КЕНТЕРБЕРИ
КЕНТЕРБЕРИ
Как и подобает конечной цели 4-миллиардолетнего путешествия, наше Кентербери обладает налетом тайны.
Это особенность, известная как происхождение жизни, но мы могли лучше назвать ее происхождением наследственности. Сама жизнь явно не определена, факт, который противоречит интуиции и традиционной мудрости. Иезекииль, Глава 37, в которой пророку повелели спуститься в долину костей, отождествляет жизнь с дыханием. Я не могу удержаться от цитирования отрывка («кость с костью своею» – столь замечательная структура языка).
Я изрек пророчество, как повелено было мне; и когда я пророчествовал, произошел шум, и вот движение, и стали сближаться кости, кость с костью своею. И видел я: и вот, жилы были на них, и плоть выросла, и кожа покрыла их сверху, а духа не было в них.
Тогда сказал Он мне: изреки пророчество духу, изреки пророчество, сын человеческий, и скажи духу: так говорит Господь Бог: от четырех ветров приди, дух, и дохни на этих убитых, и они оживут.
И, конечно, дух вошел в них. Великое полчище ожило и стало на ноги. Дыхание для Иезекииля определяет различие между мертвым и живым. Сам Дарвин подразумевал то же самое в одном из своих более красноречивых пассажей, заключительных словах «Происхождения видов» (курсив добавлен):
Таким образом, из борьбы в природе, из голода и смерти непосредственно вытекает самый высокий результат, какой ум в состоянии себе представить, – образование высших животных. Есть величие в этом воззрении, по которому жизнь в ее различных проявлениях Творец первоначально ВДОХНУЛ в одну или ограниченное число форм; и между тем как наша планета продолжает вращаться согласно неизменным законам тяготения, из такого простого начала развилось и продолжает развиваться бесконечное число самых прекрасных и самых изумительных форм.
Дарвин справедливо полностью изменил порядок событий Иезекииля. Дыхание жизни было первым и создало условия, при которых, в конечном счете, развились кости и сухожилия, плоть и кожа. Кстати, выражение «Творец» не присутствует в первом издании «Происхождения видов». Оно было добавлено во втором издании, вероятно как подачка религиозному лобби. Дарвин позже сожалел об этом в письме к своему другу, Хукеру:
Но я уже давно сожалею, что уступил общественному мнению и употребил выражение Пятикнижия – "сотворение", под которым я на самом деле только подразумеваю "появление" вследствие какого-то совершенно неизвестного нам процесса. Сущий вздор – рассуждать сейчас о происхождении жизни; с тем же успехом можно было бы рассуждать о происхождении материи.
Дарвин, вероятно, (и, на мой взгляд, справедливо) видел происхождение примитивной жизни как относительно (и я подчеркиваю относительно) легкую проблему по сравнению с той, которую он решил: как жизнь, когда-то возникшая, развивала свое удивительное разнообразие, сложность и сильную иллюзию разумного замысла. Однако позже (в другом письме Хукеру) Дарвин высказывает предположение о «совершенно неизвестном процессе», который начал все это. Он пришел к этому, размышляя, почему мы не наблюдаем возникновение жизни снова и снова.
Часто говорят, что сейчас есть все условия для создания первого живого организма, которые могли быть когда-либо предоставленными. Но если (и о! какое большое если!) бы сейчас в каком-нибудь маленьком теплом пруду со всеми видами аммония, солей фосфора, светом, теплом, электричеством и т. д., химически образовался белок, способный к дальнейшим все более сложным превращениям, то это вещество немедленно было бы разрушено и поглощено, что было невозможно в период возникновения живых существ.
Доктрина самозарождения лишь недавно была опровергнута экспериментами Пастера. Долгое время считалось, что в гниющем мясе самопроизвольно образовывались личинки, что морские уточки спонтанно порождали гусят и даже, что в грязном белье, помещенном вместе с пшеницей, самозарождались мыши. Странно, но теория самозарождения была поддержана Церковью (которая следовала в этом и многом другом за Аристотелем). Я говорю странно, потому что, по крайней мере, с ретроспективной точки зрения на прошлое, самозарождение было столь же прямым вызовом божественному созданию, какой всегда была эволюция. Идея, что мухи или мыши могли появиться самопроизвольно, весьма недооценивает громадное достижение, которым было бы создание мух или мышей: оскорбление Творца, как можно было бы подумать. Но ненаучное мышление не в состоянии понять, как сложны и, в сущности, невероятны муха или мышь. Дарвин был, возможно, первым, кто оценил всю величину этой ошибки.
Уже в 1872 году в письме Уоллесу, cооткрывателю естественного отбора, Дарвин нашел необходимым выразить свой скептицизм относительно «самозарождения коловраток и тихоходок», как говорилось в книге «Источники жизни», которой он в остальном восхищался. Его скептицизм, как обычно, попал в точку. Коловратки и тихоходки – сложные жизненные формы, превосходно приспособленные к своим соответствующим образам жизни. Для них самозарождение означало бы, что они стали приспособленными и сложными «благодаря счастливому стечению обстоятельств, а в это я не могу поверить». Счастливые стечения обстоятельств таких размеров были ненавистны Дарвину, поскольку они, по различным причинам, должны были иметь отношение к Церкви. Общим принципом теории Дарвина было и есть то, что адаптивная сложность возникает как медленные и постепенные изменения, шаг за шагом, а не единственным шагом, предъявляя слишком большие требования к слепому случаю для объяснения. Дарвиновская теория, разделив случайность на маленькие шаги, должна была дать вариации для отбора, представив единственный реалистичный результат чистой случайности как объяснение жизни. Если бы коловратки могли возникнуть подобным образом, работа всей жизни Дарвина была бы ненужной.
