Глава VIII «ОТВРАТИТЕЛЬНАЯ ТАЙНА»
Глава VIII
«ОТВРАТИТЕЛЬНАЯ ТАЙНА»

Я не знаю, все ли области науки доступны для популяризации. Наверное, не все. Иногда мне кажется, что и популяризация палеоботаники — дело безнадежное. О чем бы ни повел палеоботаник речь, ему придется так или иначе употреблять множество слов, незнакомых широкой публике. Чем-то их заменить невозможно, а без них совершенно немыслимо рассказать о самом важном и действительно интересном. Короче говоря, чтобы читать популярную книгу по палеоботанике, надо освоить некий новый язык со словами «спорангий», «птеридоспермы», «карбон», «Гондвана». У кого хватит терпения все это держать в голове?
Все же я взялся за эту книгу и вот подбираюсь к ее концу. Меня ободряет то, что среди ее читателей будет достаточное количество географов, геологов и ботаников, а среди них и те, кто интересуется ископаемыми растениями, но не в силах следить за огромной палеоботанической литературой. Полагаю, что та глава геологической летописи, о которой сейчас пойдет речь, интересует ботаников больше всего. Я собираюсь говорить о происхождении и расселении покрытосеменных, или цветковых.
Для человека, неплохо знакомого с палеоботаническими документами, происхождение покрытосеменных не составляет какой-то экстраординарной проблемы. Да, мы не знаем их предков и не знаем, как они возникли. Что ж из того, что в этом особенного? Много ли крупных групп растений, происхождение которых известно? Если отвечать честно и не выдавать шаткие и валкие домыслы за хорошо обоснованные гипотезы, то таких групп не окажется вовсе. Что мы знаем о происхождении цикадовых, гинкговых, хвойных (и вообще голосеменных), папоротников, членистостебельных, мхов, разных групп водорослей? А почти ничего не знаем.
И все же ситуация с покрытосеменными несколько особая. Это самая крупная группа современных растений. О значении покрытосеменных для человечества нет нужды говорить. Покрытосеменные — это хлеб, овощи, фрукты, топливо, строительная древесина, цветы на окнах и в цветочных магазинах, лекарства в аптеках. Классификация покрытосеменных — не отвлеченная научная проблема. Разобраться же в предложенных классификациях и отобрать наиболее удачную и практически наиболее удобную без понимания эволюции всей группы почти невозможно.
Поразительно разнообразие покрытосеменных. Достаточно вспомнить ряску и дуб, пшеницу и кактус. Много ли общего между прекрасным цветком магнолии и скромным ржаным колосом? Откуда взялось это невероятное разнообразие, как понять его? Все это волнует ботаников уже несколько столетий. С тех пор как в ботанику и биологию вообще проник исторический метод исследования, возникла, да так и не исчезла проблема происхождения покрытосеменных. Быстрое появление покрытосеменных в недавние геологические эпохи еще Ч. Дарвин назвал «отвратительной тайной» — abominable mistery. Замечу, что прилагательное abominable в толковом словаре еще поясняется словами «противная» и «ненавистная».
Ситуация и в самом деле какая-то противная. Представим себе, что некто хвалится познаниями в радиотехнике. Его просят починить что-то совсем обычное — радиоприемник или телевизор, и оказывается, что в них-то он ничего не понимает. С историческим методом в ботанике получается примерно то же. Сказано немало высокопарных слов об огромном значении историзма в систематике и морфологии растений, но мы все еще бессильны расшифровать происхождение и эволюцию самых распространенных высших растений.
Впрочем, словам Дарвина уже сто лет (он их написал в письме ботанику Дж. Д. Гукеру в 1879 г.). Может быть, с тех пор все изменилось? Многие ботаники оптимистически скажут, что изменилось немало и что мы сейчас гораздо лучше представляем себе и предков, и эволюционные пути покрытосеменных. Кто-то, может быть, расскажет про новейшие гипотезы. Некоторым из них посвящены толстенные книги. Кто-то напомнит об огромной литературе из тысяч статей и монографий на эту тему. Если продуктивность научной работы исчислять количеством и объемом опубликованных работ, то прогресс действительно будет потрясающим. Однако потрясение от прогресса мгновенно улетучится, как только мы начнем знакомиться с выводами, к которым приходят исследователи. Разнобой взглядов потрясет читателя гораздо больше, чем любой из выводов.
Я не собираюсь утверждать, что мы так и остались на уровне незнания столетней давности. Наши знания с тех пор изрядно увеличились. Ботаники несравненно лучше знают теперь строение и функционирование покрытосеменных, их образ жизни и распределение по лику планеты. Выдвинуты некоторые весьма правдоподобные гипотезы о том, какие могли существовать филогенетические отношения между отдельными родами, семействами и порядками. Что же касается самого важного — исторического отношения крупных групп покрытосеменных, то здесь прогресс ничтожен. Активность исследователей в отношении всех важнейших вопросов больше напоминает не организованное шествие, а толчею в магазине. К какому-то прилавку вдруг выстраивается огромная очередь. Выстояв несколько часов за дефицитным товаром, люди с удивлением узнают, что стояли зря: товара нет и не будет, а продают какую-то чепуху. В начале 50-х годов были описаны органы размножения глоссоптерид из гондванской флоры Южной Африки. Прямо в основании листа на ножке сидят овальные органы, в которых, как показалось сначала, мужские спорангии сочетаются с семенами. Обоеполые цветки — характернейшая черта покрытосеменных. Органы размножения глоссоптерид — тоже обоеполые, что большая редкость среди голосеменных. Читатель, может быть, помнит, что глоссоптериевые листья имеют сетчатое жилкование, и в этом отношении они, стало быть, тоже похожи на листья покрытосеменных. Родилась надежда, что именно гондванские глоссоптериевые — то недостающее звено в филогении, которое связывает голосеменных с покрытосеменными.
