Аксиома третья
Аксиома третья
Опечатки генетических программ. Редкая книга обходится без опечаток. В издательских кругах бытует характерный исторический анекдот. В 1888 году известному издателю А. С. Суворину удалось добиться у царской цензуры разрешения на издание радищевского «Путешествия из Петербурга в Москву» тиражом… в сто экземпляров. Издательская культура была у Суворина на большой высоте, а в этом исключительном случае он даже заключил пари, что издаст книгу без единой опечатки. Книга вышла – и на обложке стояло: «Сочинение А. И. Радищева» (напомню, что великого демократа звали Александром Николаевичем).
Для чего я рассказал эту историю? Мы уже убедились, что в основе жизни лежит матричное копирование, в принципе аналогичное тому же книгопечатанию. Ясно, что в каналах передачи информации от ДНК к признакам организма и от ДНК родителей к ДНК потомков должен существовать какой-то шум – те же опечатки, только на молекулярном уровне. Каналов без шума не бывает, иное дело, что шум может быть пренебрежимо малым.
Рассмотрим сначала шумы в канале ДНК – ДНК, приводящие к изменению генетических программ. В первую очередь речь у нас пойдет об упаковке генетического материала.
ДНК или РНК простейших вирусов может представлять лишь цепочку нуклеотидов, ничем не защищенную от внешних воздействий (например, от действия ферментов нуклеаз, расщепляющих нуклеиновые кислоты). Однако у сложных вирусов она заключена в белковый защитный чехол.
ДНК бактерий также единичная последовательность. Концы ее стыкуются, и образуется кольцо, похожее на тысячекратно перекрученную ленту Мёбиуса, хорошо известную любителям математики. Ясно что при репликации кольцо это должно разрываться, иначе дочерняя последовательность будет соединена с материнской, как звенья в цепи. К бактериальной ДНК могут присоединиться молекулы белков, но в общем-то она «голая».
Иное дело у высших организмов с оформленным клеточным ядром. Прежде всего генетическая программа у них – многотомное издание. Если генетическая программа бактерии закодирована в одной молекуле ДНК, одной двойной спирали, то в ядре высших организмов – эукариот – их может быть несколько: от двух у лошадиной аскариды до нескольких тысяч у некоторых одноклеточных организмов – радиолярий и ряда растений. Такие тома называют хромосомами. Считается, что каждая хромосома содержит одну молекулу ДНК, но крайней мере у животных. Однако есть сильные доводы в пользу того, что у многих высших растений в хромосоме может быть несколько десятков, а то и сотня идентичных копий. ДНК в хромосомах чрезвычайно хитроумно уложена в комплексе со специальными ядерными белками гистонами. Иначе нельзя упаковать в микронные объемы молекулы длиной во много десятков сантиметров.
Наблюдая за хромосомами во время деления клеток, исследователи обнаружили много форм изменения наследственных программ.
Читателям должно быть известно, что при образовании половых клеток хромосомы не делятся, а расходятся в дочерние клетки, так что получаются гаметы с половинным (гаплоидным) набором хромосом. У человека, например, в нормальных клетках 46 хромосом, а в яйцеклетках и спермиях – 23. При слиянии гамет диплоидный набор восстанавливается.
Но так бывает не всегда. Порой механизм, растягивающий хромосомы по дочерним клеткам, не срабатывает. Одна гамета получается совсем без ДНК, а другая с двойным ее набором. Так возникают полиплоидные клетки и организмы; о часто это наблюдается у растений.
Иногда же в одну клетку попадает лишняя хромосома, а в другой обнаруживается нехватка. Такие явления называются анеуплоидией.
При всех этих перестройках генетическая информация, заключенная в хромосомах, не изменяется. Меняется только ее количество. Полиплоидные клетки, например, могут иметь тройной, четверной и т. д. – до тысячи и более раз! – набор генов.
Анеуплоидный геном – это многотомное, но разрозненное издание. В одной клетке не хватает «тома» инструкций (обычно такие случаи у высших организмов летальны) в другой два одинаковых. Организмы с лишней хромосомой (трисомики) также часто гибнут на ранних стадиях развития или же развиваются с серьезными дефектами. Много таких случаев описано относительно человека.
Иногда перестройка может привести к тому, что хромосома распадается на части. Судьба частей различна: они могут потеряться (делеция), снова воссоединиться в составе прежней хромосомы (иногда в перевернутом виде – инверсия) или же присоединиться к другой (транслокация). Все перестройки, как правило, для организма небезразличны.
Все упоминавшиеся изменения хромосом начинаются с разрыва нуклеотидной цепи ДНК – знаменитой двойной спирали. Поэтому мы должны от тех построек, которые видны в оптический микроскоп, перейти на молекулярный уровень.