Но у самого естественного отбора должно было быть начало. В этом смысле одно какое-то самозарождение должно было случиться, хотя бы только однажды. Красота работы Дарвина состояла в том, что в единственном самозарождении, которое мы обязаны постулировать, не должно было синтезироваться ничего сложного, вроде личинки или мыши. Должно было быть лишь создано... что ж, сейчас мы приближаемся к сути проблемы. Если не дыхание, какой компонент жизни позволил естественному отбору начаться и привести, в конечном счете, после эпопеи кумулятивной эволюции, к личинкам, мышам и человеку?
Детали скрыты, возможно, безвозвратно, в нашем древнем Кентербери, но мы можем дать ключевому элементу минималистическое название, чтобы выразить то, чем он должен был быть. Это название – наследственность. Мы должны искать не происхождение жизни, которая туманна и неопределенна, а происхождение наследственности – истинной наследственности, и это означает нечто весьма точное. Ранее для объяснения этого я использовал огонь.
Огонь соперничает с дыханием за образное представление жизни. Когда мы умираем, жизненный огонь гаснет. Наши предки, которые впервые его приручили, вероятно, думали, что огонь – живое существо, даже бог. Вглядываясь в огонь или тлеющие угольки, особенно ночью, когда их грел и защищал походный костер, общались ли они в своем воображении с пылающей, танцующей душой? Огонь живет, пока Вы его подпитываете. Огонь дышит воздухом; Вы можете задушить его, прекратив подачу кислорода, Вы можете утопить его в воде. Лесной пожар пожирает лес, гоня живую добычу перед собой со скоростью и жестокостью стаи волков в (буквально) горячем преследовании. Как и в случае с волками, наши предки могли захватить детёныша огня в качестве полезного домашнего животного, приручить его, регулярно кормить и убирать его пепельные выделения. Прежде, чем было открыто искусство добывания огня, общество должно было высоко ценить меньшее искусство разумного использования захваченного огня. Возможно, живой побег домашнего костра несли в горшке для обмена с соседней группой, у которой, к сожалению, умер собственный огонь.
Было бы сделано наблюдение, что лесной пожар порождает дочерние костры, плюясь искрами и развеивая пепел по ветру, как пух одуванчика, чтобы достичь и засеять семенами сухую траву на некотором расстоянии. Могли ли философы Homo ergaster теоретизировать, что огонь не может самозародиться, а должен всегда происходить от родительского огня: либо лесного пожара, потухшего на равнине, либо домашнего костра, огороженного каменными плитами в очаге? И первые палочки для добывания огня, поэтому, уничтожают эту точку зрения?
Наши предки, возможно, даже представляли себе популяцию размножающихся лесных пожаров или родословную домашних костров, прослеженную от пылающего предка, купленного у далекого клана и используемого другими. Но это все еще не была истинная наследственность. Почему? Как Вы можете иметь размножение и родословную, но все же не иметь наследственности? Это – урок, полученный нами здесь от огня.
Истинная наследственность означала бы наследование не самого костра, а вариаций среди костров. Некоторые более желтые, чем другие, некоторые более красные. Одни ревут, другие потрескивают, какие-то шипят, дымят или сыплют искрами. У некоторых в пламени есть оттенок синего или зеленого. Наши предки, если бы они изучили своих одомашненных волков, заметили бы сильное различие между родословными собак и родословными костров. У собак подобное порождает подобное. По крайней мере, что-нибудь из того, что отличает одну собаку от другой, передано ее родителями. Конечно, кое-что также приходит со стороны: с пищей, болезнями и несчастными случаями. У огня все изменения происходят из окружающей среды, ни одно не передается по наследству от предковой искры. Они зависят от качества и сырости топлива, от направления и силы ветра, от свойства тяги очага, от почвы, от небольшого количества меди и калия, которые добавляют сине-зеленый и сиреневый оттенок к желтому пламени натрия. В отличие от собак, ни одно из качеств взрослого костра не происходит от искры, которая его породила. Синий огонь не порождает синий огонь. Потрескивающие костры не наследуют свое потрескивание от родительского костра, подбросившего свою первоначальную искру. Огни демонстрируют воспроизводство без наследственности.
Происхождение жизни было происхождением истинной наследственности; можно даже сказать, происхождением первого гена. Под первым геном, поспешу заметить, я не подразумеваю первую молекулу ДНК. Никто не знает, был ли первый ген сделан из ДНК, и я держу пари, что не из нее. Под первым геном я подразумеваю первый репликатор. Репликатором является структура, например молекула, которая создает линию своих копий. В копировании всегда будут ошибки, таким образом, популяция приобретет разнообразие. Ключ к истинной наследственности в том, что каждый репликатор больше похож на того, с которого он был скопирован, чем на случайного представителя популяции. Происхождение первого такого репликатора было невероятным событием, но это должно было случиться только однажды. С того времени его последствия поддерживали себя автоматически, и в конечном счете дали толчок, путем дарвиновской эволюции, к развитию всей жизни.