Статья была опубликована в одном из наиболее авторитетных палеонтологических журналов. Главный редактор журнала понимал возможный резонанс сообщения. Поэтому рукопись статьи была показана нескольким видным палеоботаникам. Их комментарии были опубликованы вместе со статьей. Мнение было, почти единодушным: доказательств, что у глоссоптерид действительно были обоеполые органы размножения, не представлено. Материал, который описан и изображен, слишком плохой сохранности. Общая структура органов размножения непонятна. Особенно сомнительны те структуры, которые были приняты за мужские спорангии. Словом, критика была не просто резкой, а почти уничтожающей и… бесполезной. Доверчивых и не слишком внимательных читателей оказалось немало. Гипотетические интерпретации стали широко известны, а предостережения палеоботаников прошли незамеченными. В ботанической литературе появились статьи, где глоссоптериды прямо назывались предками покрытосеменных, а Гондвана — их родиной.
Поразительные вещи происходят порой в науке. Я готов всерьез отстаивать взгляд, что механизм возникновения и распространения многих научных убеждений не многим отличается от того, что происходит со сплетнями. Кто-то что-то видел или читал, толком не разобрался, заявил во всеуслышание. Ему сразу возразили, все сказанное убедительно опровергли, а присутствовавшие чего-то недослышали. Вот и поползли пустые и досадные разговоры.
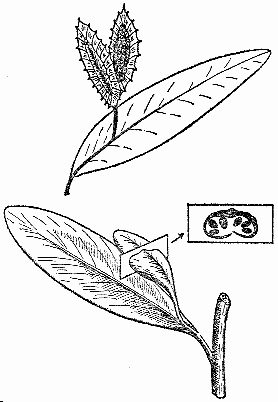
Рис. 21. Глоссоптериевый женский орган размножения, как его представила Э. Пламстед (наверху), по современной реконструкции Т. Делевориаса и Р. Гулда (внизу)
Я не хочу винить кого-то в публикации статьи. В общем была соблюдена осторожность и в выводах, и в частных интерпретациях образцов. Некоторые наблюдения были действительно интересными. Например, было очевидно, что найдены какие-то органы размножения, и они в самом деле прикреплялись к основанию листа. На некоторых образцах были хорошо видны семена. Ограничиться бы этими сообщениями, и все было бы хорошо. Впрочем, теперь поздно давать советы «задним числом».
За прошедшие с тех пор годы число находок глоссоптериевых органов размножения сильно увеличилось (рис. 21). Недавно нашли их полностью минерализованными, так что можно рассмотреть каждую клетку. От прежних интерпретаций давно ничего не осталось. Никаких обоеполых органов размножения у глоссоптериевых не было. Мы знаем, какие у них были семена и мужские спорангии и как они сидели. На сплетню, которая давно гуляет по свету, все это не повлияло.
В статье, вышедшей в 70-х годах, когда ошибка была давно ясна, читаем: «Glossopteridales, по мнению всех исследователей, хотя и не являются прямыми предками покрытосеменных или, во всяком случае, не всех покрытосеменных, но очень близки к ним». В этой цитате выражение «по мнению всех исследователей» подобно словам «все говорят» в каждой порядочной сплетне.
Проблема происхождения покрытосеменных знает немало слухов. В 50-х годах появилось сообщение, что в каменноугольных отложениях нашли пыльцу покрытосеменных. Это были любопытные оболочки почти квадратного очертания с ясными порами по углам. Сходство с пыльцой действительно было. Палинологи быстро выпустили воздух из раздувшейся сенсации. Оказалось, что эти оболочки принадлежат каким-то морским водорослям. И все же несколько лет назад один видный ботаник подошел ко мне во время перерыва совещания и спросил, какие есть новости о пыльце покрытосеменных в карбоне. Слух оказался живучим.
В 1955 г. М. Ф. Нейбург описала странное растение, найденное Е. С. Рассказовой в пермских отложениях Печорского бассейна (фиг. XIII, XIV). На оси сидят мясистые конусовидные органы, несущие на конце щетку булавовидных придатков. Между придатками Нейбург нашла еще не опавшее семя. Видимо, статья Пламстед, вышедшая незадолго до этого, повлияла на интерпретацию печорского растения (Нейбург назвала его «войновския»). Почему-то Нейбург решила, что булавовидные придатки — мужские спорангии, а значит, войновския — обоеполый орган размножения. И опять в литературе заговорили о том, что найден возможный предок покрытосеменных.
Я принимал на веру вывод Нейбург о строении войновскии, пока известный английский палеоботаник Т. М. Гаррис не стал меня с пристрастием допрашивать, какие есть доказательства, что булавовидные придатки действительно мужские спорангии. Я ничего не мог ответить. Просто так считала Нейбург. Все же пришлось войновскией заняться более обстоятельно. Оказалось, что Гаррис спрашивал не зря. В булавовидных придатках ничего общего со спорангиями не оказалось. Это были всего лишь видоизмененные чешуи, сидевшие между семенами. Такие чешуи известны у многих голосеменных.
Подобные совершенно явные недоразумения составляют значительную, но не львиную долю литературы по происхождению и расселению покрытосеменных. Больше всего в ней работ, где исследователи пытаются понять случившееся в далеком прошлом, глядя на современный растительный мир. Они изучают современные цветки и пытаются понять, как эволюционировали их части, как произошли плодолистики, тычинки, венчик. Наблюдая современные жизненные формы, они спорят о том, с деревьев или трав началась эволюция покрытосеменных. Обсуждается, какими были листья первых покрытосеменных — цельными, как у лавра, или расчлененными, как у винограда. По тому, как расселены ныне по поверхности Земли разные группы покрытосеменных, ботаники пытаются расшифровать их центры происхождения и пути расселения.
Палеоботанические материалы в этой литературе фигурируют лишь от случая к случаю. Пусть не сердятся на меня ботаники, если я скажу, что поверить в выводы, которые они делают, в большинстве случаев невозможно. Причина моего неверия очень простая. Как правило, выводы о филогении, сделанные по разным признакам, вступают в противоречие. Скажем, среди ботаников популярна идея, что травянистые формы происходят от древесных, что цветки с большим числом плодолистиков, тычинок и лепестков дали начало цветкам с малым числом частей, что пыльца с тремя бороздами примитивнее многобороздной и т. д. Если попытаться расположить в филогенетической последовательности конкретные роды или семейства, то окажется, что разные свидетели примитивности и прогрессивности дают несогласованные показания. Складывающиеся ряды форм можно «читать» филогенетически по-разному. Такого, чтобы показания всех признаков совпали, не случается почти никогда. (Слово «почти» я вставил из осторожности. Думаю, что без него можно и обойтись.)
