Глава VII КОРНИ МЕЗОФИТА
Глава VII
КОРНИ МЕЗОФИТА
В книге П. Эрлиха и Р. Холма «Процесс эволюции» есть выражение, которое мне сразу очень понравилось, и я его потом не раз использовал. Они разбирают известный «биогенетический закон» Э. Геккеля, гласящий, что онтогенез повторяет филогенез, т. е. каждый организм в индивидуальном развитии проходит стадии эволюции предков. Эрлих и Холм пишут, что на недостатки этой идеи, заставляющей каждый организм снова взбираться на свое филогенетическое древо, указывают почти все современные исследователи, а она все же «занимает видное место в биологической мифологии». Перед нами действительно биологический миф, это не просто ошибка, а скорее заблуждение от недостатка компетентности, прочно удерживающееся в поколениях. Мифический «биогенетический закон» лишь один пример, число которых можно легко умножить. Приходится признаться, что в науке мифы играют немалую роль.
Я не берусь объяснять причину их возникновения и поразительной устойчивости. Возможно, причина отнюдь не в инертности и не в неумении критически относиться к прочитанному и услышанному, а просто в самой сложности науки. Ученый не в состоянии переосмыслить и проверить каждое наблюдение и каждое обобщение. Что-то приходится брать на веру, полагаясь на компетентность коллег. Броские, но упрощенные обобщения особенно быстро распространяются и легко запоминаются, Когда потом оказывается, что на самом деле все сложнее, то такое усложненное представление уже труднее распространяется за пределы круга компетентных специалистов. Заблуждение, старая точка зрения не вытесняется новым, усложненным взглядом. Примерно так произошло и с «биогенетическим законом». Современные представления о соотношении онтогенеза и филогенеза уже не уложить в простенькую формулу. Естественно, что они не смогли вытеснить «биогенетический закон», к тому же якобы подтверждаемый таким замечательным и всем известным фактом, как зачатки жаберных щелей у человеческого зародыша.
О некоторых мифах уже шла речь выше (например, о климатах карбона и перми, описываемых в школьном учебнике биологии). О других пойдет речь в этой и следующей главе. Читатель помнит, что в становлении геологии и биологии немалую роль сыграло то, что основной материал для наблюдения сначала был европейским. Все европейское долго казалось, а отчасти кажется и до сих пор типичным, закономерным, а все за пределами Европы — или дополнением к правилу, или исключением из него.
Еще в прошлом веке были выделены палеозойская, мезозойская и кайнозойская эры, причем решающую роль в их выделении сыграла палеозоология. Палеозой — время трилобитов, панцирных рыб, древнейших четвероногих. Мезозой — время динозавров, ихтиозавров, аммонитов и белемнитов. Кайнозой — время млекопитающих. Со времен Дарвина и Лайеля в книгах изображают международную стратиграфическую шкалу, где слева приводят периоды, эпохи и века, а справа перечисляют именно эти основные группы организмов. Что касается растений, то о палеозое принято говорить как о времени споровых растений — папоротников, древовидных каламитов и плауновидных, о мезозое — как об эре голосеменных, кайнозой считают эрой покрытосеменных.
Впрочем, еще в прошлом веке возникла неувязка в сопоставлении эр животной и растительной жизни. Европейская пермь делится на две части. В ранней перми растительный мир еще сохранял многие черты предшествующего каменноугольного периода. В поздней перми главную роль играли хвойные, внешне сходные с триасовыми, т. е. мезозойскими. Споровых растений в поздней перми Европы очень мало, и среди них нет ни каламитов, ни древовидных плауновидных. Поздняя пермь по растениям казалась гораздо ближе к мезозою, чем к ранней перми. Не менее ярко несоответствие на рубеже мезозоя и кайнозоя. Если флора раннего мела еще типично мезозойская, то в позднем мелу Европы господствовали покрытосеменные, так что облик захоронений растительных остатков типично кайнозойский.
Так появилось убеждение, что деление истории Земли по эволюции животных и растений различно и что крупнейшие этапы эволюции растений на полпериода опережают этапы в эволюции животных. Отсюда следовали важные выводы. Во-первых, надо вводить самостоятельные эры для растений. Их назвали палеофитом, мезофитом и кайнофитом. Во-вторых, поскольку этапы эволюции растений меняются раньше, чем у животных, в смене первых можно было видеть причину, а в смене вторых — следствие. В цепях питания первыми меняются растения, затем меняются травоядные, а за ними и плотоядные животные. На перераспределение цепей питания и уходит полпериода. Известный палинолог Р. Потонье даже провозгласил соответствующий закон «эволюционного несогласия».
Закон «эволюционного несогласия» был сформулирован сравнительно недавно, в 50-х годах, и лишь подвел некоторый итог взглядам, сложившимся задолго до этого, в 60 — 70-х годах прошлого века. В нынешних статьях и книгах, в том числе и учебниках, можно прочесть примерно такое же изложение истории органического мира. Если же внимательно ознакомиться с фактическими материалами, то чуть ли не все в этой картине оказывается неверным, причем материалов для исправления ошибок было вполне достаточно еще 20–30 лет назад. В этой главе я постараюсь развенчать мифы о событиях на рубеже палеозоя и мезозоя, а в следующей главе мы поговорим о том, что произошло на рубеже мезозоя и кайнозоя.
Начнем с того, что считать палеозой временем расцвета и господства споровых растений можно было лишь тогда, когда все растения с папоротниковой листвой принимались за настоящие папоротники и когда не были известны ангарская и гондванская флоры. Теперь хорошо известно, что очень многие палеозойские растения с листвой папоротникового типа, характерные для европейского карбона и ранней перми, имели семена и принадлежат к «семенным папоротникам» (птеридоспермам). Само слово «папоротники» в их названии сохраняется по традиции, подобно тому как один из лишайников мы по-прежнему зовем оленьим мхом. В Ангариде и Гондване в отличие от Европы уже со среднего-позднего карбона главную роль играли голосеменные, а роль споровых была второстепенной. Поэтому если бы мы членили историю растений применительно к Сибири, то эру голосеменных пришлось бы начинать со среднего карбона. С другой стороны, для мезозоя Сибири особенно характерно большое количество папоротников — их здесь не меньше, чем голосеменных. Вообще для Сибири становится неверным характеризовать палеозой как время споровых, а мезозой — как время голосеменных.
