Митохондриальная Ева и игрек-хромосомный Адам в африканском Эдеме
Митохондриальная Ева и игрек-хромосомный Адам в африканском Эдеме
Сравнительный анализ митохондриальной ДНК (мтДНК) и Y-хромосом современных людей показал, что все современное человечество происходит от небольшой популяции, жившей в Восточной Африке 160–200 тыс. лет назад (Cann et al., 1987). Это подозрительно хорошо совпадает с археологическими данными: древнейшие «анатомически современные люди» найдены как раз в этом районе в отложениях именно такого возраста. Затем в какой-то момент, скорее всего между 100 и 60 тыс. лет назад (опять же, судя по результатам анализа мтДНК), небольшая группа сапиенсов вышла из Африки, и потомки этой группы впоследствии заселили весь мир: от них произошло все современное внеафриканское человечество (см. главу «Великое расселение сапиенсов»). С этим сегодня почти никто не спорит. Главный вопрос в том, только ли от них оно происходит. Смешивались ли африканцы по мере своего расселения с местными евразийскими популяциями или просто вытеснили их?
В течение почти двух десятилетий (с конца 1980-х до второй половины 2000-х) перевес был на стороне второй точки зрения. Главным аргументом были упомянутые результаты анализа мтДНК и Y-хромосомы. Но как на основе анализа ДНК можно судить об истории и миграциях древних людей? Пожалуй, нам нужно поближе познакомиться с методикой и логикой таких исследований.
Сначала на основе полиморфизма (разнообразия) нуклеотидных последовательностей мтДНК людей из разных регионов мира было реконструировано эволюционное (генеалогическое) дерево этой части человеческого генома. Для таких реконструкций существуют специально разработанные, весьма надежные методики. Это похоже на то, как лингвисты восстанавливают историю различных списков с одного исходного текста по изменениям (заменам, пропускам и добавлениям букв или слов), которые неизбежно накапливаются от переписчика к переписчику. Например, если в одном списке имеются ошибки А и Б, в другом — А, Б и В, в третьем — только Б, в четвертом — Б и Г, то логичнее всего предположить, что самый древний список — третий. С него были сделаны первый и четвертый, а затем с первого был сделан второй. Когда тексты длинные (и ошибок поэтому достаточно много), результаты получаются вполне достоверные. Если известно, в каких географических точках были найдены соответствующие рукописи, то можно восстановить пути миграций соответствующих копий.
В случае с ДНК вместо ошибок переписчиков используются случайные мутации (в основном нейтральные, то есть не влияющие на приспособленность и потому незаметные для отбора). Это даже удобнее, потому что люди — и, соответственно, их ДНК — размножаются с определенной частотой, более регулярно, чем средневековые переписчики копировали ту или иную рукопись. К тому же типичная частота возникновения мутаций приблизительно известна. Это аналогично тому, как если бы каждая рукопись переписывалась, например, один раз в 20 лет, а все переписчики делали примерно одинаковое количество ошибок на каждую тысячу слов. Тогда по количеству различий между двумя рукописями можно было бы определить время существования их «общего предка». Для молекул ДНК это вполне возможно (степень точности — вопрос отдельный).
Оказалось, что если построить эволюционное дерево мтДНК современных людей и двигаться по его ветвям сверху вниз (из настоящего в прошлое), то все ветви в итоге сходятся в одну точку во времени и пространстве: Восточная Африка, 160–200 тыс. лет назад. Так появилась в научной печати и в СМИ «митохондриальная Ева» (митохондрии, как мы помним, передаются по материнской линии), а вслед за ней аналогичным образом возник и «Y-хромосомный Адам» (Y-хромосома есть только у мужчин и передается от отца к сыну), живший примерно в то же время и в том же месте.
Эти результаты были восприняты общественностью очень бурно, и, как водится, мало кто понял их истинный смысл. На самом деле ничего удивительного нет ни в Адаме, ни в Еве. Любые гомологичные (то есть имеющие общее происхождение) участки ДНК где-нибудь в прошлом неизбежно сходятся в одну точку, то есть в одну предковую молекулу ДНК. И эта точка вовсе не обязательно совпадает с моментом возникновения вида. Более того, если брать разные гомологичные участки ДНК, каждый из них даст свою, отличную от других «точку схождения».
