Привередливые дамы способствуют прогрессу
Привередливые дамы способствуют прогрессу
В изучении симпатрического видообразования теория отстала от практики: конкретные случаи удалось зарегистрировать и изучить, но с механизмами, заставляющими единую популяцию разделиться на две репродуктивно изолированные части, пока нет полной ясности. Чтобы симпатрическое видообразование утвердилось в науке в качестве полноправного эволюционного механизма, нужны хорошие математические модели, показывающие, как устроен этот процесс и от каких факторов он зависит.
Многие теоретики предполагали, что без полового отбора и избирательного скрещивания симпатрическое видообразование едва ли возможно. На первый взгляд кажется очевидным, что для симпатрического видообразования представители двух зарождающихся видов должны иметь разные половые предпочтения. Выше мы рассмотрели некоторые идеи, помогающие понять, как у особей в одной популяции могут развиться разные вкусы. Сандер ван Дорн из Бернского университета (Швейцария) и его коллеги из Гронингенского университета (Нидерланды) разработали модель, показывающую, что, даже если все самки в популяции имеют одинаковые половые предпочтения, их привередливость может радикально повысить вероятность симпатрического видообразования (van Doorn et al., 2009).
Размах чувствуется уже в названии статьи. Без ложной скромности авторы озаглавили ее «О происхождении видов путем естественного и полового отбора», что является повторением заглавия перевернувшей мир книги Дарвина, за исключением предпоследнего слова.
Суть модели в следующем. Допустим, существует популяция птиц с клювами средней длины. Местность, где они живут, состоит из участков двух типов. На одних участках преобладают растения с крупными семенами, на других — с мелкими. Для расклевывания крупных семян удобнее иметь длинный клюв, для мелких — короткий. В такой ситуации может начаться дизруптивный отбор. Это значит, что преимущество получат птицы с более длинными или более короткими клювами, но не с промежуточными. Известно, что у реальных птиц, попавших в подобные условия, может произойти симпатрическое видообразование[82]. Остается только понять, как видам удается разделиться, если их представители, перелетая с участка на участок, то и дело встречаются друг с другом и никто не мешает им образовывать смешанные пары и производить гибридное потомство с промежуточными по длине клювами.
В модели все птицы изначально имеют одинаковые клювы средней длины, у самцов нет индикаторов приспособленности, а у самок — избирательности при выборе партнера, поэтому скрещивания происходят случайным образом. Все признаки могут меняться в результате мутаций. Взрослые птицы живут на своих участках, молодые могут мигрировать на другие участки. Специальная переменная задает интенсивность миграций. На участках с крупными семенами лучше себя чувствуют особи с большими клювами, и наоборот. Птица, которая лучше питается, имеет больше шансов дожить до зрелости. Кроме того, ее организм располагает большим количеством ресурсов. Самцы в результате мутаций могут приобрести способность направлять часть этих ресурсов на формирование красного оперения, которое служит индикатором приспособленности. Самки могут (тоже в результате мутаций) приобрести способность выбирать самцов по этому внешнему признаку хорошего здоровья.
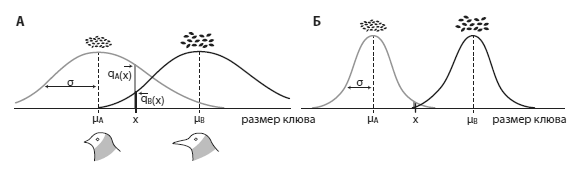
Основные принципы модели ван Дорна. Существуют участки с мелкими и крупными семенами. Для мелких семян наиболее эффективен клюв длиной ?A, для крупных — ?B. По горизонтальной оси — длина клюва, по вертикальной — приспособленность (эффективность питания при таком клюве) на участке с мелкими семенами (серые кривые) и с крупными (черные кривые). А — кривые приспособленности широкие (их ширину отражает величина ?), дизруптивный отбор действует слабо; Б — кривые приспособленности узкие, дизруптивный отбор сильный. Из van Doorn et al., 2009.
И красное оперение, и избирательность самок — «дорогостоящие» признаки. Это значит, что на их развитие тратится часть ресурсов организма, которые могли быть использованы на повышение выживаемости. Самцы платят за свою красоту снижением шансов на выживание. Именно поэтому данный признак может служить индикатором приспособленности. Если бы красное оперение было «бесплатным», все самцы быстро обзавелись бы им, а избирательность самок потеряла смысл.
В модели самцы не могут выработать других украшений, кроме красных перьев на шее. Самки, со своей стороны, не могут выработать других предпочтений, кроме любви к красношеим самцам. В частности, самки не могут научиться выбирать самцов по длине их клюва.
Главными входными параметрами модели являются интенсивность миграций молодых птиц и форма кривой приспособленности (величина ? на рисунке). Сигма отражает степень специализации, необходимой для успешной жизни на данном корме: если сигма очень большая, то с этим кормом справляется любой клюв, если сигма маленькая, то с ним может справиться только клюв определенного размера. Ясно, что эта величина влияет на силу движущего или дизруптивного отбора: чем острее нужда в особом клюве, тем мощнее будет работать отбор, и напротив, отбор будет незначительным, если корм успешно склевывают особи с разными клювами. Вместе сигма и интенсивность миграций задают интенсивность дизруптивного отбора.
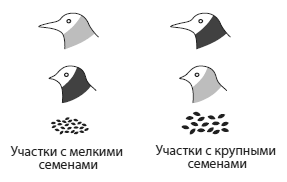
На участках с маленькими семенами лучше себя чувствуют самцы с маленькими клювами, поэтому у них хватает сил отрастить красивые, но дорогостоящие красные перья на шее — индикатор приспособленности. На участках с крупными семенами лучше живется длинноклювым самцам, а короткоклювым украшения оказываются «не по карману». Из van Doorn et al., 2009.