Участок ДНК или, при определенных условиях, соответствующей молекулы РНК – истинный репликатор. Как и компьютерный вирус. Как и письма счастья. Но все эти репликаторы нуждаются в сложном аппарате, способствующем им. ДНК необходима клетка, полностью укомплектованная уже существующими биохимическими механизмами, весьма приспособленными для чтения и копирования кода ДНК. Компьютерному вирусу нужен компьютер с каким-либо каналом связи с другими компьютерами, разработанными человеческими инженерами, чтобы повиноваться закодированным инструкциям. Письма счастья нуждаются в хорошем обеспечении идиотами с развитыми мозгами, достаточно образованными, чтобы, по крайней мере, читать. Уникально в первом репликаторе, давшем искру жизни, то, что у него под рукой не было никакого запаса чего-либо приспособленного, разработанного или образованного. Первый репликатор работал de novo, с нуля, без прецедентов и без помощи, за исключением обычных законов химии.
Мощным источником поддержки химической реакции является катализатор, и катализ в некотором виде, конечно, повлек за собой происхождение репликатора. Катализатор – вещество, которое ускоряет химическую реакцию, не будучи потребляемым ею. Вся биологическая химия состоит из катализируемых реакций, и эти катализаторы обычно являются большими молекулами белка, называемыми ферментами. Типичный фермент предоставляет углубления определенной конфигурации в своей трехмерной форме, как сосуды для компонентов одной химической реакции. Он выстраивает их друг напротив друга, вступает с ними во временную химическую связь, подбирая пару с целенаправленной точностью, которую они вряд ли выявили бы в свободном виде.
Катализаторы, по определению, не расходуются в химической реакции, которую они поддерживают, но могут образовываться. Автокаталитическая реакция – реакция, которая производит свой собственный катализатор. Как Вы можете догадаться, автокаталитическая реакция неохотно начинается, но, однажды начавшись, она сама по себе набирает обороты – в самом деле, как лесной пожар, поскольку у огня есть некоторые из свойств автокаталитической реакции. Огонь не строго катализирует, но самовоспроизводит. Химически это процесс окисления, при котором выделяется тепло, и тепло необходимо, чтобы подтолкнуть процесс к началу запуска. Однажды начавшись, он продолжается и распространяется как цепная реакция, потому что вырабатывает тепло, необходимое для своего возобновления. Другая известная цепная реакция – атомный взрыв, в этом случае реакция не химическая, а ядерная. Наследственность возникла как удачный запуск автокаталитического, или же самопорождающего, процесса. Она немедленно набрала обороты и распространилась, как огонь, в конечном счете, приводя к естественному отбору и всему тому, что за этим последовало.
Мы также окисляем углеродосодержащее горючее, чтобы выработать тепло, но не доводим его до воспламенения, потому что производим окисление управляемым способом, шаг за шагом, направляя энергию в полезные каналы вместо того, чтобы рассеять ее как неконтролируемое тепло. Такая управляемая химия, или метаболизм, является столь же универсальной особенностью жизни, как наследственность. Теории происхождения жизни должны объяснять и наследственность, и метаболизм, но некоторые авторы ставили ошибочные приоритетные задачи. Они стремились создать теорию самозарождения метаболизма, и почему-то надеялись, что из этого будет логически вытекать наследственность, как и другие полезные устройства. Но наследственности, как мы увидим, нельзя представить как полезное устройство. Наследственность должна быть первой на сцене, потому что до наследственности сама полезность не имела никакого значения. Без наследственности, и, следовательно, естественного отбора, не было бы ничего, что могло бы быть полезным. Само понятие полезности не может возникнуть, пока не появится естественный отбор наследственной информации.
Самыми ранними теориями происхождения жизни, к которым сегодня относятся серьезно, являются теории A. И. Oпарина в России и Д. Б. С. Холдейна (J. B. S. Haldane) в Англии, написанные в 1920-ых годах, не зная друг о друге. Обе придавали особое значение метаболизму, а не наследственности. Обе натолкнулись на важный факт, что атмосфера Земли до жизни должна была быть «восстановительной», чтобы жизнь могла возникнуть. Этот довольно бесполезный технический термин означает, что в атмосфере отсутствовал свободный кислород. Органические соединения (соединения углерода) при наличии вокруг свободного кислорода уязвимы для сгорания или иного окисления в углекислый газ. Нам, умирающим без кислорода в течение минут, это кажется странным, но жизнь не могла произойти на какой-либо планете со свободным кислородом в атмосфере. Как я уже объяснял, кислород был смертельным ядом для наших самым ранних предков. Все, что мы знаем о других планетах, оставляет мало сомнений в том, что первоначальная атмосфера Земли была восстановительной. Свободный кислород возник позже. Он представлял собой загрязнение окружающей среды продуктами жизнедеятельности зеленых бактерий, сначала свободно плавающих, а позже включенных в клетки растений. В некоторый момент наши предки развили способность справляться с кислородом, а позже пришли к тому, что стали от него зависеть.
Между прочим, упоминание, что кислород произведен зелеными растениями и водорослями, является упрощением. Верно, что растения выделяют кислород. Но когда растение умирает, химические реакции его разложения, эквивалентные сгоранию всех его углеродистых материалов, израсходовали бы количество кислорода, равное всему кислороду, выпущенному этим растением за всю его жизнь. Поэтому не было бы никакой чистой прибавки атмосферного кислорода, если бы не одна деталь. Не все мертвые растения разлагаются. Некоторые из них откладываются в виде угля (или его аналогов), где они изымаются из кругооборота. Если бы все ископаемое топливо в мире было сожжено человечеством, то большая часть кислорода в атмосфере была бы заменена углекислым газом, восстанавливая древнее статус-кво. Это вряд ли случится в ближайшем будущем. Но мы не должны забывать, что единственная причина, по которой у нас есть кислород для дыхания, состоит в том, что большая часть углерода в мире связана под землей. Мы сжигаем все это на свой страх и риск.