Насколько прочны фосфодиэфирные связи, скрепляющие полимерную ДНК, и насколько устойчивы пуриновые и пиримидиновые основания в ДНК к внешним воздействиям?
Это удалось установить с достаточной точностью. Чтобы вызвать единичную мутацию – наследственное изменение генетической программы, – требуется подвести каким-то способом к ДНК энергию в 2,5—3 электронвольта (эВ). Электронвольт – единица энергии: такую энергию приобретает электрон, ускоряемый напряжением в 1 вольт.
Много это или мало? Ведь ДНК в клетке находится в окружении молекул, движущихся с весьма высокой скоростью. Оказывается, что средняя энергия теплового движения молекул при тех температурах, когда жизнь возможна, составляет примерно 1/40 эВ. Иными словами, при физиологических температурах ДНК оказывается достаточно стабильной. Но проблема эта сложнее, чем кажется на первый взгляд.
Не следует забывать, что скорости молекул при хаотическом тепловом движении неодинаковы. Убедиться в этом нетрудно. В 1827 году шотландский ботаник Р. Броун, разглядывая в микроскоп каплю воды с пыльцой растений, обнаружил, что взвешенные в жидкости пыльцевые зерна микронного размера не остаются на месте, а хаотически движутся как бы непрерывно подталкиваемые беспорядочными ударами чего-то невидимого.
Достойно удивления, что Броуново движение не привлекало внимания физиков (может быть, потому, что открыл его ботаник?) до начала нашего века, до исследований Альберта Эйнштейна, польского физика Мариана Смолуховского и французского физика Жана Перрена. А ведь из него не только вытекала непреложность существования молекул, но и возможность оценить их скорости и размеры!
Что происходит с частицей при броуновском движении? Со всех сторон она подвергается ударам молекул. Если она имеет достаточно большие размеры, то удары со всех сторон оказываются скомпенсированными – частица остается на месте. Но если размер ее, допустим, 10-5см, то весьма вероятно, что с какой-либо стороны суммарный импульс будет больше, и частица сдвинется в непредсказуемую сторону.
Иначе и быть не может: ведь скорости молекул разные и флуктуации в их распределении неизбежны. Именно от этих флуктуаций зависит голубой цвет ясного неба, так как на них сильнее рассеиваются синие лучи. Будь скорости всех молекул одинаковыми, солнечный свет не рассеивался бы и солнце светило бы в черном небе, как в космическом пространстве.
Эти же флуктуации кладут предел усилению слабых сигналов в электрических цепях. В конце концов мы слышим лишь треск, результат теплового движения электронов в цепях усилителя. Чтобы уйти за этот предел, приходится охлаждать приемник жидким азотом, водородом, а то и гелием.
Отсюда однозначно следует, что в любой достаточно большой популяции молекул неизбежно найдутся такие, которые могут нарушить структуру гена и вызвать мутацию. Ясно, что такие изменения генетических программ должны обладать следующими свойствами:
1. Они случайны в том смысле, что вероятность каждого единичного изменения не равна единице. Более стабильные части гена мутируют с меньшей частотой, более лабильные – с большей, но мы можем говорить лишь о большей или меньшей вероятности мутаций.
2. Они непредсказуемы, поскольку для предсказания какой-либо мутации мы должны знать координаты и импульсы всех молекул данной клетке.
3. Они не направлены в том смысле, что изменяют генетичеекую программу без учета содержания сохраняющейся в ней информации. Поэтому они только случайно могут оказаться адаптивными, приспособительными.
Не одна температура изменяет содержание генетических программ.
Пожалуй, большее значение имеют кванты жесткого излучения начиная с ультрафиолета, быстро движущиеся элементарные частицы, молекулы веществ, способные реагировать с ДНК (химические мутагены).
Начнем с радиации. Растворы чистых нуклеиновых кислот в видимом свете прозрачны. Однако и ДНК и РНК сильно поглощают ультрафиолет. Максимум поглощения на 260 нанометрах, кванты такой энергии уже могут вызывать и вызывают мутации. Еще более эффективны рентгеновы лучи и гамма-излучение, а также быстрые частицы, возникающие при ядерных распадах. Действие жестких излучений на генетические программы изучает целая отрасль генетики радиогенетика. О ее достижениях в популярной литературе много писалось. Остановимся подробнее на другом вопросе. В случае, когда мутацию вызывает квант или элементарная частица, наиболее четко проступает принцип необходимости квантовой механики для описания мутагенеза.
