Вперед, в воздушный космос
Вперед, в воздушный космос
Рассмотрим, как из рыб получились четвероногие животные, ставшие завоевателями суши: как рыбы отправились в наземный космос. Это событие датируется концом девонского периода, 385–360 млн лет назад. Первые четвероногие были не первыми существами, покинувшими водную среду: на суше в конце ордовика уже поселились растения, по-видимому, еще в кембрии в почвах появилась богатая фауна беспозвоночных, а еще раньше обитателями суши стали бактерии и грибы. Так что рыбы, пожертвовав гармонией со своей привычной водной средой и превратившись в медлительных, неповоротливых четвероногих, пришли в населенный и гостеприимный мир. Если в воде их подстерегали хищники, а за пищу приходилось конкурировать со многими голодными едоками, то на суше крупных хищников не было и привлекало обилие доступной пищи. Поэтому рискованный и трудный выход на сушу был выгодным предприятием. Это была гигантская незанятая экологическая ниша. И эволюция поспешила заполнить ее, как только представилась возможность.
Возможность, по-видимому, обеспечили условия девонского периода. Палеоэкологическая интерпретация местонахождениий с девонскими переходными формами рисует нам мелкие, слабопроточные пресные водоемы субтропического или тропического пояса. Предполагается пониженное содержание кислорода в атмосфере и в воде. Возможно, из-за сильных периодических обмелений позднедевонским лопастеперым рыбам пришлось приспосабливаться к жизни в условиях, где вода больше не служит опорой для тела, потому что слишком мелко, где приходится дышать без помощи жабр атмосферным воздухом.

Прототакситы (Prototaxites), конусовидные сооружения девонского периода высотой от 6 до 9 м. Как выяснилось, это гигантские вымершие грибы. Богатая грибная флора существовала гораздо раньше, не меньше миллиарда лет назад. Об этом свидетельствуют, в частности, исследования Константина Наговицына из Института нефтегазовой геологии и геофизики, выделившего разнообразные и необычные ископаемые грибы из протерозойских пород возрастом 1 млрд лет. Нужно помнить, что высшие растения могли начать освоение суши только «с ведома» своих непременных спутников — грибов и бактерий. Кто же еще, если не они, станет фиксировать для растений атмосферный азот и высвобождать необходимые растениям вещества из захороненной в почве органики? Ведь без этого невозможна жизнь наземного растения. Освоение суши растениями было подготовлено предшествующим заселением суши грибами и бактериями.
Эти водоемы облюбовали различные рыбы — и их было много, включая крупных и мелких хищников. Не случайно в девонских морях 17 % родов рыб приобрели панцирь — мирные обитатели обзаводились средствами защиты. Это было время расцвета рыб — помимо бесчелюстных, появившихся не позднее ордовика, в девонских морях плавали хрящевые и костные рыбы, разделившиеся еще в силуре. А костные рыбы в девоне были уже представлены двумя группами — лучеперыми и лопастеперыми. Эти группы различаются, помимо прочего, строением плавников: у лопастеперых плавник сидит на мясистом основании, скелет которого составляют удлиненные кости, плавниковая лопасть поддерживается серией симметричных костных пластин. У лучеперых плавниковые лопасти асимметричны, а кости основания плавника укорочены.
Именно одна из групп лопастеперых с мощными грудными и брюшными плавниками, напоминающими лапы — рипидистии, — дала начало наземным позвоночным. К лопастеперым относятся современные кистеперые (латимерия) и двоякодышащие (рогозуб, протоптер, чешуйчатник). Генетический анализ показал, что из современных рыб ближайшими родственниками тетрапод являются двоякодышащие.