В новейших сводках по филогении покрытосеменных самыми примитивными считают растения, приближающиеся по общему облику к современным магнолиевым. Порядок магнолиевых стоит у корня филогенетического древа, ветви которого составляют все прочие порядки числом около сотни. Покопавшись в литературе, можно найти и другие мнения. Некоторые ботаники считают, что из магнолиевых нельзя вывести однодольных, что крупные одиночные цветки, не собранные в соцветия, признак не примитивности, а прогрессивности. Указывают, что по биохимическим признакам магнолиевые — весьма высокоразвитая, а отнюдь не примитивная группа. Правда, другие биохимики с этим не соглашаются.
Что делать при таких разнотолкованиях? Как убедиться, кто прав и кто не прав? Ботаники ищут выхода из противоречий в расширении круга анализируемых признаков. К сожалению, увеличение числа признаков ведет лишь к умножению противоречий. Новые признаки не согласуются друг с другом и со старыми. Я не буду вдаваться в теоретические рассуждения, как быть в подобных случаях, как разрешать филогенетические противоречия. Кое-что на этот счет сказано в следующей главе. Сейчас лишь замечу, что решение филогенетических проблем без обращения к палеонтологической летописи, по-видимому, принципиально невозможно.
В ботанической литературе широко распространено мнение о чрезвычайной неполноте палеонтологической летописи. Поэтому, дескать, ископаемые растения — разрозненные и мало о чем говорящие документы. В одной из сводок по эволюции и классификации покрытосеменных так и написано, что геологическая летопись, польза которой «весьма ограничена для прояснения родственных отношений между разными группами покрытосеменных, не более полезна в выявлении их предка».
В тех палеоботанических примерах, с которыми мы уже познакомились в этой главе, роль палеоботаники действительно довольно неприглядна. Скорее она была источником путаницы, а не знания. К счастью, не все палеоботанические материалы того же достоинства. Палеоботаника сейчас располагает очень интересными фактами, которые, правда, не всегда известны ботаникам. Особенно важные сведения были получены в последние 10–15 лет. Почему-то скорость их распространения куда более скромна, чем скорость сообщений об обоеполых органах размножения глоссоптериевых и войновский.
Думаю, что причина этого прежде всего в том, что новые данные плохо согласуются с наиболее признанными убеждениями. Они заставляют пересмотреть всю систему взглядов на происхождение и расселение покрытосеменных, Я недаром выделил разрядкой слово «система». Проблема происхождения и расселения покрытосеменных — сложное событие. Для его реконструкции надо учитывать широкий круг фактов, доставляемых не только ботаникой и палеоботаникой, но и науками о Земле — стратиграфией, палеогеографией, тектоникой, геохимией и др. Пренебрежение этими фактами может направить все исследование по неверному руслу.
В 1927 г. в Москве вышла книга М. И. Голенкина «Победители в борьбе за существование». Потом она дважды переиздавалась, что редко случается в научной литературе. В этой книге большое внимание было уделено распространению покрытосеменных в середине мелового периода. Еще в прошлом веке было замечено, что количество листьев покрытосеменных в верхнемеловых отложениях значительно больше, чем в нижнемеловых. Некоторые верхнемеловые захоронения растений иногда почти лишены других растительных остатков, а отпечатки покрытосеменных образуют листовые кровли. В нижнемеловых отложениях эти отпечатки встречаются, но гораздо реже. Внезапное увеличение остатков покрытосеменных в середине мела и породило ту «отвратительную тайну», о которой писал Дарвин в письме Гукеру.
В гипотезах, объяснявших этот факт, никогда не было недостатка. Большей частью они исчезали столь же незаметно, как и появлялись. Гипотезе Голенкина повезло гораздо больше. Ее сразу заметили, и ее сторонников немало до сих пор. Сочувственное изложение идей Голенкина можно найти даже в учебниках.
Голенкин предположил, что покрытосеменные стали «победителями в борьбе за существование» по нескольким причинам, как зависящим, так и не зависящим от них самих. Одним из решающих преимуществ покрытосеменных перед голосеменными и споровыми, населявшими сушу до позднего мела, Голенкин считал способность выносить яркий солнечный свет и сухость воздуха. Эти преимущества не имели большого значения до определенного момента. Раз покрытосеменные появились еще в раннем мелу и особенно не выделялись, а потом внезапно распространились по всей Земле, значит, им что-то помогло. Не могли же частные и местные факторы сработать сильно и одновременно по всей Земле!
Опираясь на некоторые косвенные соображения, Голенкин предположил, что до середины мела Земля была укутана плотными облаками, так что солнечные лучи редко пробивались к земной поверхности. В этой обстановке и появились первые покрытосеменные. Тогда они не имели серьезных преимуществ перед тенелюбивыми и влаголюбивыми голосеменными и споровыми растениями. В середине мела по каким-то космическим причинам облачный покров планеты сильно поредел. Яркие солнечные лучи хлынули на Землю. Вместе с освещенностью возросла и сухость климата. Большинство споровых и голосеменных растений, господствовавших до этого, не смогли приспособиться к революции климата и света. Они вымерли и освободили жизненное пространство покрытосеменным, «детям солнца».
Я не проводил специальных изысканий, чтобы выяснить, насколько заинтересовала гипотеза Голенкина палеоботаников. Может быть, кто-нибудь и высказывался о ней с сочувствием. Во всяком случае это не было правилом. Показательно, что в капитальной сводке А. Н. Криштофовича «Палеоботаника», вышедшей посмертным изданием в 1957 г., книга Голенкина упоминается в списке литературы, а в главе о меловых флорах о его гипотезе не говорится ни слова. Разумеется, отсутствие ссылок можно объяснять по-разному. Бывает ведь и заговор молчания против выдающейся идеи. Я с трудом верю, что в случае гипотезы Голенкина было умышленное игнорирование. Просто она не заинтересовала профессионалов-палеоботаников. О ней все знают и остаются равнодушными. Еще меньше ее популярность за рубежом.