Мы уже договорились с самого начала, что физические законы, физические аксиомы должны оправдываться и для тех случаев, когда мы имеем дело с живой материей.
Значит, и здесь должен действовать принцип квантовомеханической неопределенности, который впервые сформулировал В. Гейзенберг в 1927 году.
Выражается этот великий принцип на вид простой формулой
?p•?x?h
и утверждает, что нельзя определить одновременно координату и импульс (то есть энергию) элементарной частицы точнее, чем следует из этого неравенства.
Произведение погрешности в определении координаты (?x) на погрешность в определении импульса (?p) не может быть меньше постоянной Планка h, равной 6,626 • 10–34Дж•с. Это ничтожная величина, поэтому, живя в макромире, мы ее не замечаем. Будь она побольше, мы бы жили в мире без ясных очертаний предметов и без четких траекторий движения. Но ведь мутации происходят в микромире. Для всего гена неопределенность пренебрежимо мала. Однако чтобы вызвать мутацию, нужно довести определенное количество энергии до участка гена размером всего лишь 10-7см (так называемый радиус эффективного обмена, впервые измеренный Н. В. Тимофеевым-Ресовским и К. Циммером). Здесь уже принципом неопределенности пренебрегать нельзя. О том, что для описания мутагенеза необходимо использовать принцип Гейзенберга, писали еще Н. В. Тимофеев-Ресовский и Паскуаль Йордан в 40-х годах. В последнее время то же утверждает и Манфред Эйген. Короче, мы можем лишь с некоторой долей вероятия предполагать, обладает ли частица нужной для мутации энергией и попадет ли она в зону эффективного объема.
Не только принцип Гейзенберга – любое другое положение квантовой механики делает процесс мутагенеза вероятностным. Вот еще пара примеров. Кроме стабильных атомов углерода и фосфора в состав ДНК могут входить их радиоактивные изотопы. Вероятность этого возрастает с увеличением радиоактивного фона нашей планеты. Местоположение атомов C14 и P32 в нитях ДНК, разумеется, случайно. А радиоактивный распад – тоже случайный процесс: знать, какой из атомов в данный момент распадется и вызовет мутацию, нам не дано, и запрет этот принципиальный.
Наконец, есть мутации, не индуцированные каким-нибудь внешним по отношению к ДНК агентом, а так называемые спонтанные. Водородные связи, скрепляющие двойную спираль ДНК, обусловлены ионами водорода (протонами). Если при раскручивании спирали ДНК, которое обязательно происходит при ее репликации, оба протона перейдут к одному из оснований, возможны другие комбинации, кроме АТ и ГЦ. А неверное спаривание оснований – мутация. Переход протона от одного основания к другому называется туннельным, описывается уравнениями квантовой механики, и все, что мы можем сказать о нем, то, что он осуществляется с частотой 1011…1012в секунду. Время и место каждой единичной спонтанной мутации непредсказуемо принципиально.
А что же вещества, изменяющие структуру гена, – мутагены? Впервые в 1932—1934 годах советские ученые В. В. Сахаров и М. Е. Лобашов независимо друг от друга получили мутации у дрозофилы, действуя на мушек йодом и рядом других веществ. С того времени список мутагенов возрос тысячекратно, теперь каждое химическое соединение, которое предполагается внедрять в практику (лекарства, красители, пластмассы и т. д.), испытывается на мутагенный эффект. Открыты – И. А. Рапопортом и Ш. Ауэрбах – так называемые супермутагены, вызывающие мутации у 100 процентов обработанных ими особей.
Может показаться, что химические мутагены не имеют особого значения в природе, во всяком случае до появления химической промышленности. Простой пример свидетельствует, что это не так. Даже такое, с позволения сказать, вещество, как ион водорода, может индуцировать мутации.
Активная реакция среды измеряется, как вы знаете, в единицах pH. Это взятый с обратным знаком логарифм концентрации водородных ионов. Например, когда реакция нейтральная, pH равно семи и концентрация ионов водорода 10-7моля. ДНК довольно устойчива к подкислению или подщелачиванию, однако при pH ниже четырех начинается отщепление пуриновых оснований – гуанина и аденина.
Содержимое клетки в общем-то сильная буферная система, поддерживающая pH в районе семи (это не относится к специализированным органам вроде желудка, где высока концентрация соляной кислоты, или же слюнных желез некоторых улиток, которые могут выделять довольно концентрированную серную кислоту). Но вспомним о размерах клеток и обязательных флуктуациях молекул. Объем бактериальной клетки всего 2•10-12см3, значит, при pH = 7 в ней всего примерно 120 водородных ионов. А это означает, что очень вероятна флуктуация: ионы водорода соберутся около, например, гуанина и отщепят его от пентознофосфатного остова ДНК. Произойдет мутация, и опять же случайным, непредсказуемым образом.