Эволюционные изменения, происходившие с рипидистиями — предками первых тетрапод (земноводных), были постепенными. Из позднедевонских отложений известно более десятка представителей, сочетавших признаки земноводных и рыб. Они выстраиваются в относительно стройный ряд ископаемых форм: эустеноптерон (Eusthenopteron), пандерихт (Panderichthys), тиктаалик (Tiktaalik), элпистостега (Elpistostega), ливониана (Livoniana), эльгинерпетон (Elginerpeton), вентастега (Ventastega), метаксигнат (Metaxygnathus), акантостега (Acanthostega), ихтиостега (Ichthyostega), тулерпетон (Tulerpeton) и грирерпетон (Greererpeton). В этом ряду постепенно убывают рыбьи и накапливаются тетраподные черты, но разные системы органов совершают этот переход не одновременно, какие-то приходят к тетраподному состоянию быстрее, какие-то медленнее. Вслед за знаменитым шведским палеозоологом Эриком Ярвиком этих созданий часто называют «четвероногими рыбами».
Основное, что потребовалось изменить, чтобы получить право называться «настоящими четвероногими», — это конечности. Впрочем, четвероногость и хождение как таковое тоже возникли задолго до самих четвероногих. Ловко ходить по дну на четырех плавниках, приподнимая тело над грунтом, умели многие лопастеперые рыбы. Так что преадаптации для хождения имелись еще на «рыбьей» стадии. Но, чтобы ходить эффективно по суше, где тело весит гораздо больше, желательно все же иметь специализированную конечность. Нужно было сформировать подвижный сустав лапы и открепить пояс передних конечностей от черепа. Ведь у рыб пояс грудных плавников жестко крепится к задневисочной кости черепа, а это ограничивает движения и головы, и плавников.
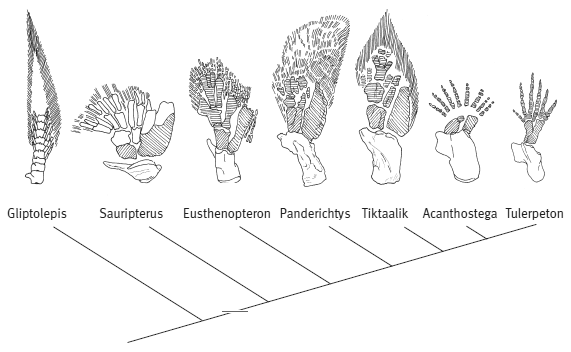
Схема изменения скелета передних конечностей у «четвероногих рыб».
Но если присмотреться повнимательнее, то окажется, что конечности (плавники) девонских лопастеперых рыб не так уж сильно отличались по своему плану строения от лап примитивных тетрапод. Хотя конечности сохранились не у всех ископаемых переходных форм, мы достаточно подробно знаем, что и как изменялось. Уже у эустеноптерона в переднем плавнике имелась кость, соответствующая плечевой кости тетрапод, и две кости, соответствующие локтевой и лучевой костям. Строение «дистальных элементов», соответствующих будущей кисти тетрапод, у эустеноптерона и пандерихта еще оставалось относительно неупорядоченным.
Далее в этом ряду поместился тиктаалик, обнаруженный в 2006 году американскими палеонтологами Эдвардом Дешлером, Нилом Шубиным и Фаришем Дженкинсом на острове Элсмир (арктическая Канада). «Тиктаалик» на языке эскимосов означает «большая пресноводная рыба, живущая на мелководье». И вправду, тиктаалик — это плоская, покрытая крупной чешуей рыба с крокодильей головой, на которой сверху сидят глаза, впереди две ноздри и большая зубастая пасть. У этой рыбы, как и у других представителей интересующего нас ряда переходных форм, часть черт сходна с лопастеперыми, а другие признаки сближают ее с четвероногими. Рыбьи признаки — это чешуя, плавниковые лучи, почти такие же, как у кистеперых, сложная нижняя челюсть и небные кости. Тетраподные признаки — укороченный череп, отделенная от пояса передних конечностей и потому относительно мобильная голова, наличие локтевого и плечевого суставов (Daeschler et al., 2006).