—————
Откуда берутся митохондриальные Евы
Любая популяция любого вида животных обязательно имела в прошлом свою митохондриальную Еву — последнюю общую праматерь всех ныне живущих представителей данной популяции по прямой материнской линии. Евы появляются автоматически и неизбежно из-за случайных колебаний частот генетических вариантов (например, вариантов митохондриальной ДНК) в популяции. Чтобы понять, почему так получается, рассмотрим простую модель. Допустим, у нас есть популяция, включающая десять самок, у каждой из которых имеется свой вариант мтДНК, немного отличающийся от остальных. Число самок (то есть размер популяции) не имеет принципиального значения: оно влияет только на среднее число поколений, требующихся для того, чтобы одна из этих самок стала Евой. Чем больше популяция, тем дольше придется ждать.
Предположим, что каждая самка оставляет после себя с равной вероятностью либо ноль дочерей, либо одну, либо две. Обозначим исходные варианты мтДНК у наших десяти самок буквами латинского алфавита:
abcdefghij (1)
Это поколение 1. Чтобы смоделировать следующее (второе) поколение, воспользуемся генератором случайных чисел. Сойдет и обычная игральная кость. Нам нужна последовательность из десяти случайных целых чисел в диапазоне от нуля до двух, чтобы определить, сколько дочерей родила каждая самка. Бросаем кость десять раз, если выпадает один или два, записываем ноль, если три или четыре — записываем единицу, если пять или шесть — двойку. У меня получилась такая последовательность:
2 2 1 2 2 1 0 0 1 1
В соответствии с этими числами «родим» для каждой самки дочерей. Учитывая, что дочь наследует мтДНК матери, выпишем распределение митохондриальных гаплотипов (вариантов) в поколении 2:
aabbcddeefij (2)
Можно заметить, что из десяти исходных гаплотипов во второе поколение перешли только восемь. Две самки (g и h) не оставили дочерей, и их митохондриальные линии пресеклись. Самок у нас теперь стало 12, поэтому для того, чтобы смоделировать поколение 3, понадобится 12 случайных чисел. Вот они:
2 2 0 0 2 0 2 1 2 0 2 1
А вот и третье поколение:
aaaaccddeeeiij (3)
В третьем поколении «потерялись» еще два гаплотипа: b и f. Продолжая моделирование, получаем последовательность поколений:
aaaeeejj (4)
eeejj (5)
ejjjj (6)
eejjjjjj (7)
eeejjj (8)
eeejj (9)
eeeej (10)
eeeejj (11)
eeeeeej (12)
eeeejj (13)
eeeej (14)
eeeej (15)
eeeej (16)
eeeeee (17)
Вот и все: к семнадцатому поколению в нашей популяции остался только один митохондриальный гаплотип из десяти исходных. Это значит, что прямые потомки по женской линии остались только у одной из исходных десяти самок.
Процесс этот абсолютно неизбежен: сколько бы мы ни взяли исходных самок с разными гаплотипами, через какое-то число поколений в популяции останется только один из них. При этом одна из древних носительниц этого гаплотипа автоматически превращается в митохондриальную Еву — последнюю общую праматерь всех особей в популяции по непрерывной женской линии.
Кстати, какая именно самка в нашей модели стала Евой для поколения № 17? Думаете, это самка «е» из поколения 1? А вот и нет: у поколения 17 есть и более поздняя общая праматерь. Это самка с гаплотипом «е» из поколения 6. Самка «е» из поколения 1, конечно, тоже является общей праматерью поколения 17 по женской линии, но она не самая поздняя из таких праматерей.
При желании можно придумать фантастические ситуации, в которых Ева никогда не появится. Но эти ситуации не имеют отношения к реальности. Например, Евы не будет, если каждая самка непременно оставляет после себя хотя бы одну дочь (нет ни бездетных самок, ни таких, кто оставил после себя лишь сыновей). Каждому ясно, что так не бывает. Обязательно какая-то часть самок умирает, не родив ни одной дочери. Исходное множество гаплотипов может не сократиться до одного и в том случае, если численность популяции будет бесконечно расти. Но так тоже не бывает: планета не резиновая, рост любой популяции рано или поздно останавливается.

Вымышленный пример генеалогического древа вариантов митохондриальной ДНК. В каждом поколении в популяции присутствует 15 самок, у каждой из которых может быть ноль, одна, две или три дочери. Все самки 16-го поколения восходят к митохондриальной Еве из поколения № 2.