Я убежден, что выделять некие планетарные эры в эволюции растений — довольно безнадежное предприятие. Невозможно подстричь под гребенку единых этапов все растительное население Земли. Правящие династии растений менялись от одной области к другой, и все подгонять под европейский стандарт так же неуместно, как привязывать историю европейских народов к смене династий китайских императоров. Поэтому сами понятия «палеофит», «мезофит» и «кайнофит», если их сохранять, наполняются существенно разным содержанием, когда мы говорим о Сибири, Европе или Индии.
Еще более сомнителен вывод, что эволюция растений опережает эволюцию животных. Цепи питания — важный компонент эволюции органического мира. Наверное, многие эволюционные события прямо или косвенно связаны с тем, кто кого ел. Только я не думаю, что мы можем ставить в прямую связь широкое расселение покрытосеменных в середине мелового периода и вымирание аммонитов и белемнитов в конце мела. Если это звенья одной цепи питания, почему-то разматывавшейся несколько десятков миллионов лет, то неплохо было бы указать хотя бы главные элементы этой цепи. Тогда можно всерьез говорить и о гипотетическом законе «эволюционного несогласия».
Обсуждая общие закономерности эволюции органического мира, исследователи часто опирались на некие обобщенные списки растений и животных, составленных чуть ли не для всей Земли в целом, отвлекаясь от тех конкретных ассоциаций животных и растений, с которыми мы встречаемся в отдельных биогеографических областях. До сих пор никто как следует не сопоставил соотношение ассоциаций наземных травоядных позвоночных и комплексов растительных остатков того же района. Мы пока не имеем представления о том, какими растениями питались палеозойские и мезозойские травоядные и какие хищники им досаждали. С другой стороны, можно предполагать, что диета травоядных, относившихся к одному семейству и близкородственным родам, но жившим в разных палеофлористических областях, различалась достаточно сильно. Скажем, травоядные позвоночные поздней перми и раннего триаса, жившие в Гондване и Западной Ангариде, — довольно близкие родственники, но питаться им приходилось совсем разными растениями. Поэтому разумно предположить, что представители одной систематической группы травоядных могли питаться разными растениями не только в пространстве, но и во времени, т. е. существенно менять диету без большого ущерба для здоровья.
Многие из тех, кто писал о мезозойском облике позднепермской флоры и относил ее поэтому к мезофиту, не учитывали источника этого представления. Оно возникло после изучения позднепермской флоры Западной Европы. Верхнепермские растительные остатки захоронены здесь в медистых сланцах мощностью всего в несколько десятков сантиметров. Большей частью это остатки четырех родов хвойных, ни один из которых не известен в триасе. Нередко встречается один вид птеридосперма Lepidopteris, другие виды которого характерны для триаса. Остальные растения медистых сланцев редки, плохо изучены, и ничего определенного об их связях с мезозойскими растениями сказать нельзя.
Упрощенными оказались и представления о возрасте флоры медистых сланцев Европы. Они принадлежат «цехштейну» (средневековые рудокопы назвали так толщу морских отложений, составляющих верхнюю часть европейской перми). После того как были изучены микрофауна и палинологические комплексы цехштейна, оказалось, что он (а тем самым и вся европейская верхняя пермь) соответствует лишь верхним частям верхней перми в других областях. Короче говоря, если придерживаться обычных взглядов на объем верхней перми (учитывая и расхождения во взглядах), то так или иначе получается, что флора цехштейна, раньше олицетворявшая верхнепермскую флору вообще, всего лишь некая частная флора, распространенная только в Европе и только в конце перми, к тому же составленная родами или вымирающими в конце перми, или неизвестно как связанными с мезозойскими родами. Только один вид цехштейновой флоры действительно связывает ее с мезозоем.
В который уже раз мы сталкиваемся с тем, что новые исследования, вместо того чтобы дополнять и подправлять устоявшиеся взгляды, подтачивают их и наконец обращают в миф. Мы многое узнаем попутно, а от главного вопроса, на который хотели получить ответ, оказываемся дальше, чем думали в начале работы. Достаточно уверенно можно говорить только о совершенных в прошлом ошибках. Главная уже упоминалась. Изучая эволюцию растительного мира, нельзя механически суммировать списки растений, найденных на разных материках, как нельзя выдавать частную флору за олицетворение всего, что было в разных местах Земли. Надо изучать становление мезозойской флоры по конкретным областям, прослеживая истоки каждой области в палеозое. Надо помнить о том, что эволюция растений протекала в экосистемах, судьба которых контролировалась различными факторами, в том числе и небиологическими. Геологические перестройки открывали и перекрывали миграционные тракты. Морские бассейны и горные цепи имели для эволюции растений такое же значение, как государственные границы для истории народов. Открытые границы ведут к усилению связей, интеграции экосистем. Закрытые границы порождают обособление. Эволюция по разные стороны от границы протекает более автономно. Нельзя забывать и об изменениях климата.
О расшифровке корней мезозойской флоры в палеозое не может быть речи, если нет надежной систематики растений. Случаев, когда палеоботаники попадались на внешнем сходстве неродственных растений, не перечесть. Примеры, когда обманчивое сходство растений приводило к неверным фитогеографическим выводам, уже приводились. Не меньше недоразумений происходило и при анализе исторической преемственности флор.
В перми Гондваны известны членистостебельные, листья которых срослись по несколько штук в ланцетные лопасти. Похожие растения нашли в триасе Европы и сделали вывод, что европейская триасовая флора генетически связана с гондванской позднепалеозойской. Может быть, это и так, но органов размножения гондванских членистостебельных с такими листьями мы до сих пор не знаем. Без этого рискованно заключать и об исторической преемственности. В гондванском палеозое есть род Buriadia. Его ветки совсем как у хвойных, и их сначала отнесли к европейскому триасовому роду Voltzia, а когда отнесли к самостоятельному роду, то все равно оставили в семействе вольциевых. Вот якобы и еще одно подтверждение родства обеих флор. Теперь мы знаем, что органы размножения буриадий и вольций совершенно различны, а сходство их побегов мало что значит для выяснения флористических связей. Вывод очевиден: для сравнения флор, для суждений об их происхождении нужно детальное изучение растений, надо получить достаточно полные реконструкции растений, знать их анатомию, органы размножения.