На этом можно было бы и закончить обзор изменений генетических программ. Но хотелось бы рассказать о недавно открытых хромосомных перестройках, удивительных и ранее казавшихся невозможными.
Сначала в геномах бактерий, а потом и в геномах высших организмов обнаружили удивительные «кусочки» ДНК, которые могут перемещаться с места на место в хромосоме. Их образно назвали прыгающими, генами. Различают две категории таких последовательностей. Есть короткие (500—1500 пар нуклеотидов) и длинные (более двух тысяч). Они могут вычленяться из единственной хромосомы бактерии (этот процесс называется эксцизией, выстриганием) и встраиваться в нее в другом месте (инсерция). И не только в хромосому, прыгающие гены могут включаться в плазмиды – последовательности ДНК, существующие в бактериальной клетке независимо. Так же легко прыгающие гены могут встраиваться в геномы бактериофагов. Удивительного в этом ничего нет: существует вполне убедительное мнение о том, что плазмиды – «домашние», прирученные бактерией фаги.
Короткие инсерционные сегменты обладают многими удивительными свойствами. Почему-то оба конца их состоят из одинаковых последовательностей. Иногда же концевые участки перевервуты: на одном конце, например, АТГАГ, на другом – ГАГТА. Встраиваясь в ген, они часто блокируют процесс транкрипции – ген теряет активность. И наоборот, встраиваясь в перевернутом виде, они могут тот же ген активировать. Иными словами, они включают и выключают гены.
Еще более удивительны длинные прыгающие последовательности транспозоны. Именно они, встраиваясь в фаги, переносят от бактерии к бактерии гены устойчивости к антибиотикам. Так что фаги не только могут убивать бактерии или быть их «приживальщиками». Они переносят в мире микроорганизмов генетическую информацию. Так, насекомые не только вредят растениям, но и опыляют их. Ведь первые цветковые растения опылялись ветром. На их пыльниках кормилось много насекомых. Перепачканные пыльцой жуки и примитивные бабочки перелетали с пыльника на пыльник, и в результате возникли такие пары «растение – насекомое», которые не могут существовать друг без друга. Юкка, например, не может опыляться иначе как с помощью юкковой моли. И между фагом, лизирующим, «пожирающим» бактерию, фагом-«приживальщиком» и приносящей пользу плазмидой, которая переносит транспозоны, есть все переходные ступени.
Прыгающие гены описаны и у высших, ядерных организмов. У дрожжевых грибков они, например, в зависимости от положения в хромосоме, определяют пол клетки. Обнаружили их также у кукурузы и дрозофилы. Вот на последнем объекте хотелось бы остановиться подробнее.
Генетиков давно занимали случаи, когда гены в популяциях плодовой мушки-дрозофилы вдруг теряли стабильность и частота мутаций возрастала во много раз. Оказалось, что нестабильность того или иного гена обусловливается встроившимся в него инсерционным сегментом. В некоторых случаях удалось проследить, как прыгающий ген в течение нескольких лет кочует от одного участка хромосомы к другому и от одной хромосомы к другой, каждый раз вызывая новую мутацию. Шведские ученые Г. Исинг и К. Рамель проследили до сорока прыжков одной такой инсерции. Результатом была или полная инактивация гена, приводящая к смерти (летальная мутация), или же мутации «розовые глаза», «грубые глаза» (с неправильным расположением фасеток) и целый ряд других.
Такие феномены наблюдались не только в экспериментальных, рассаженных по пробиркам популяциях дрозофилы, но и в природе. Известны так называемые «модные» мутации. Название несколько неожиданное, но удачное. Например, в 30-х годах частота мутаций, вызывающих желтую окраску тела и белую окраску глаз, возросла в сотни раз, и эта «мода» длилась до 40-х годов. В 1968 году у дрозофил возникла «мода» на мутацию abnormal abdomen (брюшной отдел уродливой формы), которая продолжалась пять лет и была вытеснена лишь мутацией «опаленные щетинки».
По-видимому, причина «модных» мутаций – вирусы. Известно, что вирусы могут встраиваться в хромосомы не только бактерий, но и ядерных организмов (это доказано для аденовирусов, вируса герпеса и опухолеродных перерождающих нормальную клетку в злокачественную). Инсерция вируса в геном вызывает мутацию того гена, в который он встроился.
Примечательно, что есть явное сходство между интронами, о которых говорилось в прошлой главе, и инсерционными сегментами. Те и другие могут разделять смысловую часть гена, не находят отражения в аминокислотной последовательности кодируемого геном белка и, по-видимому, могут включать и выключать гены.