Что будет, если популяция разделится на две — например, часть особей переселится на другой материк, и каждая из двух дочерних популяций будет процветать на своем материке? В этом случае в каждой из двух популяций тоже обязательно рано или поздно закрепится какой-то один из исходного набора гаплотипов. Причем, скорее всего, на разных материках это будут разные гаплотипы. Получится, что у каждой из двух популяций есть своя митохондриальная Ева. Кроме того, будет и третья Ева — общая для обеих групп. Она окажется глубже в прошлом, чем обе «персональные» Евы разделившихся популяций.
—————
По мнению ряда экспертов, приблизительное совпадение результатов по мтДНК и Y-хромосоме — не более чем случайность, отчасти объясняющаяся тем, что оба этих участка генома имеют общее свойство: они присутствуют у каждого человека лишь в одном экземпляре (точнее, в одном варианте: «экземпляров» гораздо больше, они есть в каждой клетке, но все одинаковые). Большинство других участков генома — любые участки ядерных неполовых хромосом — присутствуют в двух вариантах, один из которых получен от отца, другой от матери. Есть еще Х-хромосома, занимающая промежуточное положение: у женщин она присутствует в двух экземплярах, у мужчин в одном.
Американский антрополог и генетик Алан Темплтон еще в 2005 году обратил внимание на тот факт, что ожидаемое время схождения эволюционного дерева, построенного для отдельного участка ДНК, в одну точку зависит от того, в скольких вариантах присутствует данный участок в организме (Templeton, 2005). Быстрее всего должны сходиться как раз мтДНК и Y-хромосома (что и наблюдается, они сходятся 160–200 тыс. лет назад). Это не значит, что именно тогда и появился H. sapiens. По мнению Темплтона, это значит лишь, что данные участки генома не годятся для реконструкции более давних событий. Участки, локализованные на Х-хромосоме, сходятся в более далеком прошлом (до 2 млн лет); все остальные участки — в еще более глубокой древности, некоторые даже еще до того, как разделились эволюционные линии человека и шимпанзе. Как это может быть? Очень просто: если бы мы могли проследить во всех деталях родословную предков человечества — не какого-то конкретного человека, а всей популяции вместе, вплоть до общих предков с шимпанзе, — мы увидели бы, что эта родословная никогда не «схлопывалась» до одного-единственного индивида. Всегда, в любой момент прошлого, существовала некая предковая группа особей с различающимися генами. Поэтому «наша» эволюционная линия вполне могла унаследовать от общих с шимпанзе предков не один, а сразу несколько вариантов того или иного гена. Впоследствии мы могли растерять этот исходный полиморфизм, так что у нас в итоге остались гены, происходящие лишь от одного из исходных вариантов. Именно так и произошло с мтДНК и многими другими генами. Но это не обязательно: по некоторым генам часть исходного полиморфизма могла досуществовать (и досуществовала) до наших дней.
В общем, получается, что история мтДНК — еще не история человечества.
Каким образом по мтДНК или другому участку генома можно сделать вывод о выходе наших предков из Африки в какое-то определенное время? Это возможно в том случае, если вскоре после данного события у кого-то из переселенцев возникла мутация в изучаемом участке ДНК, которая затем в ходе экспансии размножилась. И тогда современный генетик увидит, что частота встречаемости данной мутации у внеафриканского населения, к примеру, 10 %, а в Африке ее нет. Время возникновения мутации определяется на основе других, позднее возникших мутаций, по методу «молекулярных часов». Ну а если вскоре после выхода из Африки в данном участке генома никакой мутации не возникло? Тогда, разумеется, ничего не выйдет: этот участок генома просто не сохранит следов интересующей нас экспансии.
Поэтому по одному-единственному участку генома (например, по мтДНК) нельзя делать окончательные выводы об эволюции и истории расселения человечества. Для таких выводов необходим комплексный анализ многих разных участков генома.
Темплтон проанализировал помимо мтДНК и Y-хромосомы еще 23 участка генома и пришел к следующим выводам. Разные участки ДНК сохранили следы разных событий в истории человечества. Общая картина довольно точно совпадает с той, которая реконструируется по данным археологии. Три участка ДНК сохранили следы древнейшей волны выхода из Африки около 1,9 млн лет назад. Это означает, что в наших жилах течет кровь древних азиатских (а не только африканских) эректусов!