Популярность гипотезы Голенкина поддерживается преимущественно в ботанических кругах, но я ни разу не видел, чтобы ее анализ сопровождался внимательным отношением к материалам исторической геологии. Рассматриваются покрытосеменные вообще, говорится о меловом периоде в масштабе всей Земли. Нет ни рассмотрения конкретных, известных палеоботаникам меловых флор, ничего не говорится об условиях их существования, как они видятся специалистам.
При том равнодушии, которое выказывают палеоботаники к гипотезе Голенкина, может быть, и неуместно много о ней говорить. Все же я решил разобрать ее более подробно, поскольку она показательна с методологической точки зрения. При этом я вовсе не ставлю в вину Голенкину незнание фактов, ставших известными позднее. Такого упрека заслуживают лишь те, кто поддерживает идею Голенкина до сих пор.
Главный дефект гипотезы Голенкина в том, что она приписывает всему миру то, что характерно только для Европы. За ее пределами события в середине мела выглядят иначе. В некоторых местах листовые кровли с покрытосеменными встречаются в верхних горизонтах нижнего мела. В других местах Земли, наоборот, покрытосеменные преобладают в захоронениях только в верхних горизонтах верхнего мела, а до этого главную роль играют хвойные.
Сейчас, когда стратиграфия меловых отложений стала достаточно детальной и поэтому датировки толщ с растительными остатками приобрели определенность, картина расселения покрытосеменных представилась совсем иной, чем в 20-х годах. Если опираться не только на отпечатки листьев, но и на остатки пыльцы покрытосеменных, то картина получится еще менее драматичной. В некоторых районах количество покрытосеменных действительно резко увеличивается на определенном уровне, но в разных районах соответствующие рубежи оказываются приуроченными к разным интервалам мелового периода.
Судя по длительности эпох мелового периода, процесс завоевания суши покрытосеменными затянулся на многие миллионы лет. Не будем забывать и о том, что до сих пор самые распространенные деревья на Земле — сосна и ель, т. е. голосеменные, а не покрытосеменные растения. Таким образом, разрушен главный компонент прежних взглядов — внезапность и одновременность победы покрытосеменных по всей Земле. Даже в масштабах геологического времени его нельзя считать быстрым. Палеоботаники давно отметили, что флоры в каждом регионе меняются как бы толчками. На большом участке геологического разреза мы наблюдаем примерно одного типа флору, затем на коротком отрезке состав флоры меняется, и снова наступает период спокойствия. Масштабы перемен могут быть различными. Иногда флоры меняются быстро и почти целиком, так что между старой и новой флорами не остается общих видов. В этом отношении флористические границы внутри мела, на которых в том или ином месте Земли покрытосеменные вытеснили предшественников, ничем не отличаются от прочих флористических границ. С некоторыми мы сталкивались в прошлых главах.
Поставив меловые события в один ряд с другими флористическими событиями в истории Земли, мы должны, как мне кажется, не подбирать каждой границе в каждом районе особое объяснение и уж тем более не спешить за помощью к космическим причинам, а попытаться понять общие причины, приводящие к сменам флор. Думаю, что в случае расселения покрытосеменных сработали обычные земные механизмы: перестройки барьеров, смена миграционных маршрутов, климатические перемены и пр.
Меловой период богат крупными событиями в истории лика Земли. Еще в конце юры произошел раскол Гондваны на две части. Западная Гондвана, охватывающая Африку и Южную Америку, отделилась от Восточной Гондваны (Индии, Австралии и Антарктиды). Примерно на рубеже юры и мела, как полагают некоторые тектонисты, Индия стала отходить от Антарктиды. В первой половине мела распад Гондваны продолжался, Очередь дошла до Западной Гондваны, которая стала делиться на Южную Америку и Африку (рис. 22). Этот процесс подробно прослежен по меловым отложениям, сохранившимся в приатлантических частях обоих материков (об этом рассказывалось в главе VI). В позднем мелу образовался Протоатлантический океан. Чем дальше, тем больше хирел великий средиземноморский океан Тетис. Индия отделилась от Мадагаскара и отправилась на север, чтобы войти в новый материк — Евразию. В конце мела Индия уже лежала в тропиках. Антарктида, наоборот, поехала на юг, и ее климат становился все холоднее.
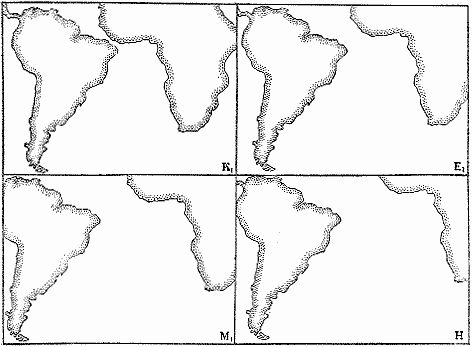
Рис. 22. Взаимное положение Южной Америки и Африки в раннем мелу (К), середине палеогена (раннем эоцене — EI), начале неогена (раннем миоцене — Ml) и сейчас (в голоцене — Н) по реконструкции Т. ван Анделя, Д. Тиде, Д. Г. Скле Итеру и В. В. Хэю
Может быть, я слишком уверенно расписываю все эти события. В истории гондванских материков и в маршрутах их путешествий (если вообще их допускать) еще немало неясного. Пусть не описанные события, а другие, тоже достаточно крупные, все же так или иначе происходили на Земле в меловое время. С тем, что в мелу происходили серьезные палеогеографические перестройки, никто спорить не будет. И при фиксистском, и при мобилистском прочтении истории приходится допускать, что в это время перестраивалась вся система циркуляции водных, а с ними и атмосферных масс. Должны были меняться и ландшафты, особенно если материкам приходилось ползти поперек параллелей. Ломались зоны растительности, а с ними и экосистемы.