А что значит: включать и выключать?
По-видимому, это выглядит так: регуляторные элементы клетки могут узнавать и не узнавать встроенный в ген интрон (инсерционный сегмент). В первом случае идет транскрипция предшественника мРНК, во втором – нет. В конце концов, это находит отражение в изменениях признаков фенотипа. И если интрон перепрыгнет в другой ген, то он будет активировать его, а не тот, из которого он ушел.
Третья аксиома. Теперь пора подытожить все, что мы знаем о наследственных изменениях генетических программ и сформулировать аксиому биологии № 3.
Прежде всего, эти изменения случайны и ненаправленны. Их можно сравнить с шум в канале информации от родителей к потомкам. Если мутация полностью исказит смысл передаваемой по цепи поколений инструкции, она будет смертельной (летальной). Это бывает, когда в результате мутации блокируется синтез жизненно важного фермента.
Наоборот, часты случаи, когда мутация не сказывается на признаках фенотипа. Вспомним, что генетический код вырожден и одна и та же аминокислота кодируется несколькими кодонами. Если в результате мутации один кодон заменится другим, но синонимичным, в полипептидную цепь включится та же аминокислота и изменения фенотипа мы не обнаружим.
Между этими двумя полюсами лежит огромная область мутаций, так или иначе изменяющих фенотип. В каких-то конкретных условиях они могут оказаться полезными, носители их с большей вероятностью оставят потомство и передадут их потомству.
Это и есть дарвиновская неопределенная изменчивость – исходный материал для эволюции.
Аксиому биологии №3 мы можем сформулировать так:
В процессе передачи из поколения в поколение генетические программы в результате многих причин изменяются случайно и ненаправленно, и лишь случайно эти изменения оказываются приспособительными.
Третья аксиома вытекает из важнейших физических постулатов – из практической невозможности знать координаты и импульсы всех молекул в клетке, из чего следует распределение энергий молекул по Максвеллу, и из принципиальной невозможности достаточно точно определить координаты и импульсы частиц, атакующих гены (принцип Гейзенберга).
А так как и перераспределение энергий в совокупности молекул, и взаимодействие молекул в химических реакциях в конце концов можно свести к квантовомеханическим процессам, то и получается, что аксиома №3 – следствие квантовой механики.
Далеко не всем это нравится. М. Эйген в книге, на которую я уже ссылался, по этому поводу вспоминает Эйнштейна. В письме от 4 декабря 1926 года Эйнштейн писал Максу Борну: «Квантовая механика внушает большое почтение. Но внутренний голос говорит мне, что это не истинный Иаков. Теория дает много, но едва ли она подводит нас ближе к тайне Старика. Во всяком случае, я убежден, что он не играет в кости…»
Старик – это бог, Не следует считать Эйнштейна религиозным. Поклонник Спинозы, он называл богом реально существующую природу.
С тех пор прошло более 50 лет: Квантовая механика многократно показала свою непротиворечивость природе, и вряд ли даже Эйнштейн сейчас выступал бы против нее. Однако в биологии до сих пор есть авторы, не признающие ненаправленной случайной изменчивости по Дарвину.
В конечном счете они склоняются к точке зрения, высказанной за полвека до Дарвина в 1809 году великим французским натуралистом Ламарком. Согласно Ламарку, организмы в процессе эволюции изменяются, но изменения эти массовые, направленные и приспособительные. Со времени Ламарка их называли по-разному: наследованием благоприобретенных свойств, ассимиляцией условий внешней среды и т. д. и т. д.
В нашей литературе наиболее основательно эту точку зрения отстаивал крупный ихтиолог и географ Л. С. Берг. Он, правда, называл свои воззрения не ламаркизмом, а номогенезом (эволюцией на основе закономерностей, то есть закономерных, а не случайных изменений). Но хотя в своей книге, вышедшей в 1922 году, Берг решительно отрекся от ламаркизма, никого это не обмануло. Уже тогда оппоненты Берга поняли, что ламаркизм и номогенез различаются лишь терминологией.
Ибо, по Бергу, наследственные изменения захватывают массы особей, идут направленно и должны быть изначально целесообразными, приспособительными. Берг, как и Ламарк, вообще считал приспособительные реакции изначальным свойством живого.
Современные номогенетики как-то стыдливо обходят последний тезис Берга, предпочитая говорить лишь о направленности наследственных изменений, а не о целесообразности их. Но направленные мутации должны быть приспособительными – ведь все организмы на Земле к каким-то условиям приспособлены.