К сожалению, выполнить все эти требования слишком непросто. Поэтому убедительные данные о происхождении мезозойской флоры накапливаются медленнее, чем хотелось бы. Мы вынуждены довольствоваться отдельными штрихами сложной картины, причем на каждый штрих уходят годы работы. Сужу об этом по собственному опыту.
С проблемой происхождения мезозойской флоры я столкнулся в общем-то случайно, еще занимаясь систематикой сибирских кордаитов. Мой университетский однокашник В. А. Селиверстов принес мне коробочку с несколькими отпечатками, найденными в верхнепермских отложениях в Вязовском овраге недалеко от Оренбурга. Отпечатки были покрыты угольной корочкой, из которой, судя по всему, можно было приготовить препараты кутикулы и изучить клетки кожицы. Похожие отпечатки тоже из Южного Приуралья незадолго до этого прислали М, Ф. Нейбург саратовские геологи. Тогда я составил опись образцов. Нейбург дала заключение о возрасте, и отпечатки предполагалось выбросить. На них почти не было видно жилок, а поэтому точное определение казалось невозможным. Почему-то образцы саратовцев остались лежать в посылочном ящике, и я вспомнил о них, только познакомившись с селизерстовскими отпечатками. На них тоже жилки почти не были видны, хотя сохранность угольной корочки была хорошей.
То, что при хорошей сохранности угольной корочки жилки плохо пропечатались на породе, показалось мне не случайным. Когда-то Нейбург поручала мне рисовать отпечатки триасовых птеридоспермов, у которых было то же самое. Я проклинал все на свете, пытаясь разглядеть направление жилок при освещении — косым светом то с одной стороны, то с другой. Не такой уж вроде бы существенный признак, хорошо ли пропечатались жилки. Скорее его надо относить за счет условий сохранности, а не своеобразия растений. Возможно, что я не придал бы этому признаку значения, если бы не сделал препаратов кутикулы с селиверстовских образцов. Получились препараты хорошо, можно было разглядеть на кутикуле каждую клетку, каждое устьице. Устьица оказались такими же, как у упомянутых триасовых птеридоспермов. Возникло подозрение, что это растения одной группы.
Коллекции от геологов, работавших в Южном Приуралье и в других районах европейской части СССР, продолжали поступать. В моем распоряжении были и коллекции, ранее полученные Нейбург от геологов. Собирался порядочный материал, с каждым годом все более интересный. Главный его смысл был в том, что он характеризовал флору Западной Ангариды, т. е. ее части, расположенной западнее Урала, и мог быть надежно привязан к стандартной стратиграфической шкале ярусов Перми. Это было важно для правильной датировки других пермских флор Ангариды, в том числе сибирских, чей возраст служил источником нескончаемых дискуссий.
К этому времени возня с кордаитовыми листьями, которым я уже посвятил почти 10 лет, успела изрядно надоесть. Появилась возможность заняться флорой, стратиграфически очень важной, но изученной еще очень плохо. Правда, первые образцы с отпечатками растений из перми Приуралья изучал еще А. Броньяр и немногие другие европейские палеоботаники. Часть из этих образцов привез из России известный английский геолог Р. И. Мурчисон, проехавший до Урала и установивший в результате путешествия пермскую систему. Он и назвал ее по тогдашней Пермской губернии. Потом отдельные коллекции описывал И. Ф. Шмальгаузен, а уже в нашем веке — М. Д. Залесский. Последний опубликовал в 1927 г. прекрасный атлас «Палеозойская флора уральских пределов Ангариды», где изобразил лучшие из собранных до него образцов, в том числе и из частных коллекций. Некоторые из них были собраны в Приуралье при разработке медных руд (медистых песчаников). Впоследствии Залесский вместе со своей сотрудницей Е. Ф.Чирковой объехал многие местонахождения и описал часть коллекций.
Нельзя сказать, чтобы западноангарская флора была совсем плохо изучена, когда я собрался ею заняться. Число описанных видов было довольно большим — много десятков. Воспользоваться же этими описаниями было почти невозможно: они были короткими, иллюстрации — скудными, а главное, не изучались детали жилкования, строение кутикулы, органы размножения растений. Более того, наиболее важные образцы оказались давно потерянными (в частности, почти вся коллекция Залесского погибла во время войны), а на оставшихся образцах то не было угольной корочки, то отпечатки плохо сохранились.
Коллекции, переданные геологами и хранящиеся в моих шкафах, были достаточно обширны, но и они не внушали полного доверия. Часто геологи собирают не те отпечатки, которые наиболее интересны, а то, что попалось на глаза и привлекло внимание. Когда я смотрел одну коллекцию, привезенную из окрестностей Кунгура, то обратил внимание, что в ней нет листьев кордаитов. Решив, что в этом кроется интересная закономерность, я поделился своими мыслями с человеком, собравшим образцы. «Да что вы! — услышал я в ответ. — Там было много кордаитов, но я не брал их! Целых листьев не нашел, а обрывки такие скучные!» Короче говоря, надо было поехать по главным местонахождениям самому.
В целом поездки были удачными, удалось разыскать многие классические местонахождения растений. В некоторых случаях в розысках помогли старожилы. Один из них прекрасно помнил приезд М. Д. Залесского и повел меня туда, где собирал растения «профессор из центра». Довелось побывать и там, где вел раскопки В. П. Амалицкий в поисках гондванских позвоночных и где он нашел листья, принятые им за глоссоптериевые. На Малой Северной Двине, там, где раньше неподалеку стояла деревня Соколки, сохранился гигантский раскоп, где поместился бы большой многоквартирный дом. Жители окрестных деревень до сих пор помнят рассказы отцов и дедов, на которых произвели впечатление и найденные скелеты «ящеров», и щедрая оплата нелегкого труда, и бесплатно раздававшаяся рабочим мазь от комаров. К сожалению, найти в этом раскопе ископаемые растения нам не удалось (правда, в нем выросла отменная современная смородина).