Все эти события отнюдь не были катастрофическими, хотя в отдельных регионах перестройки совершались быстро. Желающие могут пофантазировать, как в обстановке смуты и ломки экосистем покрытосеменные подняли головы, стали одолевать соперников — споровых и голосеменных — и вытесняли их из ландшафтов.
Я не берусь сам рассуждать на эти темы, поскольку мы знаем еще слишком мало. И вообще сейчас здесь надо поменьше рассуждать, а более тщательно собирать и анализировать сохранившиеся документы. Сведения об ископаемых флорах по всем видам растительных остатков, особенно по пыльце и спорам, надо связать с конкретным палеогеографическим фоном, составлять подробные палеофлористические карты, много карт, для всех районов, где сохранились подходящие отложения. Такие исследования ведутся, хотя и в ограниченном масштабе. До последнего времени большая часть карт использовала фиксистские реконструкции палеогеографического фона. Теперь надо на все посмотреть и с мобилистских позиций. В отношении покрытосеменных такую попытку недавно сделали американские исследователи ботаник Р. Рейвн и палеоботаник Д. Аксельрод (он, если помнит читатель, когда-то резко протестовал против мобилистских идей).
Задача реконструкции меловых событий в растительном покрове Земли осложняется тем, что приходится опираться на палеогеографические реконструкции, которые сами зависят от интерпретации палеоботанических данных. Достоверность схемы расположения континентов подлежит палеоботаническому экзамену. Геологические и палеоботанические (и вообще палеонтологические) данные еще предстоит свести в единую непротиворечивую систему. Только тогда можно будет более уверенно судить о том, где были центры формирования отдельных групп покрытосеменных, как и когда шло их расселение. Не выполнив этой работы хотя бы начерно и не построив палеофлористических карт всего мира хотя бы для каждого века мелового периода (таких карт пока нет), нет смысла увлекаться гипотезами о конкретных причинах того, что произошло в растительном покрове Земли в те далекие времена.
Гипотеза Голенкина — не единственная, которой не было суждено выдержать проверку фактами. Новые материалы заставляют усомниться и в других гипотезах, особенно касающихся места и времени возникновения покрытосеменных. Зашатались и гипотезы об облике первых покрытосеменных. Я не хочу сказать, что есть ответы на все эти вопросы. Можно говорить лишь о сокращении числа возможных вариантов ответа на них.
Я уже говорил о том, что многие ботаники пытались найти ответы чуть ли не на все вопросы, глядя на современный растительный мир, и что исходной формой покрытосеменных чаще всего считают древесные растения с крупными одиночными цветками, напоминающими цветки магнолий. Построив на этой основе предполагаемую филогению покрытосеменных, ботаники попытались отыскать и родину этих растений. Оказалось, что семейства и роды, стоящие в нижней части предполагаемого филогенетического древа, не расселены равномерно по всей Земле, а сконцентрированы в отдельных районах. Самое большое их скопление отмечено в Юго-Восточной Азии. Ее-то и нарекли предполагаемой родиной покрытосеменных.
До последнего времени эти взгляды нельзя было непосредственно проверить по палеоботаническим данным. К тому же палеоботаников и ботаников долго сбивали с толку остатки якобы покрытосеменных, которые указывались в юрских и даже триасовых отложениях. Скажем, в юрских отложениях описали пыльцу с тремя бороздами, сходную с пыльцой современной эвкоммии. В триасе нашли отпечатки, сильно напоминающие листья однодольных. Теперь большая часть этих остатков предполагавшихся покрытосеменных из домеловых отложений изучена вновь, и их принадлежность покрытосеменным признана или ошибочной, или очень сомнительной.
С другой стороны, в нескольких достаточно полных разрезах меловых отложений тщательно изучены остатки листьев и пыльцы, принадлежность которых к покрытосеменным достаточно определенна. Об этих работах стоит кратко рассказать. Им посвятили свои статьи палеоботаник Л. Д. Хики и палинологи Д. Дойл, М. Ван Кампо, Б. Люгардон и некоторые другие.
Главная особенность проведенного ими исследования в том, что они широко применили электронно-микроскопическую технику. Еще в прошлом веке, когда в распоряжении ботаников появились неплохие световые микроскопы, было обнаружено, что пыльца растений имеет многослойную оболочку. У разных растений структура слоев оказалась неодинаковой. В последние десятилетия для изучения пыльцы сначала современных, а затем и ископаемых растений стали использовать электронный микроскоп.
Из пыльцевого зерна приготавливают тончайшие срезы, которые потом изучают при огромных увеличениях. Для исследования поверхности пыльцевого зерна на нее напыляют в вакууме тончайший слой золота или другого металла и помещают зерно под сканирующий электронный микроскоп. Максимальное увеличение, применявшееся при изучении пыльцы, достигало миллиона раз. Как отмечал шведский палинолог Г. Эрдтман, который одним из первых применил электронную микроскопию для изучения спор и пыльцы, при таких огромных увеличениях пыльцевое зерно липы, имеющее в диаметре около 40 микрон, увеличивается до 40 м. По нему можно было бы совершать экскурсии. Правда, на практике такими увеличениями не пользуются. Обычно ограничиваются увеличениями в несколько тысяч или в первые десятки тысяч раз. Но и при увеличении в 10 тыс. раз диаметр пыльцы липы будет 40 см, т. е. больше футбольного мяча.
Пыльца одних растений имеет достаточно сложное строение, замысловатый рельеф. Различить разные группы растений по такой пыльце можно и в световой микроскоп. У других растений пыльца гораздо проще. Например, пыльца цикадовых, гинкговых, беннеттитов, некоторых птеридоспермов и покрытосеменных может выглядеть в световой микроскоп почти одинаково — в виде маленькой булочки без заметного рельефа. В этом случае без электронной микроскопии обойтись нельзя. Сделав срезы с пыльцевых зерен и рассмотрев тонкую структуру их оболочек, можно отделить пыльцу покрытосеменных от голосеменных и различить разные группы голосеменных (рис. 23). Именно так и изучили ту пыльцу из юрских и нижнемеловых отложений, которую предположительно считали принадлежащей покрытосеменным.