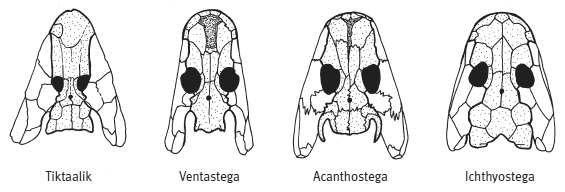
Черепа «четвероногих рыб» и первых тетрапод. Из статьи Ahlberg et al., 2008. Nature. V. 453. P. 1199–1204.
По сравнению с пандерихтом у тиктаалика скелет передних конечностей приобрел чуть более оформленный вид, так что становится уже более-менее понятно, откуда взялись у потомков тиктаалика косточки пясти и пальцев. Тиктаалик и пандерихт уже могли сгибать свой передний то ли плавник, то ли лапу в суставе, который у их потомков назовут локтевым. А еще тиктаалик опробовал одно небольшое, но важное нововведение — он почти избавился от жаберной крышки, которая еще была у пандерихта (к функциям жаберной крышки мы еще вернемся).
Вместе с жаберной крышкой потерялась и жесткая связь между поясом передних конечностей и черепом. Голова освободилась от передних конечностей (или конечности от головы). То и другое стало более подвижным. Теперь можно было начинать учиться нормально ходить. У тиктаалика ребра сплюснулись, соединения позвонков окостенели. За счет этого он стал хуже сгибаться, зато тело приобрело устойчивость, что очень важно для ходячего, а не плавающего образа жизни.
Утрата жаберной крышки оказалась на редкость полезна в дальнейшем. Сочленение остатков жаберной крышки с головой вроде бы стало рудиментарным довеском для наземного, дышащего легкими существа. Но оно не исчезло вовсе, а начало медленное и крайне важное для всех наземных четвероногих путешествие внутрь черепа, превращаясь потихоньку в крошечные слуховые косточки. В действительности слуховые косточки начали формироваться еще до утери жаберной крышки. Как было показано Пером Альбергом и Мартином Бразо из Уппсалы, пандерихт, а не тиктаалик первым приспособил под примитивное стремечко (слуховую косточку) одну из костей сочленения жаберного аппарата с черепом (гиомандибулу). Эта кость была у него тонкой, изящной, она прилегала к дыхательному отверстию (первой жаберной щели), которое расширилось у пандерихта. У рыб это отверстие называется брызгальцем, оно имеется у эмбрионов тетрапод, но по ходу развития эмбриона становится полостью среднего уха и евстахиевой трубой. Пандерихт дышал воздухом, насосный аппарат жаберной крышки (см. ниже) ослабился, и части его переориентировались на выполнение другой функции — восприятие звука. От рыб, близких к пандерихту, стремечко унаследовали все тетраподы, включая нас (Brazeau, Ahlberg, 2006).
У вентастеги, которая по строению черепа занимает промежуточное положение между тиктааликом и акантостегой, от конечностей, к сожалению, почти ничего не сохранилось. Широкая форма рыла, строение черепной коробки сближают вентастегу с акантостегой, а форма и пропорции покровных костей черепа — с тиктааликом.
Широкая челюсть вентастеги была усажена мелкими острыми зубами. На длинном теле — около метра-полутора в длину — имелось две пары коротеньких конечностей с пальцами (сколько пальцев — неизвестно) и хвост с плавником, который поддерживали плавниковые лучи длиной около 7 см. Вентастега жила в солоноватоводных мелких прибрежьях и, имея внушительные размеры, охотилась на рыб. Как и у пандерихта с тиктааликом, у вентастеги часть бывшего сочленения жаберного аппарата с черепом была преобразована в слуховую косточку — стремечко. Куда относить вентастегу — еще к рыбам или уже к тетраподам, т. е. древнейшим амфибиям, — вопрос формальный, но если у нее действительно были пальцы, то, пожалуй, второй вариант предпочтительнее.
