перекомбинирования доменов.
Приводит ли более интенсивная работа сигнального каскада к реальному повышению половой активности? Чтобы это выяснить, исследователи смешивали модифицированные дрожжи с дикими дрожжами другого пола (альфа) и подсчитывали число успешных слияний. Оказалось, что те клетки, у которых скорость реагирования каскада на феромон увеличилась, действительно спариваются чаще, чем контрольные, и наоборот — те клетки, у которых чувствительность каскада снизилась, спариваются реже.
Рекордсменами оказались дрожжи с искусственным белком, состоящим из половинок Ste50 и Ste7, и с другим химерным белком, собранным из фрагментов Ste5 и Ste11. Эти «половые гиганты» спаривались втрое чаще, чем контрольные исходные дрожжи.
Для некоторых случаев удалось расшифровать молекулярные механизмы возникших изменений. Например, белок, собранный из регуляторного домена Ste5 и каталитического домена Ste11, создал новый «обходной» путь передачи сигнала от рецептора Ste2 к Fus3. Это заставляет клетку интенсивнее реагировать на феромон, что повышает вероятность спаривания.
Может ли повышение половой активности дрожжей, наблюдавшееся в эксперименте, быть полезным? Будет ли оно поддержано отбором, если такая мутация возникнет в природной популяции? Судя по тому, что мы знаем о действии отбора на склонность к сексу (см. главу 3), это возможно, особенно в неблагоприятных и переменчивых условиях. В эксперименте повышение половой активности ничуть не повредило здоровью подопытных клеток.
Такие исследования показывают, что перекомбинирование доменов может быть важным источником эволюционных новшеств — наряду с другими «крупномасштабными» мутациями, такими как дупликация генов и появление новых регуляторных участков ДНК. Важно, что перекомбинирование может приводить к мгновенному появлению новых признаков, тогда как дупликация генов создает новшества не сразу, а лишь после того, как две копии гена хоть немного «разойдутся» по своим функциям, накопив нуклеотидные замены.
—————
Комбинаторика, поставленная на поток
Большинство генов у эукариот состоит из кодирующих участков — экзонов и некодирующих вставок между ними — интронов. Интроны — потомки мобильных генетических элементов, буйно расплодившихся в геномах ранних эукариот. Экзонно-интронная структура генов облегчает создание новых белков комбинаторным путем, хотя возникла она, конечно, не для этого. Тем не менее, раз уж так получилось и гены эукариот приобрели экзонно-интронную структуру, это открыло перед эволюцией интересные возможности.
Интроны необходимо удалить, прежде чем синтезировать белок на основе инструкций, записанных в гене. Если этого не сделать, интроны начнут транслироваться, и вместо рабочего белка получится ерунда. Для удаления интронов из молекул матричных РНК (мРНК), считанных с гена, развился механизм сплайсинга. Мобильные элементы — предки интронов — сами себя вырезали из мРНК. Это были «самосплайсирующиеся» элементы (такие и сейчас есть у некоторых прокариот). Если бы они этого не делали, зараженный ими геном стал бы нежизнеспособным, а вместе с ним погибли бы и они сами. У древних эукариот функция сплайсинга перешла от интронов к специальным молекулярным машинкам — сплайсосомам. В состав сплайсосом входят молекулы РНК — наследники тех частей древних самосплайсирующихся интронов, которые, собственно, и осуществляли самосплайсинг. Интроны, разумеется, должны содержать (и действительно содержат) в своей последовательности нуклеотидов специальные сигнальные участки, по которым сплайсосома распознает интрон и определяет, где у него конец и где начало — иначе интрон не может быть вырезан.
С универсальной системой вырезания интронов у эукариот появилась возможность изготавливать новые белки путем альтернативного сплайсинга. Так называют ситуацию, когда одна и та же молекула мРНК в зависимости от условий комбинируется (на генетическом жаргоне это звучит как «сплайсируется») по-разному. Например, в клетках печени из конкретной мРНК вырезаются только интроны, а в клетках мозга из нее же вместе с двумя соседними интронами вырезается и расположенный между ними экзон. Получается два разных белка, синтезируемых на основе одного и того же гена.
Недавно выяснилось, что альтернативный сплайсинг чаще всего используется у самых сложных организмов, таких как наземные позвоночные, и особенно млекопитающие. Более того, оказалось, что у приматов — к которым мы имеем честь принадлежать — альтернативный сплайсинг развит в полтора-два раза сильнее, чем у других млекопитающих (Barbosa-Morais et al., 2012).
Именно поэтому у человека и других приматов разнообразие белков намного выше, чем у дрозофилы или червя C. elegans, хотя по числу белок-кодирующих генов все эти животные не так уж сильно отличаются друг от друга. Как мы помним из главы 3, нельзя до бесконечности наращивать количество полезных генов в геноме, потому что это повышает риск генетического вырождения. Так вот, похоже на то, что альтернативный сплайсинг стал для высших животных удобным способом увеличивать разнообразие белков, не повышая числа генов в геноме.
Альтернативный сплайсинг позволяет по-разному комбинировать экзоны одного и того же гена. При этом порядок экзонов не меняется: разница только в том, что одни экзоны могут быть выброшены вместе с интронами, а другие оставлены. В некоторых генах даже возникли целые наборы — «кассеты» альтернативных экзонов, причем в зрелую мРНК после сплайсинга попадает только один экзон из каждой кассеты. Об одном из таких генов (DSCAM) мы рассказали в заключительной главе «Рождения сложности».