Атомы кислорода всегда присутствовали в ранней атмосфере, но не высвобождались в виде газа. Они были связаны в соединениях, таких как углекислый газ и вода. Сейчас углерод главным образом заключен в живых телах или – в намного большем количестве – в горных породах, таких как мел, известняк и уголь, которые происходят из останков некогда живых тел. Во времена Кентербери эти же атомы углерода главным образом содержались бы в атмосфере в виде составных газов, таких как углекислый газ и метан. Азот, ныне основной атмосферный газ, был бы в восстановительной атмосфере соединен с водородом в виде аммиака.
Опарин и Холдейн поняли, что восстановительная атмосфера благоприятна для самопроизвольного синтеза простых органических соединений. Вот собственные слова Холдейна, которые я цитирую из его знаменитой заключительной фразы:
Теперь, когда ультрафиолетовые лучи оказали действие на смесь воды, углекислого газа, и аммиака, получено многообразие органических веществ, включая сахар и, очевидно, некоторые из материалов, из которых построены белки. Этот факт был продемонстрирован Бэйли (E. C. C. Baly) и его коллегами в лаборатории в Ливерпуле. В нынешнем мире такие вещества разлагаются – то есть разрушаются микроорганизмами. Но до возникновения жизни они должны были накапливаться, пока примитивные океаны не достигли консистенции горячего разбавленного бульона.
Это было написано в 1929 году, больше чем за 20 лет до часто упоминаемого эксперимента Миллера и Юри (H. C. Urey), который, как можно было бы понять из сообщения Холдейна, был своего рода повторением эксперимента Бэйли. Однако Э. Ч. Бэйли не изучал происхождение жизни. Его интересовал фотосинтез, и его целью было синтезировать сахар с помощью ультрафиолетовых лучей, направленных в воду, содержащую растворенный углекислый газ, в присутствии катализатора, такого как железо или никель. Именно Холдейн, а не сам Бэйли с его отличительным блеском (Сэр Питер Медавар описал Хэдлайна как самого умного человека, которого он когда-либо знал.) ожидал чего-то замечательного, вроде эксперимента Миллера-Юри, и считал его возвращением к работе Бэйли.
Миллер под руководством Юри взял две колбы, одна выше другой, связанных двумя трубками. Нижняя колба содержала нагретую воду, представляя первобытный океан. Верхняя колба содержала модель первобытной атмосферы (метан, аммиак, водяной пар и водород). Через одну из этих двух трубок пар поднимался от нагретого «океана» в нижней колбе и попадал в «атмосферу» в верхней колбе. Другая трубка шла обратно вниз, из «атмосферы» в «океан». По пути она проходила через искровую камеру («молнии») и камеру охлаждения, где пар конденсировался, формируя «дождь», который пополнял «океан».
Всего через неделю этого подобия рециркуляции океан стал желто-коричневым, и Миллер проанализировал его содержание. Как предсказал Холдейн, он стал бульоном из органических соединений, включая не менее чем семь аминокислот, непременных строительных блоков белков. Среди этих семи были три – глицин, аспарагиновая кислота и аланин – из списка 20, обнаруженных в живых существах. Более поздние эксперименты по этой теме, но с заменой углекислого или угарного газа на метан, достигли похожих результатов. Мы можем сделать разумный вывод, что биологически важные маленькие молекулы, включая аминокислоты, сахара и, в немалой степени, стандартные блоки ДНК и РНК, способны самопроизвольно образовываться при лабораторном моделировании различных версий первобытной Земли Опарина/Холдейна.
До Опарина и Холдейна мыслители, рассуждающие о происхождении жизни, предполагали, что первые организмы должны были являться какой-либо разновидностью растений, возможно, зеленых бактерии. Люди привыкли к идее, что жизнь зависит от фотосинтеза, производства органических соединений, запускаемого солнечным светом, сопровождаемого выделением кислорода. Опарин и Холдейн со своей восстановительной атмосферой подумали о том, что растения вышли на сцену позже. Ранняя жизнь возникла в море существовавших ранее органических соединений. Для еды был бульон, и не было потребности в фотосинтезе – по крайней мере, пока бульон не закончился.
Для Опарина жизненно важным шагом было возникновение первой клетки. И, разумеется, у клеток, как и организмов, есть важное свойство: они никогда не возникают самопроизвольно, а всегда от других клеток. Было простительно отождествлять с появлением жизни возникновение первой «клетки» (метаболизатора), а не первого «гена» (репликатора), как буду делать я. Среди более современных теоретиков с тем же уклоном выдающийся теоретический физик Фримен Дайсон (Freeman Dyson) осознавал и отстаивал это. Большинство современных теоретиков, включая Лесли Оргела (Leslie Orgel) в Калифорнии, Манфреда Эйгена (Manfred Eigen) и его коллег в Германии, и Грэма Кэрнс-Смита в Шотландии – более одинокие индивидуалисты, но это ни в коем случае не перечеркивает первостепенное значение самокопирования, и хронологически, и в отношении его центральности: по- моему, это справедливо.