Мне доводилось слышать выступление физика, в котором выдвигалась такая идея. Мутации сами по себе вредны или нейтральны и обеспечить эволюцию материалом не могут. Движущая сила эволюции – приспособительные изменения, которые происходят крайне редко (например, раз в тысячу лет). Из-за их редкости мы их обнаружить не можем, поэтому все опыты, опровергающие приспособительное изменение генетических программ, оказываются бездоказательными.
Но физик должен был бы понять, что номогенез, или ламаркизм, – называйте, как хотите, – основан на нарушении принципа причинности. Следствие (приспособленность) в нем определяет причину (изменение генетической программы). И опять мы возвращаемся к Аристотелю с его конечной причиной и целью одновременно. Такие взгляды на эволюцию, называются телеологическими.
Для меня представляется совершенно непостижимым, что в наши дни находятся вполне серьезные ученые, не понимающие того, что было предельно ясно еще в прошлом веке Герцену. Вспомните его «Былое и думы»:
«Естествоиспытатели, хвастающие своим материализмом, толкуют о каких-то вперед задуманных планах природы, о ее целях и ловком избрании средств… Это – фатализм в третьей степени, в кубе; на первой кипит кровь Януария, на второй орошаются поля дождем по молитве, на третьей открываются тайные замыслы химического процесса, хвалятся экономические способности жизненной силы, заготовляющей желтки для зародышей и т. п.».
Сказано как будто про наших современных номогенетиков. Я уже писал об этом подробнее в другой своей книге[7]. Здесь, пожалуй, будет довольно нескольких примеров того, как направленная приспособительная изменчивость вступает в противоречие с принципом причинности.
Допустим существование неких актов приспособления, чрезвычайно редких и потому ненаблюдаемых. Нет проще приема сделать их наблюдаемыми. Надо лишь увеличить число наблюдений. Пример из той же физики: нейтрино взаимодействует с веществом с чрезвычайно низкой вероятностью, эта элементарная частица может пронизать насквозь земной шар, и вероятность того, что она вступит в реакцию с каким-либо ядром, совершенно ничтожна. Но физики уверенно детектируют нейтрино и антинейтрино. Детектор здесь – большое количество атомов хлора (обычно в составе той жидкости, которая применяется в химчистке – тетрахлорэтилена). А источник искомой частицы – ядерный реактор, испускающий мощные потоки нейтрино.
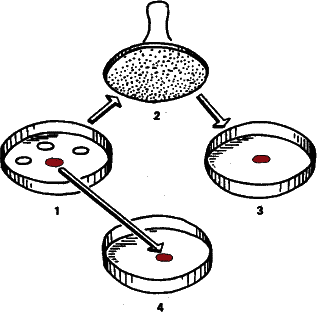
Рис. 36. Супруги Ледерберги выращивали в чашках Петри на поверхности агара тысячи колоний бактерий, а затем стерильным бархатным штампом переносили их с сохранением прежнего положения в другие чашки, где среда содержала смертельную дозу антибиотика. Если число пересеваемых колоний было достаточно велико, всегда находилась хотя бы одна, устойчивая к антибиотику. При этом нетрудно было убедиться, что материнская колония также состоит из устойчивых бактерий, хотя с антибиотиками они и не встречались.
Американские исследователи Джошуа и Эстер Ледерберги исследовали процесс приспособления бактерий к антибиотикам. Этот феномен – проклятие современной медицины. Прославленные пенициллин, стрептомицин, олеандомицин быстро теряют свою эффективность Нужно сказать, что помогают микробам в данном случае и врачи со слабой биологической подготовкой. Они прописывают больным недостаточные дозы (из боязни «как бы чего не вышло»), то есть фактически ведут в популяциях бактерий отбор на устойчивость к антибиотику. Или же, наоборот, впадают в другую крайность и закармливают пациента антибиотиками при пустяковой простуде, а в результате тот же антибиотик окажется недейственным при двустороннем воспалении легких. Вспомните о переносах генов плазмидами и бактериофагами! Ген, определяющий, например, устойчивость бактерии к стрептомицину, легко может быть передан даже бактериям другого вида. В последнее время возникла серьезная проблема с гноеродными золотистыми стафилококками – современный врач с набором антибиотиков оказался в положении хирурга прошлого века, бессильного перед послеоперационными осложнениями. Выход в одном – все время обновлять арсенал антибиотиков, по возможности обгоняя бактерий.