Коллекции собрались огромные — многие тысячи отпечатков из десятков местонахождений. Только усевшись за эти коллекции, я понял, какая трудная работа предстоит. Изучение материала затянулось и продлится еще не один год. Как ни приятно иметь дело с материалом хорошей сохранности, но корпеть над ним надо куда дольше, чем над отпечатком, на котором ничего не видно, кроме оттисков жилок. С иным образцом приходится заниматься неделями. Поэтому сейчас рано делать определенные выводы. Зато можно более осмысленно сформулировать некоторые проблемы, в том числе и проблему происхождения мезозойской флоры.
То, что изучение западноангарской флоры поможет разобраться в происхождении мезозойской флоры, стало понятно довольно быстро. Из Вязовского оврага, откуда были образцы В. А. Селиверстова, в первую же поездку в 1965 г. я привез много образцов. Помимо все тех же листьев с неясным жилкованием и хорошей угольной корочкой удалось найти и интересные органы размножения. Внешне они похожи на небольшой гриб: тонкая ножка, а на конце шляпка. Край шляпки фестончатый, а поверхность с радиально расходящимися от центра ребрами. Из угольной корочки, составляющей шляпку, получился неплохой препарат кутикулы. Под микроскопом можно было хорошо видеть устьица, такие же как и на селиверстовских листьях. В Вязовке нашлись и листья, угольная корочка которых была тонкой и на свет можно было видеть черные тяжи жилок. Они шли плотным пучком вдоль оси листа, а затем расходились к краям. На иных экземплярах можно было рассмотреть оттиски жилок, отпечатавшихся на породе. Там были какие-то дополнительные складки, их можно было принять за соединения между жилками.
Возникло предположение, что именно эти листья В. П. Амалицкий, а затем М. Д. Залесский приняли за глоссоптериевые. Я вспомнил, что в Палеонтологическом музее АН СССР им, Ю. А. Орлова, где хранятся найденные Амалицким скелеты, есть и привезенные им отпечатки растений. Предположение в общем подтвердилось. Отпечатки были действительно очень похожи, но на образцах Амалицкого не было угольной корочки, чтобы окончательно отождествить листья с Малой Северной Двины и Вязовского оврага. Во избежание недоразумений пришлось обозначать вязовские листья другим родовым названием. В качестве такового я выбрал Tatarina — по татарскому ярусу, в отложениях которого листья были найдены.
Палеоботаники часто вынуждены использовать разные родовые названия для листьев, у которых известно или нет строение кутикулы. Нередко листья, неотличимые по форме и жилкованию, принадлежат разным систематическим группам (например, цикадовым и беннеттитам), и установить это можно только по клеточному строению кожицы, отпечатавшемуся на кутикуле. Листья с неизвестным строением кожицы могут принадлежать любой из групп, но как-то надо их называть. Для таких листьев вводят особый род, который не относят ни к одной из групп. Поскольку отпечатки, собранные Амалицким и описанные Залесским сначала как Glossopteris, а затем как Pursongia, могли иметь разную кутикулу, в том числе и отличающуюся от той, что я видел у вязовских листьев, было рискованно называть последние Pursongia. Это название можно было оставить для всех отпечатков сходного облика и лишенных остатков кутикулы.
Я уже говорил, что по строению кутикулы татарины Вязовки оказались похожими на триасовые птеридрспермы. Более важным было то, что эти триасовые птеридоспермы — семейство пельтаспермовых — имели такие же, как в Вязовке, грибообразные органы размножения. Их еще в 30-х годах детально описал известный английский палеоботаник Т. М. Гаррис. Под шляпкой «гриба» прикреплялись семена, по опадении которых оставались рубцы. В этом месте кутикула прерывалась отверстиями. Уже потом я обнаружил оттиски округлых рубцов от опавших семян и у органов размножения татарин.
Большое количество этих растений в те же годы привезли из скважин Севера европейской части СССР геологи А. Г. Олферьев и В. Р. Лозовский. Местами порода была превращена в сланец из-за огромного скопления листьев татарин. Тут же встречались и «шляпки» (рис. 19). Количество татарин в отложениях татарского яруса (последнего яруса перми) было примерно таким же, как кордаитов в перми Сибири. Ясно, что это была главная группа растений в конце перми по всей Западной Ангариде. В те же годы И. А. Добрускина занималась триасовыми птеридоспермами и пришла к выводу, что пельтаспермовые гораздо более характерны для триасовых флор, чем можно было полагать раньше. Таким образом установилась первая достоверная прямая связь между пермской и триасовой флорами, подтвержденная и изучением микроструктуры листьев, и знанием женских органов размножения.
Одновременно стала ясна ошибка и Амалицкого, и Залесского, и последующих палеоботаников, связывавших пурсонгии (и, стало быть, татарины) с гондванскими глоссоптериевыми.