На что была бы похожа наследственность без клетки? Разве это не проблема курицы-и-яйца? Конечно, да, если мы соглашаемся с тем, что для наследственности требуется ДНК, ДНК не может быть реплицирована без многочисленных вспомогательных молекул, включая белки, которые могут быть созданы только с помощью закодированной информации ДНК. Но только из того, что ДНК – основная самореплицирующая молекула, которую мы знаем, не следует, что она является единственной, которую можно себе представить, или единственной, которая когда-либо существовала в природе. Грэм Кэрнс-Смит аргументировано доказал, что первоначальные репликаторы были неорганическими минеральными кристаллами, с более поздней узурпацией ДНК, вступившей в главную роль, когда жизнь эволюционировала до пункта, где такой переход под генетический контроль стал возможным. Я не буду приводить здесь его доводы, частично потому что я уже сделал свою лучшую попытку в «Слепом часовщике», но также и по более веской причине. Кэрнс-Смит приводит наиболее ясные из прочитанных мною доводов, что самовоспроизведение имело первостепенное значение, и ДНК должна была иметь какого-либо предшественника, природа которого неизвестна, с оговоркой, что тот демонстрировал истинную наследственность. Я считаю позором, что эта неопровержимая часть его аргументов стала связанной в общественном сознании с его более спорными и спекулятивными доводами в пользу минеральных кристаллов в качестве предшественников.
Я не имею ничего против минеральной теории кристаллов, и я разъяснял ее ранее, но что я действительно хочу подчеркнуть, это первенствующую роль репликатора, и есть большая вероятность, что была более поздняя передача управления к ДНК от некоторого предшественника. Я могу подробнее остановиться на этом вопросе, преднамеренно перейдя в этой книге к различным специфическим теориям того, каким мог быть тот предшественник. Каковы бы ни были ее основные достоинства как первоначального репликатора, РНК, конечно, лучший кандидат, чем ДНК, и она была взята за образец предшественника многими теоретиками в их так называемом «мире РНК». Чтобы представить теорию мира РНК, я должен отвлечься на ферменты. Если репликатор – звезда жизненного шоу, то ферменты играют одну из главных ролей, более чем просто второстепенную роль.
Жизнь крайне зависит от виртуозной способности ферментов катализировать биохимические реакции довольно вычурным способом. Когда я впервые узнал о ферментах в школе, расхожее (и, на мой взгляд, ошибочное) мнение, что наука должна преподаваться на основе обыденных примеров, подразумевало, что мы плевали в воду, чтобы продемонстрировать способность слюнного фермента амилазы переваривать крахмал и образовывать сахар. От этого мы получили впечатление, что фермент похож на агрессивную кислоту. Биологические стиральные порошки, которые используют ферменты, чтобы вываривать грязь из одежды, создают такое же впечатление. Но это – разрушительные ферменты, служащие для расчленения больших молекул на их меньшие составляющие. Конструктивные ферменты вовлечены в синтез больших молекул их меньших компонентов, и они делают это, ведя себя как «автоматизированные антрепренеры», и я объясню почему.
Внутренность клетки содержит раствор из тысяч различного рода молекул, атомов и ионов. Они могли бы попарно объединяться друг с другом почти бесконечным числом различных способов, но в основном они этого не делают. Таким образом, есть огромный ассортимент потенциальных химических реакций, которые могут возникнуть в клетке, но большая часть из них не происходит. Помните об этом, размышляя над следующим. В химической лаборатории на полках есть сотни бутылок, все надежно закупоренные, таким образом их содержимое не сталкивается друг с другом, если химик не желает этого, когда добавляет порцию из одной бутылки к порции из другой. Вы можете сказать, что полки в химической лаборатории также предоставляют огромный ассортимент потенциальных химических реакций, которые могут возникнуть. И снова большая часть из них не происходит.
Но представьте себе, что Вы берете все бутылки со всех полок и выливаете их в один чан, наполненный водой. Нелепый акт научного вандализма, все же такой чан в значительной степени являет собой то, что происходит в живой клетке (Хотя, по общему признанию, с большим количеством мембран, которые усложняют картину.). Сотни компонентов тысяч потенциальных химических реакций не удерживаются в отдельных бутылках в ожидании требуемых реакций друг с другом. Вместо этого они все время смешиваются в одном общем пространстве. Но, тем не менее, они ожидают, в основном не вступая в реакцию, пока этого не требуется, как будто находятся в реальных бутылках. Нет никаких реальных бутылок, но есть ферменты, работающие как автоматизированные антрепренеры, или мы могли бы даже назвать их автоматизированными лаборантами. Ферменты способны к распознаванию, почти так же, как радиоприемник, когда ловит отдельные радиостанции, игнорируя сотни других сигналов, одновременно бомбардирующих его антенну неразберихой несущих частот.
Предположим, что есть важная химическая реакция, в которой компонент А объединяется с компонентом B, чтобы образовать продукт Z. В химической лаборатории мы добиваемся этого, взяв с одной полки бутылку с этикеткой А, а с другой полки бутылку с этикеткой B, смешав их содержимое в чистой колбе и обеспечив другие необходимые условия, такие как высокая температура или взбалтывание. Мы добиваемся определенной реакции, которая нам нужна, взяв только две бутылки с полки. В живой клетке много молекул А и много молекул B, плавающих в окружающей жидкости среди огромного разнообразия молекул, где они могут встретиться, но даже при встрече объединяются редко. В любом случае, их встреча не более вероятна, чем тысячи других возможных комбинаций. Теперь мы вводим фермент, названный abzase, который конкретно приспособлен, чтобы катализировать реакцию A+B=Z. В клетке есть миллионы молекул abzase, и каждая действует как автоматизированный лаборант. Каждый лаборант abzase захватывает одну молекулу, не с полки, но свободно плавающую в клетке. Затем он захватывает дрейфующую молекулу B. Он твердо удерживает А своими захватами так, чтобы тот был обращен в определенном направлении. И так же твердо он удерживает B, чтобы он примыкал к A только в правильном положении и ориентации, чтобы скрепить с A и создать Z. Фермент также может делать другие вещи – аналогично лаборанту-человеку уметь обращаться с мешалкой или зажигать Бунзеновскую горелку. Он может образовать временное химическое соединение с A или B, обмениваясь атомами или ионами, которые, в конечном счете, возвращаются, таким образом, получается, что фермент остается таким же, как был, катализатором. В результате всего этого в определенной формы «захватах» молекулы фермента образуется новая молекула Z. Тогда лаборант выпускает новый продукт Z в жидкость и ждет проплывающий мимо другой компонент А, после чего захватывает его, и цикл возобновляется.