Оказалось, что первая достоверная пыльца покрытосеменных появляется в нижней половине нижнемеловых отложений. Почти на том же уровне в разрезе появляются и первые достоверные остатки листьев покрытосеменных. Разнообразие листьев и пыльцы довольно быстро увеличивается вверх по разрезу. Д. Доил и Л. Хики проследили этот процесс в нижнемеловых отложениях штатов Виргиния, Мэриленд и Делавэр и составили выразительную схему, которую я воспроизвожу (рис. 24).
Примерно такую же последовательность форм палеоботаники наблюдали и в других местах Земли, хотя в разных местах первую пыльцу покрытосеменных находят не на одном стратиграфическом уровне. Раньше всего она появляется в районах, прилежащих к нынешней Атлантике, — в Англии, на западе Экваториальной Африки. В Австралии это произошло с запозданием примерно в 10 млн. лет. Сравнительно поздно она появляется и в геологических разрезах Юго-Восточной Азии. Таким образом, палинологические данные никак не подтверждают идею о первом появлении покрытосеменных в Юго-Восточной Азии. Более вероятно их африканское происхождение, поскольку именно в Африке, если верить палинологическим данным, быстрее всего и раньше, чем в других местах, нарастало разнообразие покрытосеменных (рис. 25). Именно здесь они впервые становятся заметным компонентом растительности. В отложениях аптского века (второго от конца мелового периода) доля пыльцы покрытосеменных в спорово-пыльцевом комплексе достигает здесь 5 %. В других местах такого процента покрытосеменные достигают позже. Впрочем, данные на этот счет еще предстоит свести воедино и как следует проанализировать.
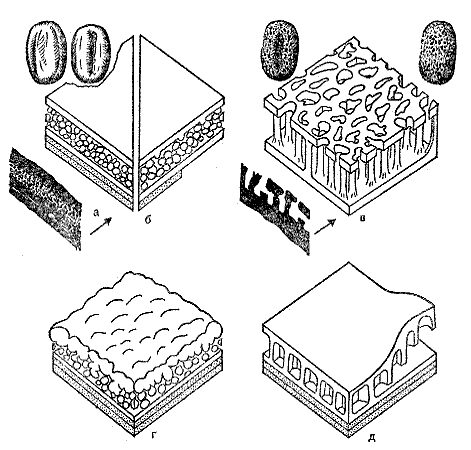
Рис. 23. Структура оболочки спор и пыльцы у разных растений — прогимноспермов (предголосеменных), голосеменных и покрытосеменных, по данным электронной и световой микроскопии; а — современные гнетовые, юрский Eucommiidites (слева вверху — общий вид пыльцевого зерна Eucommiidites с двух сторон; слева внизу — вид его оболочки в разрезе в электронный микроскоп); б — некоторые сережкоцветные, розоцветные, сложноцветные и другие покрытосеменные; в — многие покрытосеменные, в том числе древнейшие (в углах наверху показан общий вид пыльцевого зерна нижнемелового Clavatipolienites с двух сторон; слева внизу — вид его оболочки в разрезе в электронный микроскоп); г — прогимноспермы, кипарисовые, араукариевые и другие голосеменные; д — цикадовые (левая часть рисунка), птеридоспермы, кордаитовые, сосновые, подокарповые (по Д. Дойлу, М. ван Кампо и В. Люгардону)
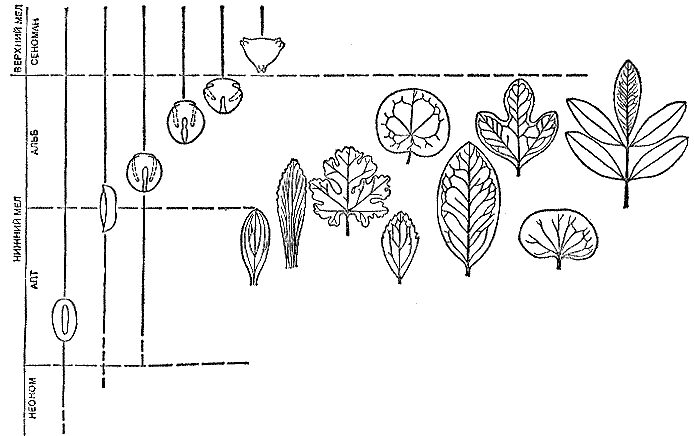
Рис. 24. Распространение в меловых отложениях Америки, Африки и Западной Европы основных типов пыльцы и листьев покрытосеменных; во второй колонке слева — ярусы (века) мелового периода (по Д. Дойлу и Л. Д. Хики с дополнениями)
Сейчас пыльца меловых покрытосеменных интенсивно изучается в разных странах, и, надо полагать, нынешние представления о ее распространении будут существенно меняться. Однако едва ли мы когда-нибудь вернемся к гипотезе, что их колыбелью была Юго-Восточная Азия. Дело в том, что в течение мела и большей части палеогена Юго-Восточная Азия входила в единый палеогеографический пояс, примыкавший к океану Тетис (или его остаткам) и протягивавшийся далеко на восток вплоть до Западной Европы. Еще в 1968 г. голландский палинолог И. Муллер обнаружил, что палинологические комплексы палеогена (точнее, эоцена) Калимантана и Западной Европы очень сходны. Уже из этого можно заключить, что нынешняя Юго-Восточная Азия в ботаническом отношении — остаток обширной территории, когда-то заселенной примерно однотипной растительностью. Теперь она скорее дом для престарелых, убежище для тех, кого климатические перемены выгнали из других мест.