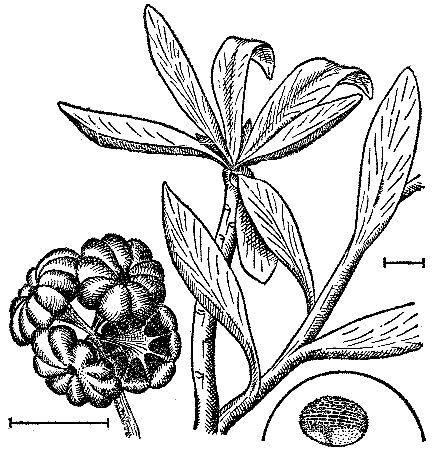
Рис. 19. Облиственный побег верхнепермской татарины (Tatarina) и связываемые с такими растениями женские органы размножения (Peltaspermum) слева и пыльца (Vittatina) в кружке; длина линейки 1 см
Связь пермской флоры Западной Ангариды потянулась не в палеозой Гондваны, а в мезозой северных материков, поскольку в триасе пельтаспермовые особенно характерны для Европы, Северной и Средней Азии, Китая. Их остатки находили и в Гондване, но тоже только в триасе. Так что можно полагать, что они проникли сюда с севера. Древнейшие достоверные остатки пельтаспермовых птеридоспермов сейчас известны в кунгурском ярусе (последнем ярусе нижней перми) Среднего Приуралья. Круг предшественников мезозойской флоры, обнаруженных во флоре Западной Ангариды, не ограничивается пельтаспермовыми. Другие растения были известны и И. Ф. Шмальгаузену, и М. Д. Залесскому, но они почему-то не разглядели в них предтеч мезозоя. Впрочем, Шмальгаузен вообще мало занимался этой флорой, да и коллекции у него были небольшими. Что же касается Залесского, то он, по-видимому, был целиком поглощен идеей о близком родстве ангарской и гондванской флор. Кроме того, расшифровка общности западноангарских и мезозойских растений была невозможна без достаточно детальных исследований. Внешне же языковидные листья пурсонгий-татарин гораздо больше похожи на глоссоптериевые, чем на сильно рассеченные папоротникообразные листья триасовых пельтаспермовых. Примерно то же случилось и с другими растениями, которые, как я склонен считать, дали начало мезозойским родам.
В западноангарской флоре эти растения можно проследить до отложений кунгурского яруса. Глубже их корни теряются. В отложениях предшествующего, артинского яруса остатков растений уже не так много, а еще ниже по разрезу историю западноангарской флоры можно проследить только по палинологическим данным, т. е. по остаткам спор и пыльцы. Пока, к сожалению, это только принципиальная возможность, которую трудно реализовать. Мы уже знаем состав спор и пыльцы в докунгурских отложениях Западной Ангариды, но не знаем, каким растениям принадлежали те или иные формы спор и пыльцы.
Некоторые палинологи пытались решить эту задачу, сравнивая пермскую пыльцу с пыльцой современных растений. Скажем, у многих современных хвойных пыльца снабжена воздушными мешками, чаще всего двумя. Такую же пыльцу находят и в пермских отложениях, так и называя ее пыльцой хвойных. Такой путь более чем рискованный, поскольку палеоботаникам известно несколько групп растений (помимо хвойных), обладавших пыльцой с двумя мешками. Это различные птеридоспермы и растения с глоссоптериевой листвой.
Первоначально я надеялся проследить с помощью палинологии хотя бы историю пельтаспермовых птеридоспермов. Их пыльца была изучена у триасовых представителей, и можно было надеяться, что и в перми она была того же облика — в виде французской булки. Я попросил палинологов обработать несколько образцов, в которых было особенно много листьев татарин. Мне принесли пробирки с выделенной из породы пыльцой. Тщетно я просматривал под микроскопом препарат за препаратом. Пыльцы в них было очень много, и самой разнообразной, но ожидавшейся не было. Зато было особенно много пыльцы с многочисленными ребрами (род Vittatina) и мешковой с ребристым телом. Оставалось предполагать или то, что пыльца моих пельтаспермовых не сохранялась в ископаемом состоянии (такие растения с крайне неустойчивой к захоронению пыльцой известны), или что она была другого облика, чем в триасе. Мысль, что татарины принадлежат не пельтаспермовым, а другим растениям с иной пыльцой, я не допускал, поскольку знал их женские органы размножения и строение кутикулы листьев. Вскоре А. В. Гоманьков, с которым я разделил изучение западноангарской флоры и который занялся пельтаспермовыми, обнаружил, что иногда под «шляпками» их органов размножения огромными массами скапливается пыльца типа Vittatina. Трудно поверить, что так поведет себя пыльца чужого растения. Почти одновременно как будто решился вопрос и с мешковой пыльцой, имеющей ребристое тело. Я копался с непонятным обугленным остатком, найденным вместе с татаринами и похожим на какую-то сильно деформированную, сплющенную шишку. Разглядеть в обугленной лепешке ничего не удавалось, и я решил обработать химически небольшой кусочек, чтобы понять, что это такое. После обработки окислителем и щелочью с кусочком произошло нечто странное. Он превратился в студенистую белесую массу, буквально набитую однородной пыльцой с двумя мешками и отчетливой ребристостью тела. Помимо массы пыльцы нашлись мелкие обрывки листовой кутикулы, такой же, как на листьях татарин. Дальше можно было не ломать голову над строением обугленной лепешки: это был копролит (окаменелый навоз) какого-то животного, объедавшего мужские органы размножения татарин и прихватывавшего листья (фиг. X).
Кто бы мог подумать, что животное, пасшееся в татариновых зарослях и оставившее аккуратную кучку, тем самым окажет услугу палеоботанике? Жаль только, что мало было этих животных и поэтому в наших коллекциях после тщательного их пересмотра нашлось лишь два других копролита, изучение которых еще впереди. Опущу дальнейшие перипетии исследования и сразу сделаю вывод: мы с А. В. Гоманьковым сейчас склонны считать, что растения с татариновыми листьями имели оба типа ребристой пыльцы, т. е. с мешками и без мешков, а также неребристую двухмешковую пыльцу.
Пыльца этих типов известна по всему разрезу перми и в небольшом количестве встречается в верхах карбона. Впрочем, из этого мы не заключаем, что пельтаспермовые появились в карбоне. Известно, что такую же пыльцу имели и другие растения, в том числе и некоторые гондванские глоссоптериевые. Там находили ребристую пыльцу с мешками, проникшую в семезачатки семян или еще не выпавшую из мужских спорангиев. Хитро все переплелось. Находим похожие побеги в разных местах, а потом оказываются разными органы размножения. Потом находим похожую пыльцу, и снова оказывается, что под одинаковой внешностью кроются разные растения. В который уже раз получают палеоботаники уроки осторожности.