Если бы не было никакого автоматизированного лаборанта, то свободно плавающий A иногда врезался бы в свободно плавающий B при правильных условиях образования соединения. Но такое случайное событие было бы редким, не более обычным, чем случайные столкновения с большим количеством других потенциальных партнеров, в которых либо A, либо B могли бы участвовать. Возможно, столкновение с C и создание Y. Или B мог бы врезаться в D и образовать X. Благодаря случайному медленному перемещению все время создается небольшое количество Y и X. Но все зависит от присутствия лаборанта, фермента abzase. В присутствии abzase Z ставится на поток (с точки зрения клетки) в промышленных количествах: фермент обычно спонтанно увеличивает норму реакции в пределах от миллиона до триллиона раз. Если бы был введен другой фермент, acyase, то A соединился бы с C вместо B, снова же на скорости быстро мчащегося ленточного конвейера, создавая щедрый запас Y. Это все те же молекулы А, о которых мы говорили, не ограниченные бутылкой, но свободно объединяющиеся с B или с C, в зависимости от того, какой фермент присутствует для их захвата.
Скорости выработки Z и Y будут, поэтому, зависеть, кроме всего прочего, от того, сколько каждых из двух конкурирующих лаборантов, abzase и acyase, плавает в клетке. А это зависит от того, какой из двух генов включен в ядре клетки. Однако все немного сложнее: даже если молекула abzase присутствует, она может быть инактивирована. Один способ, которым это может случиться, состоит в том, что появляется другая молекула и занимает активную «впадину» фермента. Это – как если бы на автоматизированного лаборанта временно надели наручники. Наручники напоминают мне, между прочим, необходимость исполнить ритуал, предупреждающий что, как всегда бывает с метафорами, есть риск, что «автоматизированный лаборант» может ввести в заблуждение. У молекулы фермента фактически нет рук, чтобы их протягивать и захватывать компоненты, такие как A, уже не говоря о надевании наручников. Вместо этого у него есть специальные зоны на поверхности, к которой A, скажем, обнаруживает сродство, или из-за аккуратного физического соответствия впадине определенной формы, или еще из-за какой-то неясной химической особенности. И это сродство может быть временно сведено на нет способами, которые напоминают преднамеренное переключение выключателя.
Большинство молекул фермента – механизмы особого назначения, которые делают только один продукт: скажем, сахар или жир; пурин или пиримидин (стандартные блоки ДНК и РНК), или аминокислоты (двадцать из них – стандартные блоки природных белков). Но некоторые ферменты больше похожи на программируемые станки, требующие перфоленту, на которой указано, что им делать. Самые выдающийся среди них – рибосомы (Кратко объясненные в «Рассказе Taq».), большие и сложные станки, построенные и из белка и из РНК, которые сами создают белки. Аминокислоты, стандартные блоки белков, уже сделанные ферментами специального назначения и плавающие повсюду в клетке, могут быть захвачены рибосомой. Перфолентой служит РНК, конкретно «информационная РНК» (иРНК). Информационная лента, сама скопировавшая информацию с ДНК в геноме, вводит данные в рибосому и, когда она проходит через «считывающую головку», соответствующие аминокислоты собираются в белковую цепь в порядке, заданном лентой, использующей генетический код.
То, как эта спецификация работает, известно, и это замечательно. Есть набор маленьких транспортных РНК (тРНК), каждая длиной приблизительно 70 оснований. Каждая из тРНК избирательно присоединяется к одному и только одному из двадцати видов обычных аминокислот. На другом конце молекулы тРНК находится «антикодон», триплет, точно дополняющий короткую последовательность иРНК (кодон), который определяет специфическую аминокислоту согласно генетическому коду. Когда лента иРНК движется через считывающую головку рибосомы, каждый кодон иРНК связывается с тРНК, имеющей подходящий антикодон. Это заставляет аминокислоту, свисающую с другого конца тРНК, выстраиваться в ряд в положении, определенном «антрепренером», прикрепляясь к растущему концу формирующегося белка. Как только аминокислота прикрепляется, тРНК уходит на поиски новой молекулы аминокислоты предпочитаемого типа, в то время как лента иРНК медленно продвигается вперед в другую позицию. Таким образом, процесс продолжается, и шаг за шагом формируется белковая цепь. Удивительно, но одна аппаратная лента иРНК может справиться с несколькими рибосомами одновременно. Каждая из этих рибосом перемещает свою считывающую головку вдоль различных частей ленты, и каждая формирует свою собственную копию вновь создаваемой цепи белка.