Но вернемся к Ледербергам. Они взяли несколько сот чашек Петри с питательным агаром и посеяли на них культуру бактерий, чувствительных к антибиотику. На тех местах, где на поверхность питательного студня попадали бактерии, эти бактерии начинали расти, делиться, и через некоторое время агар покрывался колониями, скоплениями бактерий, выросших из одной клетки-родоначальницы. Исследовали сразу сотни и тысячи колоний, что увеличивало шансы на обнаружение редко наблюдаемого эффекта.
Затем Ледерберги запаслись несколькими сотнями штампов – кружков, равных по диаметру чаше Петри и обтянутых бархатом. Штампы стерилизовались в автоклаве, чтобы не занести в культуры чего-нибудь лишнего. Затем стерильный штамп прикладывался к поверхности агара; в каждой колонии часть бактерий прилипала к ворсинкам бархата. А затем штамп прикладывался к поверхности агара в другой чашке.
Но этот агар уже содержал антибиотик, причем в концентрации, смертельной для данных бактерий.
И «отштампованные» на его поверхности колонии бактерий перестали развиваться и погибли. Но была подмечена любопытная закономерность: если колоний достаточно много, среди них обязательно найдется хотя бы одна, которая приживется на ядовитом агаре и будет разрастаться, как ни в чем не бывало.
Казалось бы, ясно: произошло направленное целесообразное изменение по Ламарку или же Бергу. Правда, неясно, почему приспособилась лишь одна колония из многих, но это уже деталь.
Но колонии переносились на агар с антибиотиком штампами с сохранением прежнего положения. Поэтому мы всегда можем найти материнскую колонию и убедиться, что она также состоит из бактерий, устойчивых к антибиотику! Приспособление возникло до того, как на популяцию подействовал фактор– антибиотик Налицо явное нарушение принципа причинности. Следствие предшествует причине. Но если мы примем, что подобные изменения генетических программ возникают в результате чисто случайных процессов и в некоторых случаях оказываются полезными, никакого нарушения не будет. Но это уже не ламаркизм или номогенез, а отбор случайных мутаций по Дарвину.

Рис. 37. Когда в Англии для борьбы с крысами стали применять антикоагулянт варфарин, очень скоро появились устойчивые к нему популяции «суперкрыс». Эта карта опубликована в 1970 году. Через два года пришлось бы залить красным цветом всю территорию Великобритании. Таково могущество отбора, пусть бессознательного.
Второй пример касается уже высших организмов. Когда в практику борьбы с крысами вошли антикоагулянты, дератизаторам, то есть крысоловам, показалось, что наступает золотой век. У крыс, поевших приманки с варфарином, кровь переставала свертываться, и животные погибали от милейшей царапины, например из-за песчинки, задевшей стенку кишечника. И вдруг везде, где применяли коагулянты, появились популяции крыс, которые на этой отраве благоденствовали. Их назвали суперкрысами.
Приспособление по Ламарку или отбор мутации по Дарвину? Генетики быстро установили, что гены, дающие носителю устойчивость к антикоагулянтам, имелись в популяциях крыс задолго до их внедрения, они встречаются и там, где варфарина и прочих подобных препаратов никогда не применяли. Но встречаемость их крайне низка: у таких крыс повышена способность крови свертываться, и они часто гибнут от тромбов, закупорки кровеносных сосудов. Применение антикоагулянтов произвело среди крыс полную переоценку ценностей: прародителями суперкрыс стали немногочисленные особи – носители этих прежде невыгодных генов.
Из этого примера, кстати, становится ясным, что широко распространенное мнение о вредности мутаций, ненаправленных изменений генетических программ, попросту неверно. Нет мутаций во всех условиях вредных, как нет и безусловно полезных. Все попытки расклассифицировать мутации без учета данных конкретных условий обречены на провал.
Вот еще примеры с нарушением принципа причинности. На севере пресмыкающиеся, в частности змеи, редки. У нас Полярный круг переходит только обыкновенная гадюка благодаря специальному приспособлению. У северных гадюк развитие яиц начинается еще в яйцеводах, так что они рождают уже вполне сформированных змеенышей. А вот африканские примитивные гадюки откладывают яйца. Самая крупная наша гадюка – гюрза – также живородящая, но в южных районах Средней Азии откладывает яйца.
Налицо явное приспособление, повышающее скорость развития яиц в условиях прохладного климата с резкими суточными изменениями температуры. Беременная гадюка большую часть дня проводит где-нибудь в защищенном от ветра месте на припеке, отчего температура ее тела поднимается до 28°, а на ночь скрывается в какой-нибудь расщелине или под корнями деревьев. Фактически она превращается в инкубатор. Несомненно, для будущих гадючат этот признак полезен. А вот полезен ли он самой гадюке? Позволительно усомниться: ведь она лежит на открытом месте, так что ее может схватить любой канюк или аист, да и обмен у «разогретой» змеи идет куда быстрее, так что требуется больше пищи. Энергетически это невыгодно.