В недостатке осторожности повинны все палеоботаники. Мне тоже случалось сильно обжигаться и попадаться на обманы внешнего сходства. Поэтому я не вправе первым бросать в кого-то камень. Если я сейчас выскажу упрек некоторым палинологам, то не потому, что сам безгрешен, а только потому, что этот упрек уже высказывали многие до меня,
Я имею в виду многочисленные указания в палинологической литературе на находки в палеозойских отложениях родов, характерных для мезозоя и даже кайнозоя. Ту самую ребристую безмешковую пыльцу, которую сейчас можно довольно уверенно связать с пельтаспермовыми, палинологи сравнивали с пыльцой современной группы покровосеменных, почти не известной широкой публике (знают только принадлежащую покровосеменным лекарственную эфедру). О хвойных я уже говорил. В перми Сибири находят пыльцу на первый взгляд того же типа, что и у мезозойских цикадовых, гинкговых и беннеттитовых. Соответственно в палинологических статьях порой пишут, что в перми Сибири жили эти группы растений, предшественники мезозоя. При этом делают сразу несколько ошибок. Во-первых, такую же пыльцу, как и у перечисленных групп, имели и пельтаспермовые птеридоспермы, которые тоже просятся в компанию. Во-вторых, пермская пыльца Сибири на самом деле совсем иная, чем у этих мезозойских групп.
Было показано, что за безмешковую пыльцу палинологи часто принимают оторвавшиеся и сложившиеся в лодочку мешки одномешковой пыльцы растений с листвой рода Rutloria. В такие же лодочки сворачиваются и оставшиеся без мешков тела. Наконец, та же пыльца, еще не созревшая, тоже складывается в лодочку. Три типа сохранности одной и той же пыльцы описываются как нечто совсем разное, да еще относятся к группам, характерным для мезозоя. В результате у мезозойской флоры отрастают ложные корни.
До сих пор речь шла только о Западной Ангариде как об источнике мезозойской флоры. Читатель может подумать, что это был какой-то сравнительно небольшой (в масштабе планеты) рассадник растений, которые при подходящих условиях расселились в мезозое и заполнили всю Землю. Едва ли дело происходило так, В литературе не раз выдвигались подобные точки зрения на происхождение и крупных систематических групп растений (например, покрытосеменных, о которых пойдет речь в следующей главе), и целых флор. В качестве главного источника флоры мезозоя выдвигались сибирская пермская флора и западноевропейская цехштейновая флора. Думаю, что на самом деле все было иначе. Каждая из флористических областей перми сделала больший или меньший вклад в растительный покров мезозоя. Кроме того, мезозойские группы растений, прежде чем осесть на своих местах, могли совершить далекие путешествия, основывая вторичные центры расселения.
Для реконструкции этого сложного процесса надо хорошо знать филогению основных мезозойских групп растений. Тут главный пробел в наших знаниях. В типично мезозойских флорах, населявших Землю от конца триаса примерно до середины мела, главную роль играют гинкговые, лептостробовые, или чекановскиевые (о них будет рассказано в следующей главе), цикадовые, беннеттиты, различные хвойные и папоротники, местами — птеридоспермы. Происхождение всех этих групп остается полностью неизвестным. Предполагают, что цикадовые произошли от каких-то палеозойских птеридоспермов, а гинкговые от кордаитов. Мезозойские хвойные филогенетически связывают с палеозойскими. Все эти и другие гипотезы пока подкрепляются не конкретными находками переходных форм, а некими абстрактными схемами, показывающими, как могли эволюционировать отдельные органы.
Может быть, в будущем мы будем полнее знать филогению основных мезозойских групп, а пока к проблеме происхождения мезозойской флоры приходится подходить иначе. Мы можем проанализировать, в каких пермских флорах больше всего представителей тех групп, которые особенно характерны для мезозоя, и каким образом соотносятся палеофлористические области перми и мезозоя.
При таком подходе картина получается следующей. Начнем с Ангариды. Типично ангарские пермские флоры, известные в Сибири и составленные преимущественно кордаитами и членистостебельными, едва ли сделали серьезный вклад в мезозойскую флору. Правда, здесь встречаются отдельные растения, которые сравнивают с мезозойскими родами, но в других местах Земли соответствующих растений гораздо больше. Иное дело — Западная Ангарида, о которой уже много говорилось. Эта территория лежала тогда в низких широтах, на краю тропической зоны, причем климат был примерно таким же, как в современном Средиземноморье, или более теплым. Судя по почти полному отсутствию углеобразования, в этих местах было довольно сухо. Неподалеку стояли Прауральские горы. Считается, что предгорные области с теплым и не слишком влажным климатом, большим разнообразием экологических условий особенно благоприятны для быстрой эволюции растений. Западная Ангарида хорошо отвечает перечисленным требованиям.
В последние годы накапливается все больше данных, что того же типа флора не только заселяла Приуралье и соседние с ним части Восточной Европы, но и уходила в Казахстан, Среднюю Азию и далее на восток, в Китай. Видимо, Ангарида была окружена поясом (назовем его Субангарской областью; рис. 20), в котором аборигены смешивались с пришельцами с севера и юга. На опубликованных в палеоботанической литературе палеофлористических картах эта область пока отсутствует, а очерчена лишь ее западная часть, получившая название Восточно-Европейской области (в поздней перми) и Уральско-Казахстанской области (во второй половине ранней перми).
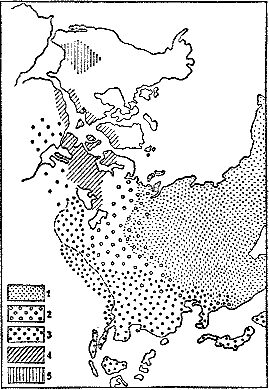
Рис. 20. Фитогеография середины перми (куньгурский, уфимский и казанский века): 1 — Ангарская область; 2 — Субангарская область (кружками показана и примесь субангарских растений в других флорах); 3 — Катазиатское царство; 4 — Атлантическое царство (включая цехштейновую флору); 5 — Северо-Американское царство
Гипотеза о существовании Субангарской области складывалась у меня постепенно из разрозненных наблюдений. В 1971 г. я был в Алма-Ате и смотрел коллекции палеоботаника К. 3. Сальменовой, собранные в Прибалхашье и Джунгарии. Хотя сохранность растительных остатков была довольно плохой (на растительных остатках нет угольной корочки, и поэтому нельзя приготовить препараты кутикулы), некоторые растения разительно напоминали приуральские. Возможность связи юго-востока Казахстана с Приуральем подтверждалась и некоторыми палинологическими данными по пермским отложениям района Джезказгана и Южного Казахстана (Чуйской впадины и Малого Каратау). Спорово-пыльцовые комплексы этих мест сходны с приуральскими. Затем ташкентский геолог А. С. Масумов прислал мне коллекцию, собранную в Чаткальском хребте. Снова обнаружились растения, до этого считавшиеся характерными для Среднего Приуралья. Потом К. 3. Сальменова показала мне прибалхашские отпечатки рода Phylladoderma, типичного для европейской части СССР. Этот же род В. И. Бураго нашла в верхней перми Приморья.