Каждая новая белковая цепь заканчивается, когда иРНК, вводящая свои данные в рибосому, полностью проходит считывающую головку этой рибосомы, и белок отделяется. Он сворачивается в сложную трехмерную структуру, форма которой определяется, по законам химии, последовательностью аминокислот в цепи белка. Сама эта последовательность обусловлена порядком кодовых символов вдоль иРНК. И этот порядок, в свою очередь, определяется комплементарной последовательностью символов вдоль ДНК, которая составляет основную базу данных клетки.
Закодированная последовательность ДНК, поэтому, управляет тем, что происходит в клетке. Она устанавливает последовательность аминокислот в каждом белке, которая определяет трехмерную форму белка, которая в свою очередь придает этому белку его особые ферментативные свойства. Важно, что контроль может быть при этом косвенным, как мы видели в «Рассказе Мыши», гены определяют, какие другие гены должны включиться и когда. Большинство генов в любой клетке выключено. Поэтому из всех реакций, которые могут произойти в «чане, полном разнородных компонентов», в любой момент фактически происходит лишь одна или две: те, чьи специфические «лаборанты» активны в клетке.
После этого отступления к катализу и ферментам мы теперь переходим от обычного катализа к особым случаям автокатализа, некоторая версия которого, вероятно, играла ключевую роль в происхождении жизни. Вспомните наш гипотетический пример, где молекулы A и B соединяются, чтобы создать Z под влиянием фермента abzase. Что, если сам Z – свой собственный abzase? Я имею в виду, что, если молекула Z имеет как раз подходящую форму и химические свойства, чтобы захватить один компонент A и один B, свести их вместе в правильной ориентации и соединить их, чтобы создать новый Z, точно такой же, как он сам? В нашем предыдущем примере мы могли сказать, что количество abzase в растворе будет влиять на количество произведенного Z. Но теперь если Z и abzase фактически – одна и та же молекула, мы нуждаемся только в одной единственной молекуле Z, чтобы начать цепную реакцию. Первый Z захватывает А и В и соединяет их, создавая больше Z. Затем эти новые Z захватывают больше А и В, образуя еще больше Z, и так далее. Это – автокатализ. При подходящих условиях популяция молекул Z будет расти по экспоненте – подобно взрыву. Такого рода вещи создают обнадеживающее впечатление как составляющие происхождения жизни.
Но это все гипотетически. Джулиус Ребек (Julius Rebek) и его коллеги из института Скриппса в Калифорнии сделали это реальностью. Они исследовали некоторые замечательные примеры автокатализа в реальной химии. В одном из их примеров Z был amino adenosine triacid ester (AATE), A был аминоаденозином, B был пентафторфенил эфиром, и реакция происходила не в воде, а в хлороформе. Само собой разумеется, ни эти специфические химические детали, ни длинные названия, конечно, помнить не обязательно. Важно то, что продукт химической реакции является своим собственным катализатором. Первая молекула AATE сформировалась с трудом, но, будучи однажды сформированной, немедленно запустила цепную реакцию, так как все больше синтезировалось самой AATE, служащей своим собственным катализатором. Словно этого было не достаточно, этот ряд блестящих экспериментов продолжал демонстрировать истинную наследственность в определенном здесь смысле. Ребек и его команда создали систему, в которой существовал больше чем один вариант автокатализируемого вещества. Каждый вариант катализировал свой синтез, используя свой предпочитаемый тип одного из компонентов. Это повысило перспективы истинной конкуренции в популяции образований, демонстрируя настоящую наследственность и поучительную начальную форму дарвиновского отбора.
Химия Ребека очень искусственна. Однако его сообщения красиво иллюстрируют принцип автокатализа, согласно которому продукт химической реакции служит своим собственным катализатором. Это – что-то вроде автокатализа, который необходим для происхождения жизни. Могла ли РНК, или нечто похожее на РНК, в условиях ранней Земли автокатализировать свой собственный синиез в стиле Ребека, и в воде, а не в хлороформе?
Проблема очень сложная, как объяснил немецкий Нобелевский лауреат в области химии Манфред Эйген (Manfred Eigen). Он указал, что любой процесс саморепликации подвергается вырождению в результате копирования ошибок – мутаций. Вообразите популяцию реплицирующихся образований, у которых есть высокая вероятность ошибки в каждом случае копирования. Если закодированное сообщение должно противостоять разрушительному действию мутации, то, по крайней мере, один член популяции в любом поколении должен быть идентичным своему родителю. Если, например, есть десять кодовых единиц («букв») в цепи РНК, средняя доля ошибок на одну букву должна быть меньше, чем одна десятая: мы можем тогда ожидать, что, по крайней мере, у некоторых членов следующего поколения будет полный комплект зачастую исправленных кодовых букв. Но если процент ошибок больше, произойдет неумолимое вырождение в течение поколений буквально из-за единственной мутации, независимо от того, насколько сильно давление отбора. Это называют катастрофой ошибок. Катастрофы ошибок в геномах составляют главную тему интересной книги Марка Ридли (Mark Ridley) «Демон Менделя», но нас в данный момент интересует катастрофа ошибок, которая непосредственно угрожала происхождению жизни.
Короткие цепочки РНК и даже ДНК могут спонтанно самореплицироваться без фермента. Но доля ошибок тогда намного выше, чем в присутствии фермента. И это означает, что прежде чем образовался бы ген достаточной длины, создающий белок для действующего фермента, растущий ген был бы разрушен мутацией. Это – заколдованный круг происхождения жизни. Ген, достаточно большой, чтобы описать фермент, был бы слишком большим, чтобы быть точно реплицированным без помощи фермента того типа, который он пытается описать. Таким образом, очевидно, система не может начать работать.