Следующие в ряду — акантостега и ихтиостега, которых принято считать уже «настоящими» тетраподами. Их конечности представляют собой всего лишь еще один небольшой шажок в сторону дифференциации дистальных частей скелета (стопы и кисти). У них окончательно оформились три крупные кости конечности (в передней лапе это плечо, локтевая и лучевая кости), тогда как многочисленные косточки кисти пока еще были мелкими и изменчивыми. Но у акантостеги уже точно были настоящие пальцы, а плавниковых лучей, которые еще сохранялись у тиктаалика, у нее не было. При этом число пальцев у акантостеги варьировало от пяти до восьми: признак еще не «стабилизировался». Изменчив он и у ихтиостеги. Более стабильное число пальцев характерно для тулерпетона (шесть пальцев); также у него имелись уже совсем «тетраподные» локтевая, лучевая и пястные косточки. У грирерпетона число пальцев стабилизировалось — теперь их будет всегда пять.
Кроме конечностей выход на сушу потребовал глубокого преобразования дыхательной и кровеносной системы. Опять же кажется, что переход от дыхания водой к дыханию воздухом невозможен мелкими улучшениями конструкции — нужно какое-то масштабное преобразование сразу обеих систем. Но это, как выясняется, не так. Подобно постепенному преобразованию конечностей так же постепенно происходил и переход от жаберного дыхания к легочному. И это можно увидеть на том же ряду ископаемых четвероногих рыб.
Итак, нужно сформировать легкие и, соответственно, два круга кровообращения и трехкамерное сердце. Такое строение кровеносной и дыхательной систем, судя по палеонтологическим данным, было чуть ли не у всех девонских лопастеперых рыб. Сохранилось оно и у современных двоякодышащих рыб. В качестве органа воздушного дыхания лопастеперые стали использовать особое впячивание передней части пищевода. Рыба заглатывает воздух, который отправляется в этот пузырь, оплетенный кровеносными сосудами, — это наипростейшее легкое.
У двоякодышащих пузырь выполняет две функции: примитивного легкого и примитивного гидростатического органа. У большинства (но не у всех) костных рыб раздувшееся выпячивание пищевода специализировалось для выполнения второй функции и превратилось в «настоящий» плавательный пузырь. Но у девонских рыб этот вырост пищевода совмещал функции легкого и регулятора плавучести.
В девонских водоемах обретение дополнительного органа дыхания было более чем оправдано: содержание кислорода в атмосфере было понижено, а доля углекислого газа, наоборот, повышена по сравнению с современностью. Поэтому в воде, особенно в стоячей, рыбам было трудно дышать одними жабрами. И они приобрели дополнительный орган для дыхания атмосферным воздухом. Так что для выхода на сушу уже велась «подготовка» исподволь — легкие для дыхания воздухом и прилагающаяся к ним модифицированная система кровообращения уже были припасены у девонских лопастеперых рыб. Разумеется, отбор поддерживал их как адаптации для жизни в бедной кислородом воде. О том, для чего они потом пригодятся, ни рыбы, ни отбор знать не знали.
Появление дыхательного пузыря — примитивного легкого — создало предпосылки для еще одного новшества: эффективного воздушного насоса. У костных рыб воду через жабры гонит жаберная крышка: рыба закрывает рот и поднимает среднюю часть крышки, увеличивая объем жаберной полости (ее мягкая оторочка плотно прижата к телу). Соответственно, давление в жаберной полости уменьшается и, когда рыба открывает рот, туда устремляется вода. Затем рыба закрывает рот, жаберная крышка опускается, оторочка отходит от края и вода вытекает наружу, омывая жабры.