Неожиданной была коллекция из Афганистана, собранная душанбинским геологом В. И. Дроновым вблизи Кабула (фиг. XI). Здесь смешивались растения, характерные для верхней перми Поволжья и Прикамья, и явные пришельцы из Китая. Последнее меня не удивило, поскольку комплекс катазиатских растений был еще в 50-х годах найден английским палеоботаником Р. Вагнером в верхней перми на востоке Турции. Позже чехословацкий геолог П. Чтироки прислал мне фотографии растений, собранных им в Ираке и ошибочно отнесенных к гондванским родам и видам. Это тоже были катазиатские растения. Наконец, надо упомянуть коллекцию, собранную еще в начале 30-х годов шведским геологом Г. Бекселлом в хребте Наньшань восточнее коленообразной излучины Хуанхэ. Здесь выше отложений с катазиатской флорой были собраны растения, отнесенные шведским палеоботаником Т. Г. Галле к видам, характерным для перми Сибири. Из этого был сделан вывод, что вся сибирская ангарская флора моложе катазиатской. К коллекции Бекселла, оставшейся неописанной, никто не возвращался более 40 лет. Потом в Москве побывали шведские ученые Э. Норин и Б. Болин, которые пообещали прислать фотографии образцов Бекселла. Определения Галле оказались ошибочными, но дать новые по фотографиям было рискованно. Надо было изучать сами образцы. В 1977 г. М. В. Дуранте, до этого исследовавшая верхнепалеозойскую флору Монголии, поехала в Швецию. Ее работа продолжилась в Москве, куда была прислана часть образцов. Исследования коллекции Бекселла еще продолжаются, но уже можно сделать вывод, что и здесь есть немало растений, которые мы привыкли считать западно-ангарскими.
Точно оконтурить Субангарскую область пока невозможно. Трудно расшифровать ее соотношение с соседями как с севера, так и с юга. Пермская история районов с субангарской флорой крайне запутана. Ситуация осложнилась и тем, что в это время, примерно в начале поздней перми, катазиатские растения нашли дорогу на запад Евразии. Другие растения шли им навстречу (например, в верхней перми Дарваза найдены некоторые цехштейновые растения). С севера приходили сибирские растения, а с юга, видимо, гондванские. В верхнепермских отложениях Сибири и Приморья попадаются листья, очень похожие на глоссоптериевые. В некоторых местах получались удивительные смешанные флоры. Так, в палинологических комплексах верхней перми Соляного кряжа австралийский палинолог Б. Балм нашел странную смесь западноевропейских, приуральских и гондванских родов. Еще предстоит разобраться с соотношением субангарской и североамериканской флор. Судя опять же по палинологическим данным, между ними было немало общего.
Субангарская флора — неожиданно обнаружившееся белое пятно на палеофлористических картах перми. Его закрытие — одна из главных задач палеофлористики палеозоя на ближайшие годы. Не требуется пояснять, что одновременно удастся многое понять и в происхождении мезозойской флоры.
Южнее субангарской флоры располагались Катазиатская область (на востоке Евразии) и Атлантическая область (с флорой цехштейна). Обе они могли сделать некоторый вклад в мезозойскую флору. Для первой характерно большое количество растений с листьями, похожими на цикадовые. Может быть, здесь был центр происхождения цикадовых. В цехштейне много хвойных, и, хотя ни один род не перешел в триас, можно наметить филогенетические связи между цехштейновыми и триасовыми хвойными. Много хвойных и в Северной Америке.
Для понимания происхождения мезозойской флоры много могут дать детальное изучение и точная датировка растений Сибири, которые пришли здесь на смену кордаитам и пермским членистостебельным. Впервые эти растения были собраны в Кузбассе М. Ф. Нейбург и в Тунгусском бассейне Л. М. Шороховым еще в 30-х годах. Кузнецкие растения сразу отнесли к триасу, а дискуссия о возрасте тунгусских растений затянулась. Сначала их относили то к перми, то к триасу, а затем единодушно остановились на триасовом возрасте. Когда в 50-х годах началось детальное изучение отложений с тунгусскими растениями (из так называемой корвунчанской серии), выяснилось, что остатки животных (ракообразных, двустворчатых моллюсков) и спорово-пыльцевые комплексы в нижней части этих отложений скорее пермские, чем триасовые. Палеоботаник и стратиграф Г. Н. Садовников пришел к выводу, что корвунчанская серия состоит из двух частей. Нижняя относится к самым верхам перми, а верхняя — к триасу. Это решение кажется мне вполне разумным, хотя и нуждающимся в дополнительном обосновании. Многие палеоботаники придерживаются иного мнения. Они считают, что корвунчанская серия целиком принадлежит триасу. В подтверждение указывают на мезозойский облик корвунчанской флоры (фиг. XII), на резкую смену состава как самих отложений, так и растительных остатков на нижней границе корвунчанской серии.
Все это так. Действительно, вулканогенная корвунчанская серия резчайшим образом отличается от подстилающих угленосных отложений. Я согласен и с тем, что флористическая граница на этом рубеже тоже чрезвычайно резкая. Однако из этого никак не следует, что перестройки и в режиме осадконакопления, и в растительном покрове должны были произойти именно на рубеже перми и триаса. Они могли произойти и раньше, и позже. Мы должны сначала сопоставить разрезы Тунгусского бассейна с эталонными разрезами верхов перми и низов триаса, а уж потом решать, когда, где и что случилось.