Разрешением заколдованного круга, которое предлагает Эйген, является теория гиперцикла. Она использует старый принцип – разделяй и властвуй. Закодированная информация подразделяется на субъединицы, достаточно маленькие, чтобы лежать ниже порога катастрофы ошибок. Каждая субъединица – отдельный мини-репликатор, и он является достаточно маленьким, чтобы, по крайней мере, одна копия выжила в каждом поколении. Все субъединицы кооперируются вокруг некоторой важной большей функции, достаточно большой, чтобы перенести катастрофу ошибок, если катализируются одним большим химикатом вместо того, чтобы быть разделенными.
В теории, как я ее описал, есть опасность, что вся система может быть нестабильной, потому что некоторые субъединицы будут самореплицироваться быстрее, чем другие. Здесь вступает в силу хорошо продуманная часть теории. Каждая субъединица преуспевает в присутствии других. Точнее говоря, производство каждой катализируется присутствием другой, так что они формируют цикл взаимозависимости: «гиперцикл». Он автоматически препятствует тому, чтобы любой элемент шел в разнос. Он не может сделать это, потому что зависит от своего предшественника в гиперцикле.
Джон Mэйнард Смит указал на подобие гиперцикла и экосистемы. Количество рыбы зависит от популяции дафнии (водяных блох), которыми они питаются. В свою очередь, количество рыбы влияет на популяцию птиц, поедающих рыб. Птицы создают гуано, которое помогает цветению водорослей, на которых разрастаются дафнии. Полный цикл взаимозависимости является гиперциклом. Эйген и его коллега Питер Шустер (Peter Schuster) предложили некоторый молекулярный гиперцикл как решение загадки замкнутого круга происхождения жизни.
Я собираюсь покинуть теорию гиперцикла в этом месте и возвратиться к предположению, которое полностью с ней совместимо, что РНК в былые времена, когда жизнь только зарождалась, и белки еще не существовали, возможно, выполняла функцию своего собственного катализатора. Это – теория РНК-мира. Чтобы понять, насколько она правдоподобна, мы должны рассмотреть, почему белки способны быть хорошими ферментами, но плохими репликаторами; притом, что ДНК способна быть хорошим репликатором, но плохим ферментом; и наконец, почему именно РНК могла бы быть достаточно хорошей в обеих ролях, чтобы избежать замкнутого круга.
Трехмерная форма в значительной степени несущественна для активности фермента. Белки способны быть хорошими ферментами, потому что они могут принимать почти любую, какую хотите, пространственную форму в трех измерениях как автоматический результат их аминокислотной последовательности в одном измерении. Химическое сродство аминокислот с другими аминокислотами в различных участках цепи определяет конкретный узел, в который скручивается белковая цепь. Таким образом, трехмерная форма белковой молекулы определяется одномерной последовательностью аминокислот, а сама она обусловлена одномерной последовательностью кодовых знаков в гене. В принципе (практика – другое дело, и ужасно трудное) можно записать последовательность аминокислот, которая самопроизвольно свернулась бы почти в любую, какую пожелаете, форму: не только форму, создающую хорошие ферменты, но и любую произвольную, специально выбранную форму (Действительно, есть много различных последовательностей аминокислот, которые приводят к одной и той же форме, что является одной из причин сомневаться относительно наивных расчетов астрономического «неправдоподобия» отдельной белковой цепи, полученной возведением ее длины в 20 степень.). Эти разносторонние таланты белков определяют их способность действовать в качестве ферментов. Белок способен выбрать одну из сотен потенциальных химических реакций, которые могут произойти в клетке, наполненной смесью компонентов.
Белки поэтому образуют замечательные ферменты, способные скручиваться в узлы любой желаемой формы. Но они – отвратительные репликаторы. В отличие от ДНК и РНК, составные элементы которых имеют определенные правила соединения («правило спаривания оснований Уотсона-Крика», обнаруженное этими двумя вдохновленными молодыми людьми), у аминокислот нет таких правил. ДНК наоборот, является отличным репликатором, но отвратительным кандидатом на роль фермента в жизни. Поэтому, в отличие от белков с их почти бесконечным разнообразием трехмерных форм, у ДНК есть только одна форма, известная двойная спираль. Двойная спираль идеально подходит для репликации, потому что две стороны лестницы легко отделяются друг от друга, каждая при этом представляет собой шаблон для присоединения новых букв согласно правилу спаривания оснований Уотсона-Крика. От этого не много пользы для чего-нибудь еще.
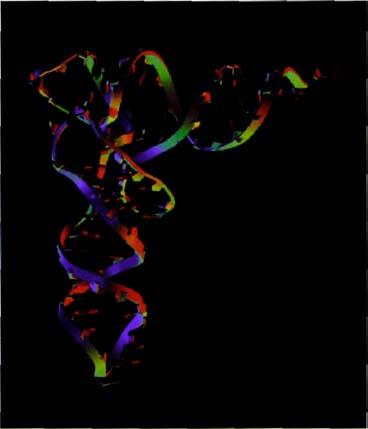
Легко скручивается в узлы. Компьютерная графика транспортной РНК, сдвоенной для создания миниатюрной двойной спирали.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Кентербери
Кентербери Наш “Кентербери” исполнен тайн. В этой точке эволюции появилась жизнь, однако лучше было бы назвать ее точкой происхождения наследственности. Сама жизнь не имеет четкого определения – это факт, который противоречит интуиции и традиции. Пророк (Иезекииль 37),