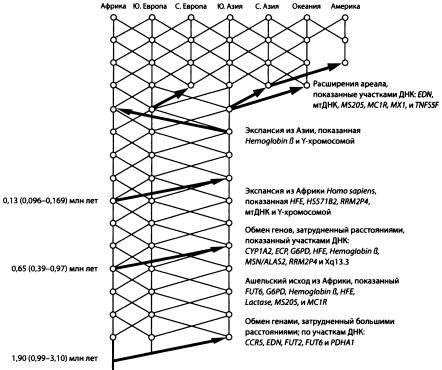
Схема истории человечества по А. Темплтону. Справа перечислены участки ДНК (гаплотипы), подтверждающие то или иное событие. Видно, что мтДНК и У-хромосома «работают» лишь начиная с последнего выхода из Африки около 100 тыс. лет назад. По рисунку из Templeton, 2005.
Семь участков ДНК свидетельствуют о втором исходе из Африки около 0,65 млн лет назад. Это, наверное, позднеашельская экспансия. Гейдельбергские люди, представители этой волны, — тоже наши предки. Наконец, еще пять участков ДНК (в том числе мтДНК и Y-хромосома) подтверждают третий исход из Африки около 130 тыс. лет назад.
Кроме того, данные Темплтона показывают, что обмен генами между евразийскими и африканскими популяциями наших предков практически никогда не прекращался, хоть и был сильно затруднен большими расстояниями. Получается, что древнее человечество все-таки не было совокупностью полностью изолированных популяций (рас, подвидов, видов?) — оно было относительно единым на протяжении двух последних миллионов лет.
Выводы Темплтона оспаривались одними экспертами, в то время как другие разными путями приходили к похожим результатам. Но вплоть до публикации сенсационных результатов прочтения геномов неандертальца и денисовца (речь о которых пойдет ниже), преобладала точка зрения о полном вытеснении африканскими сапиенсами всех древних евразийских популяций.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
2. Хромосомный набор клетки
2. Хромосомный набор клетки Важная роль в клеточном цикле принадлежит хромосомам. Хромосомы — носители наследственной информации клетки и организма, содержащиеся в ядре. Они не только осуществляют регуляцию всех обменных процессов в клетке, но и обеспечивают передачу
Адам запаздывает
Адам запаздывает Очевидно, что сразу же возникает вопрос: «Действительно ли большое количество полиморфизмов Y-хромосомы указывает на африканское происхождение современного человека?» Ответ на него — «Да, несомненно», и результаты научного исследования,
После обезьян были «Адам» и «Ева»
После обезьян были «Адам» и «Ева» Разгадка вопроса о происхождении человека в конечном итоге оказалась спрятанной в самом геноме человека. Не зря же некоторые ученые называют ДНК «величайшим археологическим ископаемым всех времен». Природой, как бы специально для
После обезьян были «Адам» и «Ева»
После обезьян были «Адам» и «Ева» Разгадка вопроса о происхождении человека в конечном итоге оказалась спрятанной в самом геноме человека. Не зря же некоторые ученые называют ДНК «величайшим археологическим ископаемым всех времен». Природой, как бы специально для
Адам и Ева
Адам и Ева Когда говорят, что человек произошел от обезьяны, да еще ссылаются при этом на Дарвина, это не совсем грамотно. В «Происхождении человека»[29] сказано: человек и современные человекообразные обезьяны имеют общего предка. Разумеется, этот вымерший предок тоже
Глава XIII К КОМПАНИИ ПРИСОЕДИНЯЕТСЯ АДАМ
Глава XIII К КОМПАНИИ ПРИСОЕДИНЯЕТСЯ АДАМ История, которую я излагаю в этой книге,— это история мира, запечатленная в гене, наиболее легко поддающемся расшифровке — митохондриальной ДНК. Можно сказать, что это весть от Евы. Изящество и простота, с которой митохондриальная
Хромосомный этап формирования пола
Хромосомный этап формирования пола Пол начинает определяться во время оплодотворения. В ядрах клеток человека одна пара хромосом различна у мужчин и женщин. У женщин эта пара под микроскопом похожа на две буквы X, а у мужчин – на буквы XY. Соответственно, эти хромосомы и
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва)
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва) Полное алиментарное голодание успешно применяется при лечении некоторых психических и
Адам и Ева
Адам и Ева Наши подробные изыскания в плане развития мужского и женского организма, возможно, вызвали у некоторых особо любознательных читателей в памяти историю о божественном творении человека. Как известно, Господь вначале сотворил Адама (то есть, мужчину) и лишь
Глава 6. Адам и Ева (разные и равные)
Глава 6. Адам и Ева (разные и равные) Мужское и женское тело Антропологические исследования показывают, что мужчины и женщины различаются по строению тела. У мужчин преобладает мышечный тип конституции: для них характерны широкие плечи и узкие бедра. У женщин преобладают