Сопоставить разрезы можно по остаткам животных и растений. Животные довольно единодушно высказались в пользу пермского возраста нижней части корвунчанской серии. С растениями получилось сложнее. Правильно, что в корвунчанской серии есть мезозойские роды. Но будем точными. Эти роды хотя и мезозойские, но никогда не считались характерными для нижнего триаса. Они обычно считались верхнетриасовыми и даже юрскими. Корвунчанскую серию никто не относит к верхнему триасу или юре. Стало быть, все равно допускается, что некоторые роды спускаются в Сибири ниже по разрезу, чем в других местах Земли. Если же разрешается им спуститься до нижнего триаса, то почему надо закрывать им дорогу в пермь? При этом в эталонных разрезах верхов перми, т. е. в Западной Ангариде, облик флоры уже мезозойский. Здесь господствуют пельтаспермовые птеридоспермы — листья татарин, о которых много говорилось. Кстати, и в нижней части корвунчанской серии есть прослои с большим количеством татарин. В Западной Ангариде кордаиты полностью исчезают не на рубеже перми и триаса, а раньше. Поэтому связывать исчезновение кордаитов в Сибири с границей перми и триаса пока нет оснований.
Если возраст корвунчанской флоры действительно частично: пермский, то и эта флора может быть включена в число предшественников мезозоя. Впрочем, возможно, что Сибирь была не первичным, а вторичным центром расселения мезозойских растений, как бы временным лагерем в середине маршрута. На это предположение наводят, во-первых, все те же пельтаспермовые, которые появились в Субангарской области намного раньше, чем в Сибири, а во-вторых, почти полное отсутствие преемственности корвунчанской флоры и ее предшественницы в самой Сибири.
Я вообще склонен думать, что смена палеозойских флор на мезозойские происходила на фоне расширявшейся миграции растений, ранее сидевших по своим областям. Еще в середине перми началось оживленное движение по периферии Ангариды вдоль Субангарской области. В начале поздней перми тронулись в путь катазиатские растения, из Китая покатились на запад, добрались до Турции, Ирака и, видимо, пошли еще дальше. В самом конце перми (если раннекорвунчанская флора еще относится к перми) началось заселение Сибири новой флорой, пришедшей на смену кордаитам.
В триасе этот процесс продолжался. В раннем триасе чуть ли не по всему миру распространились своеобразные плауновидные плевромейи, сначала описанные в Европе, а потом в окрестностях Рыбинска, на Мангышлаке, в Приморье, на северо-востоке СССР. Недавно Г. Н. Садовников нашел огромные скопления их остатков на восточном Таймыре. Близкие растения найдены в Средней Сибири, Индии и Австралии. Где они появились, мы не знаем. Возможно, что их родиной была опять же Западная Ангарида, где в верхней перми встречаются похожие плауновидные.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Корни гипотезы
Корни гипотезы Пожалуй, только два факта (оба негативного свойства!) были положены Митчелом в основу его гипотезы в далеком уже 1961 году. Это невозможность найти химические продукты, которые были бы общими для дыхания и фосфорилирования, и необъяснимость роли мембран и
ГЛАВА XV.
ГЛАВА XV. Обработка земли под яровые хлеба.Обработку земли под яровые хлеба я начинаю тотчас после уборки озими . Только при соблюдении этого условия можно рассчитывать на самый обильный урожай.Поля, поросшие сорными травами и покрытые густым жнивьем, я вспахиваю
Глава I
Глава I Восемь лет назад я написал небольшую книгу «Аксиомы биологии»[1], в конце которой высказал предположение, что возможно создание общей теории эволюции последовательно реплицирующихся систем. Завершил я книгу словами: «Под эту категорию попадают не только объекты
КОРНИ И КРОНА
КОРНИ И КРОНА — Вот мы тут живем, — сказал Кролик очень медленно и громко, — все мы, и вдруг ни с того ни с сего мы однажды утром просыпаемся и что мы видим? Мы видим какое-то незнакомое животное! Животное, о котором никогда и не слыхивали раньше! — Какое насекомое? —
Глава 2. ДНК
Глава 2. ДНК На стене паба “Орел” в Кембридже висит синяя мемориальная доска, установленная в 2003 году в честь пятидесятилетия одного случая, когда разговоры в этом пабе приняли не совсем обычный оборот. Во время обеда 28 февраля 1953 года два завсегдатая “Орла”, Джеймс
ГЛАВА 5
ГЛАВА 5 С помощью Пам Смарт мне удалось разработать более простую и эффективную методику работы с людьми, испытывающими фантомные ощущения в отсутствующих конечностях, чем та, которая была описана в пятой главе.Мы провели серию опытов с людьми, у которых были
Глава 4 Животные корни религии
Глава 4 Животные корни религии Во многих религиях центральное место занимает не верование, а ритуальное поведение. Так, в иудаизме, например, от верующего требуется в первую очередь не знание догматов, а определенное, строго регламентированное поведение, соблюдение
Глава 8 Он и она
Глава 8 Он и она Равенство не означает одинаковость Мужчины и женщины ведут себя по-разному во многих ситуациях. В этом нет сомнения. Основной вопрос, который вызывает споры уже много тысяч лет: предопределены ли различия в их поведении природой или же они – результат
Глава 1. Вид
Глава 1. Вид ТЕМЫ• История эволюционных идей• Современное эволюционное учение• Происхождение жизни на Земле• Происхождение человекаВ настоящее время на нашей планете обитает несколько миллионов видов живых организмов, каждый из которых по своему уникален. Каким
Глава 10. Уши{10}
Глава 10. Уши{10} Того, кто заглянет поглубже в ухо, чтобы увидеть, как устроен наш орган слуха, ждет разочарование. Самые интересные структуры этого аппарата скрыты глубоко внутри черепа, за костяной стенкой. Добраться до этих структур можно только вскрыв череп, удалив мозг,
Эволюционные корни ритуала
Эволюционные корни ритуала Антропологи уже давно пришли к выводу, что ритуалы в древнейших человеческих сообществах способствовали выживанию, поддерживая в данном клане или племени чувство особой идентичности и общей судьбы[91]. Ритуал постоянно напоминал членам