Все это возможно в плотной водной среде, когда требуется обеспечить сквозной ток воды через жабры. В разреженной воздушной среде такой насос будет крайне неэффективным, ведь помимо разной плотности среды у дыхательного насоса и задачи другие. Нужно не гнать воздух сквозным потоком, а вентилировать слепой воздушный мешок. Но эту задачу первые тетраподы (земноводные) толком так и не решили. Это удалось лишь рептилиям. А у земноводных принцип нагнетания воздуха в легкие примерно тот же, что и у дышащих воздухом рыб. Они используют преобразованный аппарат жаберных дуг для расширения глоточной полости и, соответственно, для вдоха. Отработанный воздух из легких выдыхается за счет сокращения мускулатуры легких (а у двоякодышащих рыб он выходит за счет более высокого давления в воде по сравнению с воздушной поверхностью над водой, куда рыба выставляет голову для вдоха-выдоха).
Особенности жаберного аппарата, приспособленного для вдоха-выдоха за счет движения глотки, а не жаберной полости, хорошо отражаются в скелете и современных, и ископаемых девонских лопастеперых. Для более эффективного выведения углекислого газа из организма лопастеперые используют жабры, а земноводные, утратившие жабры, используют кожу («кожное дыхание»). Это создает массу ограничений, из-за которых земноводные так и не смогли далеко уйти от воды.
Но в воздушной среде жаберная крышка все равно не работала в качестве воздушного насоса, поэтому от нее пришлось отказаться, и это открыло наземным обитателям дополнительные возможности — косточки жаберной крышки ушли внутрь черепа. И начали формировать среднее ухо. Жаберная крышка частично редуцировалсь у тиктаалика. Это означает, что тот способ дыхания, которым продолжали пользоваться другие рыбы, стал ему больше не нужен. Тиктаалик дышал в основном воздухом при помощи легких и, возможно, кожей.
Раз у тиктаалика редуцировалась жаберная крышка, это означает, что ему пришлось научиться глотать без использования всасывающего жаберного насоса — рыбы втягивают частицы пищи с током воды. Тиктаалик глотал только за счет движения глотки и головы. У акантостеги, наследницы тиктаалика, еще сохранялись внутренние жабры, а у ихтиостеги они уже редуцировались (наружные жабры у личинок амфибий сохраняются и сегодня). Следовательно, последователи тиктаалика тоже глотали за счет движений головы и челюстей. В дальнейшем по мере редукции остатков жаберного аппарата у древних амфибий происходило постепенное развитие шейного отдела.
Таким образом, в строении дыхательной и кровеносной систем при переходе от рыб к тетраподам не было резких скачков, а большинство важных изменений произошло еще на «рыбьем» этапе. Каждое новшество в дыхании и кровообращении не создавалось на пустом месте, а лишь улучшало требуемые функции на основе развития имеющихся приспособлений.
—————
Плавники превращаются в пальцы: взгляд генетика
Недавно было обнаружено семейство генов, необходимых для развития плавников у рыб и получивших название actinodin (and). У наземных позвоночных этих генов нет. Если отключить у рыбы гены and, то, во-первых, в парных плавниках не развиваются эластичные плавниковые лучи — лепидотрихии (но в хвостовом плавнике они развиваются нормально), во-вторых — меняется характер работы ряда ключевых генов-регуляторов (в том числе hoxd13a и sonic hedgehog), которые есть не только у рыб, но и у четвероногих и от которых зависит передне-задняя полярность зачатка конечности (будь то плавник или нога). Похожие изменения в работе этих генов-регуляторов у четвероногих, возникающие в результате некоторых мутаций, приводят к полидактилии (многопалости).
По-видимому, потеря генов and древними четвероногими привела к утрате лепидотрихий в парных конечностях, однако в хвостовом плавнике лепидотрихии могли сохраниться. Именно так обстоит дело у древнейших четвероногих, обнаруженных в ископаемой летописи, таких как акантостега и ихтиостега. У этих животных уже не было плавниковых лучей на парных конечностях, но они еще сохранялись в плавниковой складке, окаймлявшей хвост. У более ранних переходных форм, таких как тиктаалик, плавниковые лучи на парных конечностях имелись.