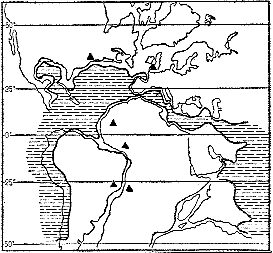
Рис. 25. Места находок древнейшей (доальбской) раннемеловой пыльцы покрытосеменных; жирной линией показана древняя береговая линия
В последние годы тщательно изучались и листья древнейших покрытосеменных из нижнемеловых отложений. Если не обращать внимания на тонкие особенности жилкования, то эти листья можно сблизить с современными семействами и даже родами. Палеоботаники долго так и поступали. Кажущаяся близость раннемеловых и современных покрытосеменных была одной из причин существовавшего убеждения, что раннемеловая летопись этих растений свидетельствует лишь об их расселении, а не о первых шагах эволюции. Собственно происхождение покрытосеменных относили к юре, триасу или даже перми.
Как мы уже видели, палинологические материалы не поддержали этой гипотезы. Внимательное изучение отпечатков листьев подтвердило палинологические наблюдения. Оказалось, что если принять во внимание тонкие детали жилкования, то уже нельзя сблизить раннемеловые листья не только с родами, но и с семействами современных растений. Это что-то совсем иное, а сходство с современными родами и семействами чисто внешнее. Таким образом, более правдоподобно предположение, что в раннемеловую эпоху происходило не просто расселение форм, появившихся гораздо раньше, а сама выработка новых форм. Первые представители семейств, доживших до наших дней, появились позже, лишь в позднем мелу. Современные роды имеют еще более скромную историю. Немногие появились в конце позднего мела, а остальные лишь в палеогеновом и неогеновом периодах.
В литературе обсуждались и ландшафтные условия, в которых произошли покрытосеменные. Среди ботаников и палеоботаников довольно популярна гипотеза, что покрытосеменные спустились с гор. Ее впервые независимо друг от друга выдвинули В. А. Вахрамеев и Ч. Арнольд. Гипотеза «горной колыбели» действительно позволяла многое понять. В горах нет условий для захоронения растительных остатков. Накапливающиеся здесь толщи рано или поздно перерабатываются и переоткладываются. Захоронение растительных остатков может быть устойчивым только в низинах. Исключения крайне редки. Естественно, что от растений, населяющих горы, нам достается преимущественно пыльца. Пока раннемеловые покрытосеменные соотносились с современными семействами и родами, часто высокоорганизованными, горная гипотеза позволяла объяснить, почему мы не находим в мезозое по-настоящему примитивных покрытосеменных.
После того как раннемеловые покрытосеменные оказались более своеобразными, чем считалось раньше, и появились основания полагать, что именно в раннем мелу они сделали первые эволюционные шаги, потребность в горной гипотезе стала падать. Против нее были выдвинуты и прямые возражения, в том числе методологические. Она, как заметил В. А. Красилов, «полностью снимает контроль палеоботанических фактов и открывает простор для чисто умозрительных построений. Можно допустить существование цветковых в пермо-триасе, а если такой возраст покажется недостаточно древним, то отодвинуть их возникновение в карбон или даже докембрий… Эта свобода действия, вероятно, в немалой степени способствовала исключительной популярности теории горного происхождения цветковых, которую не смогли поколебать отдельные критические высказывания…».
Такие критические высказывания, как собственные, так и принадлежащие другим специалистам, В. А. Красилов и суммировал. Я остановлюсь лишь на некоторых из них и добавлю свои соображения. Во-первых, те горы, которые представлялись возможной родиной покрытосеменных, возникли геологически слишком поздно. В тех местах, где мы находим древнейшие остатки покрытосеменных, гор по соседству не было. Во-вторых, долго и безвылазно сидеть на горах покрытосеменные едва ли бы смогли. В течение юры и первой половины раннего мела (т. е. до появления первой пыльцы покрытосеменных в геологической летописи) климатические колебания вызывали движение поясов растительности по склонам гор. В периоды похолодания покрытосеменные имели все возможности спуститься в низины и попасть в устойчивые захоронения хотя бы в виде пыльцы. Далее, те современные горные формы, которые якобы сохраняют архаичность и свидетельствуют о горном происхождении первых покрытосеменных, известны в палеогеновых и неогеновых отложениях, накопившихся в низинах. Наконец, если покрытосеменные действительно появились в горах низких широт, то можно ожидать их спуск в низины в более высоких широтах. Тогда, чем ближе к экватору, тем позднее должны появляться в разрезе остатки покрытосеменных. На самом деле мы видим обратную закономерность.
К сожалению, от древнейших покрытосеменных нам пока достаются только пыльца и отпечатки листьев (фиг. XV). Ботаникам же больше всего хочется узнать, какими были первые цветки. Об этом геологическая летопись пока умалчивает. Наиболее древние органы размножения относятся уже к концу раннего мела и представлены не цветками, а плодами (фиг. XVI). Если бы эволюция покрытосеменных начиналась с крупных цветков, как у магнолии (так считают многие ботаники), то мы находили бы вместе с листьями и остатки крупных лепестков. Когда отцветает магнолия, то земля бывает усыпана побуревшими лепестками. Ничего похожего на лепестки крупных цветков палеоботаники не находят в нижнемеловых отложениях. Есть и другие палеоботанические свидетельства того, что первые покрытосеменные имели не крупные, двуполые и одиночные цветки, а мелкие, однополые, собранные в кистевидные соцветия. Не исключено, что и название «цветки» к ним мало подходило, поскольку у них, может быть, не было лепестков, так что они больше напоминали по внешнему виду органы размножения голосеменных.
Какая группа голосеменных растений дала начало покрытосеменным, мы все еще в неведении. Многие свойственные покрытосеменным признаки появляются независимо у разных голосеменных. У некоторых даже образовывалось подобие завязи. В качестве возможных прародителей перебывали чуть ли не все группы высших растений. Высказывалось предположение, что происхождение покрытосеменных сопровождалось гибридизацией между разными группами. Речь не обязательно идет об обычной половой гибридизации. Генетическую информацию от одной группы к другой могли переносить, скажем, вирусы. С точки зрения генетики в этом нет ничего невероятного.