Однако альтернативный сплайсинг не позволяет комбинировать экзоны, входящие в состав разных генов. Для этого нужны перестройки уже не на уровне мРНК, а на уровне геномной ДНК: нужны транспозиции, т. е. перемещения фрагментов ДНК из одного места генома в другое. Это не редкость — случайные транспозиции являются одним из классических типов геномных мутаций, а для геномных паразитов — транспозонов это и вовсе стиль жизни. Вопрос в том, играет ли перекомбинирование экзонов в геномной ДНК существенную роль в появлении эволюционных новшеств.
У эукариот есть немало белков, скорее всего, возникших именно этим способом. Интересно, что они крайне неравномерно распределены по эволюционному дереву. Их очень много у животных, причем у всех — от губок до млекопитающих. У других эукариот (растений, грибов и протистов) их намного меньше. Функции большинства таких многодоменных белков животных связаны с поддержанием целостности организма (взаимодействие клеток, формирование тканей, прием и передача сигналов, формирование межклеточной среды и т. п.). Похоже, на ранних этапах эволюции животного царства был своеобразный бум формирования новых генов путем перекомбинирования экзонов (Patthy, 1999).
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
2. Белки хроматина
2. Белки хроматина Мы уже знаем, что хроматин состоит из ДНК и гистонов в равном весовом количестве и негистоновых белков (НГБ), которых в неактивных районах хромосомы всего 0,2 веса ДНК, а в активных — более чем 1,2 (в среднем НГБ мепьше, чем ДНК). Мы знаем также, что гистоны
Клетки, белки и гены
Клетки, белки и гены Жизнь есть способ существования белковых тел. Ф. Энгельс Наше тело является империей клеток, каждая из которых представляет собой миниатюрную фабрику для производства белков. Многие из этих важнейших макромолекул могут быть выделены из организма в
11. Белки в колесе
11. Белки в колесе Чтобы познакомиться с современными поисками решения проблемы живых часов, обратимся к исследованиям, проводившимся биологами в последние годы. В числе первых следует, пожалуй, назвать работу Патриции де Курси.В 1955 году де Курси получила диплом
Глава 5 Новые гены, новые признаки
Глава 5 Новые гены, новые признаки В предыдущих главах мы уже познакомились с несколькими примерами появления эволюционных новшеств. Внимательный читатель заметил, что новые гены и новые признаки могут возникать разными способами. Пожалуй, нам пора эти способы
СВИДАНИЕ 5. ОБЕЗЬЯНЫ СТАРОГО СВЕТА
СВИДАНИЕ 5. ОБЕЗЬЯНЫ СТАРОГО СВЕТА Поскольку мы достигли этого свидания и готовы приветствовать Копредка 5, нашего прародителя в приблизительно 1.5-милионном поколении, мы пересекаем важную (хотя и несколько произвольную) границу. Впервые в нашем путешествии мы покидаем
Пастер создает новые методы и новые вакцины
Пастер создает новые методы и новые вакцины Бешенство — роковое заболевание, от которого не было спасения. Каждому человеку, укушенному бешеным животным, грозила неминуемая смерть. До получения Пастером вакцины против бешенства из каждых 100 человек, укушенных и
Новые методы, новые вакцины
Новые методы, новые вакцины Поиски методов получения новых живых вакцин проводятся на основе современных научных данных об изменчивости микроорганизмов. Раскрывая тончайшее строение и химический состав микробной клетки и вирусной частички, изучая их физиологию и
Белки мембран.
Белки мембран. Мембранные белки отвечают за функциональную активность мембран и на их долю приходится от 30 до 70%. Белки мембран отличаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его – интегральные белки, разными
Белки
Белки Пищевая ценность белка обеспечивается наличием незаменимых аминокислот, углеводородные скелеты которых не могут синтезироваться в организме человека, и они соответственно должны поступать с пищей. Они также являются основными источниками азота. Суточная
Белки
Белки Белки имеют первостепенное значение в жизни организмов. Огромное разнообразие живых существ в значительной степени определяется различиями в составе имеющихся в их организме белков. Например, в организме человека их известно более 5 млн.Белки – это полимеры,
Научить старого зяблика петь по-новому
Научить старого зяблика петь по-новому Вся эта история об инцесте прекрасно демонстрирует, как врожденные свойства и воспитание вместе работают над формированием нашего характера. Механизм избегания инцеста запускается социально: в детстве у вас развивается
Ну а белки?
Ну а белки? В шестидесятых годах я неоднократно пытался завести в доме и белок, но каждая такая попытка кончалась самым печальным образом. Через некоторое время белки слабели, задние конечности у них отнимались и несчастные животные в судорогах погибали. Поначалу я
Рандеву № 5 Обезьяны Старого Света
Рандеву № 5 Обезьяны Старого Света Приближаясь к рандеву № 5 и готовясь к встрече с сопредком № 5 (нашим прародителем приблизительно в полуторамиллионном поколении), мы пересекаем важный (хотя и произвольно выбранный) рубеж. Впервые с начала путешествия мы вступаем в
Глава 9 Эскизные разработки конструктора природы
Глава 9 Эскизные разработки конструктора природы Представьте себе животное, из тела которого изъято сердце с кровеносной системой, или органы дыхания, или же система пищеварения. А может, не изъято, а при рождении не заложено. К тому же это животное не единичное, а все его