Логично предположить, что утрата генов and и вызванная ею дестабилизация работы регуляторных генов у древнейших четвероногих создала предпосылки для развития у них большого числа (до восьми) почти одинаковых пальцев. В дальнейшем у четвероногих восстановился исходный, «нормальный» характер работы генов-регуляторов, причем теперь их работа уже не зависела от утраченных генов and. Результатом этого могло стать автоматическое сокращение числа пальцев до пяти. Так изучение генетики современных животных помогает понять молекулярные механизмы событий, произошедших сотни миллионов лет назад (Zhang et al., 2010).
—————
Мы рассмотрели ряд переходных форм, связывающих лопастеперых рыб с примитивными четвероногими. Совершенно непонятно, где в этом ряду находится искомый великий «макропереход». Приобретение выростов пищевода, в которые можно набирать воздух? Нет, это просто адаптация рыб к плохо аэрируемым водоемам. Аналогичные органы воздушного дыхания (например, в виде модификаций ротовой полости) возникали и в более поздние времена у разных рыб, оказавшихся в схожих условиях. Приобретение костей поясов конечностей у эустеноптерона? Едва ли, ведь это тоже была чисто «рыбья» адаптация к большей подвижности плавников. Может, назначить этот рубеж по приобретению подвижного сустава у лап, т. е. где-то между эустеноптероном и тиктааликом? Но ведь это просто адаптация рыб к ползанию по дну мелких луж и переползанию из одного пересохшего водоема в другой, развивавшаяся к тому же долго и постепенно. Еще есть вариант — присвоить «макро»-титул тиктаалику, у которого передние конечности отделились от черепа и стали свободными лапками… хотя и это всего лишь улучшенный вариант адаптации для передвижения по дну на мелководье. Утрата плавниковых лучей и приобретение пальцев? При большом желании можно, конечно, объявить эту небольшую модификацию дистальных участков конечностей тем самым эпохальным «превращением рыбы в амфибию». Действительно, именно в этом месте непрерывного ряда переходных форм специалисты проводят формальную грань между рипидистиями и тетраподами. Но не лучше ли честно признать, что палеонтологические данные показывают просто длинный ряд постепенных маленьких изменений, каждое из которых само по себе никак не тянет на «макрособытие»?
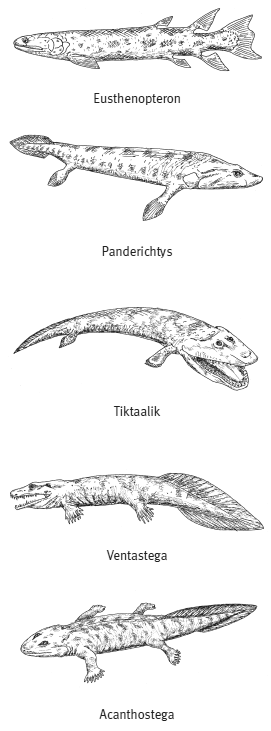
Реконструкции внешнего облика некоторых позднедевонских «четвероногих рыб». Сверху вниз: эустеноптерон, пандерихт, тиктаалик, вентастега, акантостега.
Всюду в этом ряду мы видим последовательные адаптации, служившие их обладателям верой и правдой в конкретных условиях. Ни одна из них по отдельности не заслуживает статуса «макроизменения». Сопоставимые по масштабу эволюционные преобразования происходят и в современной природе вокруг нас, и в лабораторных экспериментах, и в повседневной работе селекционеров. И уж конечно, если бы первые тетраподы вымерли, так и не дав начало большой и разнообразной группе животных, разумные осьминоги ни за что не выделили бы ихтиостегу, акантостегу и их родню в особый класс — считали бы их самое большее отдельным тупиковым семейством лопастеперых. И были бы правы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Индуцированные мутации. Дрозофила летит в космос
Индуцированные мутации. Дрозофила летит в космос Получить искусственные мутации, изменить своими руками наследственность — об этом генетики мечтали данно. Но не так-то легко было подобрать ключик! Наследственность всего живого надежно защищена от внешних воздействий.