Впрочем, как ни интересно обсудить возможных предков покрытосеменных и пути их эволюции, здесь не место этим заниматься. Иначе нам придется залезать в дебри морфологии и систематики растений. К сказанному я хочу добавить лишь одно. Проблема происхождения покрытосеменных настолько интересует ботаников, что они спешат ее решить, не дожидаясь, пока будут расшифрованы палеоботанические документы. Странным образом, интерес к проблеме почти не стимулирует исследователей заняться этими документами как следует. О происхождении покрытосеменных рассуждают сотни людей; статьи, посвященные умозрительному разбору предлагавшихся гипотез, исчисляются тысячами. Палеоботаников же, которые занимаются конкретными документами по древнейшим покрытосеменным, тщательно ищут и изучают их остатки в меловых отложениях, анализируют распределение в пространстве и времени уже открытых остатков, буквально единицы. Их можно пересчитать по пальцам. Если бы несколько процентов энергии, затрачиваемой на умозрительные дискуссии, направить на палеоботанические исследования, то, можно надеяться, многие аспекты проблемы стали бы быстро проясняться.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава VIII Манипуляции с молекулами
Глава VIII Манипуляции с молекулами В предыдущих главах мы неоднократно говорили о роли ДНК, о функции генов, о синтезе РНК как о первом этапе реализации генетической информации. Почти во всех случаях об этих молекулярных процессах судят, не видя самих молекул, фактически
Глава VIII Половая патология
Глава VIII Половая патология Ссылаемся здесь на известное сочинение Краффт-Эбинга: «Половая психопатия», а также на труды Гавелока Эллиса, Гиршфельда и др., в которых приведены многочисленные наблюдения, которых мы здесь, однако, не будем касаться. Должно сказать, что, не
ГЛАВА VIII.
ГЛАВА VIII. Влияние строения производительного слоя земли, в зависимости от обработки, на рост растений.Растения питаются через листья или корни. Задача обработки сводится к тому, чтобы облегчить корням добывание пищи из почвы. Корни обладают способностью получать пищу от
Глава VIII Парадоксы Галена
Глава VIII Парадоксы Галена Образ Галена предстает перед нами в окружении парадоксов.Он проложил путь и сам же перегородил его стеной. Не будь его, труды греческих врачей, вероятно, безвозвратно канули бы в вечность. А вместе с тем его практическая деятельность более чем на
Глава VIII. Инстинкт
Глава VIII. Инстинкт Инстинкты подобны привычкам, но отличны от них по происхождению. Многие инстинкты столь удивительны, что их развитие, вероятно, покажется читателю трудностью, достаточной для ниспровержения моей теории. В целом я могу заранее сказать, что не занимаюсь
Глава 211. Абиогенная (химическая) эволюция (VIII)
Глава 211. Абиогенная (химическая) эволюция (VIII) Гипотезы о происхождении жизни на Земле исходят в основном из двух предположений. Это либо гипотеза панспермии (что многих не устраивает, поскольку, как они полагают, лишь отодвигает событие в прошлое и не решает задачу), либо
Глава VI ТАЙНА ПАЦИФИКИ
Глава VI ТАЙНА ПАЦИФИКИ Каждый вечер в девять пятнадцать из международного аэропорта Лос-Анджелеса отправляется рейс авиакомпании Эн-Зи-Ай в Новую Зеландию. За тридцать секунд он проносится над узкой полоской суши, отделяющей конец взлетной полосы от океана. На моторах
Глава VIII ПЕРВЫЕ ЕВРОПЕЙЦЫ
Глава VIII ПЕРВЫЕ ЕВРОПЕЙЦЫ Археологические раскопки в Боксгрове (возле Чичестера) в английском графстве Сассекс продолжались десять лет и были результативными, хотя и неэффективными. Сейчас в Боксгрове карьер, а полмиллиона лет назад здесь была прибрежная равнина, узкая
Глава VIII Система пищеварения
Глава VIII Система пищеварения Пища и питание Мне всегда нравилось выражение «Мы — то, что мы едим»; это относится к кошкам и собакам в той же степени, что и к человеку. От неполноценной пищи прежде всего страдает система пищеварения; другие органы — печень, поджелудочная
Глава 211. Абиогенная (химическая) эволюция (VIII)
Глава 211. Абиогенная (химическая) эволюция (VIII) Гипотезы о происхождении жизни на Земле исходят в основном из двух предположений. Это либо гипотеза панспермии (что многих не устраивает, поскольку, как они полагают, она лишь отодвигает событие в прошлое и не решает
ГЛАВА VIII. ТАИНСТВЕННЫЕ ВЕЩЕСТВА
ГЛАВА VIII. ТАИНСТВЕННЫЕ ВЕЩЕСТВА «Пагуба моряков» Среди бурунов. Зноя и тумана Скрипели и звенели паруса, Раскатывался окрик капитана, И таяли матросов голоса. Был долог путь, Вспененный и упорный. Меж низких звезд И оголенных вод… Эд. Багрицкий Многие морские
Глава 12 Тайна жажды
Глава 12 Тайна жажды Кажется вполне естественным, что одно из самых ранних упоминаний о жажде пришло к нам из пустынь Египта. Около сорока столетий назад Синух, приближенный фараона Аменемхета I, чуть не умер от жажды, когда пересекал Суэцкий перешеек. В одном из бесценных
ГЛАВА VIII БОЛЕЗНИ ЗУБОЧЕЛЮСТНОЙ СИСТЕМЫ
ГЛАВА VIII БОЛЕЗНИ ЗУБОЧЕЛЮСТНОЙ СИСТЕМЫ В наше время ветеринарная стоматология стала выделяться как отдельная, самостоятельная дисциплина, изучающая не только различные заболевания и отклонения в ротовой полости у животных, но и методы их диагностики, лечения и
Глава VIII. Есть ли у человечества будущее?
Глава VIII. Есть ли у человечества будущее? Этот вопрос вызывает интерес у многих современников. Он затрагивается в ряде монографий самого последнего времени (Назаретян, 2001; Глэд, 2005; Арутюнов и Стрекова, 2006; Зубов, 2002).Прежде всего, следует определиться, что понимать под