В этой модели у птиц нет возможности выработать презиготическую изоляцию, т. е. перестать скрещиваться с чужаками и производить гибридное потомство. Парадоксальным образом симпатрическое видообразование происходит и без этого. Под успешным видообразованием в данном случае понимается разделение популяции на две четко различающиеся по длине клюва субпопуляции с нулевой или крайне низкой численностью «промежуточных» особей.
Сначала авторы исследовали поведение модели без индикатора приспособленности и половой избирательности. Как и следовало ожидать, в такой ситуации симпатрическое видообразование происходит только при экстремальных значениях параметров: либо при почти полном отсутствии миграций, либо при очень маленькой сигме. В остальных случаях силы дизруптивного отбора хватает лишь на то, чтобы поддерживать в популяции высокий уровень изменчивости по длине клюва, но разорвать популяцию надвое дизруптивному отбору не удается.

Зависимость симпатрического видообразования от полового отбора. По вертикальной оси — интенсивность миграций (0 — птицы никогда не переходят с участка на участок, 1 — полное перемешивание в каждом поколении). По горизонтальной оси — ? (ширина кривой приспособленности, показывающая допустимую вариабельность клювов, успешно справляющихся с данным кормом). Косой штриховкой показана область, в которой симпатрическое видообразование происходит даже без помощи полового отбора. В пределах области, закрашенной точками, симпатрическое видообразование происходит только в том случае, если самцы имеют возможность обзавестись индикатором приспособленности, а самки — половой избирательностью. Сила дизруптивного отбора возрастает по диагонали сверху вниз и справа налево. Серым цветом отмечена область, в которой отбор является не дизруптивным, а стабилизирующим (особи с промежуточной длиной клюва имеют максимальную приспособленность). Из van Doorn et al., 2009.
Затем модельным птицам разрешили приобретать в результате мутаций половую избирательность и способность расходовать часть ресурсов на красные перышки. Эффект оказался неожиданно сильным: теперь исходный вид стал распадаться надвое даже при активной миграции и большой сигме. Таким образом, модель показала, что половой отбор по индикатору приспособленности способен радикально повысить вероятность симпатрического видообразования.
На первый взгляд это кажется странным. Допустим, два вида уже разделились. Но при высоком уровне миграции многие короткоклювые самки будут селиться на участках с большими семенами. Если они выживут, то самцов будут выбирать по красному оперению. На участке с большими семенами красное оперение будет у самцов с большими клювами. Таким образом, короткоклювые самки на «неподходящем» для них участке систематически будут делать «неправильный» выбор и производить потомство с промежуточным размером клюва. Однако в модели частота встречаемости промежуточных клювов после успешного симпатрического видообразования близка к нулю. Как это получается?
Разгадка состоит в том, что приспособленность в этой модели, как и в жизни, — понятие относительное. Выживет ли на данном участке птица с данным размером клюва — зависит не столько от того, хорошо ли ее клюв соответствует семенам на участке, сколько от исхода конкуренции с себе подобными. Если семена большие, а клюв у птицы маленький или средний, это еще не смертный приговор. Если у всех птиц вокруг тоже маленькие или средние клювы, у данной птицы есть хороший шанс выжить. Но если все кругом имеют большие клювы, ее шансы ничтожны. Любая популяция производит больше потомков, чем может прокормиться на данной территории. Выживают не те, кто «вообще» хорошо приспособлен к данной среде, а те, кто приспособлен хорошо по сравнению с остальными.
Таким образом, в исходной ситуации, когда у всех птиц средние по размеру клювы, на любом из участков можно нормально жить, имея средний клюв. Но, как только популяция под действием дизруптивного отбора начинает распадаться на две, для среднеклювых птиц наступают тяжелые времена. На обоих типах участков их безжалостно вытесняют специалисты — длинноклювые и корткоклювые конкуренты. Половой отбор действует просто-напросто как усилитель дизруптивного отбора. Когда же популяции начинают расходиться, дизруптивный отбор усиливается дополнительно еще и из-за самого факта расхождения. Чем дальше оно зашло, тем эффективнее отбраковка промежуточных особей. В итоге отбор начинает убирать среднеклювых птиц из популяции быстрее, чем они нарождаются.
Разумеется, рассмотренный в модели механизм — не единственный возможный. Симпатрическое видообразование станет еще более вероятным, если разрешить птицам в ходе эволюции менять алгоритм миграций. Например, птицы могут выработать алгоритм «мигрируй, только если вокруг нет удобных для тебя семян» или «селись только там, где много удобных семян». Это резко снизит частоту гибридизации. Еще одна возможность связана с развитием механизма выбора партнера «с оглядкой на себя» или с использованием импринтинга (запечатления). Например, самка может предпочитать партнеров с таким же клювом, как у нее самой или у ее отца.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Привередливые мужчины
Привередливые мужчины Возникает парадокс. Мужчины в глубине души и в своих фантазиях — сексуальные оппортунисты. Казалось бы, таковые не должны быть особо привередливы. Тем не менее мужчине важно, как выглядит женщина — важнее, чем ей, как выглядит он.Спортивная машина
Ценностно-нейтральный и ценностно-нагруженный подход к прогрессу
Ценностно-нейтральный и ценностно-нагруженный подход к прогрессу Прогрессивна ли эволюция? Есть по крайней мере одно разумное определение прогресса, при котором я готов его защищать. Начнем с того, что понятие прогресса можно применять в нестрогом минималистском