Иммунология и космос
Иммунология и космос Иммунология и космос — одна из самых современных связей иммунологии.Как видите, все новые и новые связи. Надо сказать, что мы не можем упрекнуть в этом нашу иммунологию. Все эти союзы и сочетания очень многое дали и сугубо теоретической биологии,
Глава 25 Микробиология и космос
Глава 25 Микробиология и космос Человечество не останется на Земле, но в погоне за светом и пространством… завоюет все околосолнечное пространство. К. Э. Циолковский Исследование и освоение космического пространства связано с развитием новых научных направлений, к
КОЛБЫ И КОСМОС
КОЛБЫ И КОСМОС У биологов прошлого века не было нашей техники, но не было и наших проблем. Не было и проблемы происхождения жизни — биогенеза. Все казалось ясным: либо творение, либо самозарождение. Творение находилось за пределами науки, а самозарождение было очевидным.
Вперед на восток!
Вперед на восток! Основная масса людей эпохи верхнего палеолита начала распространяться в восточном направлении. Как и в случае других древних человеческих миграций, эта почти наверняка не была сознательным перемещением из одного места в другое. Скорее всего, ей
История Земли и жизни — окно в космос
История Земли и жизни — окно в космос На пороге космической эры, в эпоху бурного развития всех отраслей наук, история Земли и жизни приобретает особо важное значение. От изучения поверхности земного шара исследователи переходят к происхождению, эволюции и дальнейшей
ЕСТЕСТВЕННЫЙ ОТБОР: НЕ ВСЕ В ЭВОЛЮЦИИ ДВИЖЕТСЯ ВПЕРЕД
ЕСТЕСТВЕННЫЙ ОТБОР: НЕ ВСЕ В ЭВОЛЮЦИИ ДВИЖЕТСЯ ВПЕРЕД Шок Дарвина Леонардо да Винчи (1452–1519) говорил: «В природе нет ошибок, но знай: ошибка есть в тебе». Слишком совершенным казалось этому гению все богатство и разнообразие форм в природе, чтобы хоть немного усомниться в
Вперед, к современности
Вперед, к современности Время шло, и наши предки оставляли нам все больше свидетельств своих любовных перипетий. Полмиллиона лет назад объем мозга людей, обитавших на территории современной Эфиопии, уже достигал 1300 куб. см, то есть был равен по размеру мозгу современного
Глава V Космос и океан
Глава V Космос и океан И вот уже в космическом пространстве Друг друга окликают корабли. Я думаю о гордом постоянстве, О мужестве сынов моей Земли! Николай
«Два шага вперед, затем шаг назад…»
«Два шага вперед, затем шаг назад…» К концу 1980–х годов корпорации–производители трансгенных семян, благодаря новому влиянию ВТО и при полной поддержке Белого дома, стали явно увлекаться возможностью захвата контроля над мировыми продовольственными ресурсами. Они все
ПРЫЖОК В КОСМОС
ПРЫЖОК В КОСМОС Писательница и путешественница Елена Блаватская рассказывала о книге «Станцы Цзяна». Когда она захотела предъявить саму книгу мировой общественности, «Станцы» загадочным образом исчезли из новейшего сейфа в громадном отеле… «Вероятно, это дело рук
Путь вперед
Путь вперед Закончив первое фМРТ-исследование, я вернулся к своим энцефалограммам и интервью с новыми заключен ными. В свободное время я написал статьи о пяти ис следованиях для диссертации и подачи в рецензируемые журналы.Моя диссертация близилась к завершению,
Глава 13 Вперед, и только вперед
Глава 13 Вперед, и только вперед Фундаментальные аспекты наследственности оказались, к нашему удивлению, довольно просты, а потому появилась надежда, что, возможно, природа не так уж непознаваема, а ее не раз провозглашаемая самыми разными людьми непостижимость –