Как и все признаки, повышающие вероятность выживания у потомства, но бесполезные и даже вредные для родителей, это свойство гадюки необъяснимо с точки зрения прямого приспособления. Разве что неродившиеся змееныши передают по какому-то каналу информации своей будущей маме: «Мама, чтобы мы выжили, не спеши откладывать яйца, в которых мы развиваемся». По-видимому, дело обстоит проще: шел отбор на закрепление в популяции этого признака, возникшего случайно.
Еще более наглядны примеры из жизни растений. Как вы знаете из школьного курса, для высших растений характерно чередование поколений. Размножающийся спорами спорофит порождает растение, продуцирующее половые клетки – гаметы (гаметофит). Если у папоротников и плаунов гаметофит самостоятельное, хотя и редуцированное растение, у цветковых он паразитирует на спорофите. То, что мы видим у дуба и фиалки, сосны и кактуса, это все спорофит. А гаметофит – это части цветка: завязь и пыльцевые зерна. Мужской гаметофит – пыльцевое зерно – состоит всего-то из трех клеток!
А теперь посмотрите на цветок лютика. Лепестки его, принадлежащие спорофитному поколению, образуют идеальной формы рефлектор, концентрирующий солнечные лучи на пыльниках и завязи, чтобы быстрее развивался гаметофит. Опять приспособительный признак, пригодный для будущего, некое «воспоминание о будущем» и необъяснимый с точки зрения прямого приспособления. Разве что мы наделим цветок способностью предвидеть и солидными знаниями оптики и физической химии.
Подытожим все сказанное. Случайные, ненаправленные и неприспособительные изменения генетических программ (мутации) – непреложный факт, доказанный тысячами разных способов. Теперь мы знаем их молекулярные механизмы и уверенно повышаем частоту их возникновения. Не можем лишь одного – вызывать направленную нужную нам мутацию. Но, как мы должны были бы убедиться, это невозможно до тех пор, пока мы не знаем структур и функций всех генов в клетке и не научимся изменять один ген, не изменяя остальных. В природе без вмешательства человека это невозможно принципиально.
Почему же до сих пор есть люди, яростно борющиеся с неопровержимыми фактами?
Вероятно, по той простой причине, что не могут принять идею случайных изменений («Старик не играет в кости»!).
И по той же причине номогенез (в смягченном, ретушированном, но сохранившем всю свою сущность виде) поддерживается некоторыми исследователями до наших дней. «Наука – враг случайностей», все должно быть закономерно. А то, что такие подходы, мягко говоря, философски беспомощны, номогенетиков не волнует. Лишь бы не принять в качестве источника материала для эволюции случайный процесс. От теории требуется, чтобы она объясняла, почему на земле должны быть кошки и мыши и почему мыши должны прятаться от кошек в норы.
О подобных взглядах иронично писал Энгельс («Диалектика природы», очерк «Случайность и необходимость»): «Если тот факт, что определенный стручок заключает в себе шесть горошин, а не пять или семь, представляет собою явление того же порядка, как закон движения солнечной системы или закон сохранения энергии, то на деле не случайность поднимается до уровня необходимости, а необходимость снижается до уровня случайности». По большому счету эволюции наличие на Земле кошек и мышей столь же случайное событие, как наличие в стручке шести или семи горошин, и бессмысленно выводить его из свойств протопланетной туманности.
Мутации: много их или мало? Не так давно казался существенным вопрос: много или мало мутаций изменяет генетические программы? Физик Эльзассер полагал, что их чересчур много, настолько, что ДНК просто не может быть хранителем наследственной информации. Эту роль он отводил неким нематериальным «биотоническим законам».
Конечно, доказать мутацию в результате действия нематериального фактора трудно, но еще труднее доказать самое его существование. Поэтому перевес был на стороне другой гипотезы: мутаций слишком мало чтобы они могли обеспечить должную скорость процесса эволюции.
Действительно, мутация каждого отдельно взятого гена происходит с довольно низкой частотой (10-4…10-8). Многих к тому же удивлял один непонятный факт: у микроорганизмов мутации отмечались реже, чем у высших организмов. Но дело в том, что у бактерий и геном меньше. Легче набрать без опечаток газету, чем книгу достаточно большого объема. Когда пересчитали частоту мутаций на единицу содержания ДНК в геноме, скорости мутагенеза у всех земных организмов практически совпали.
Представление о том, что мутаций недостаточно для объяснения темпов эволюции, кажется очень привлекательным для врагов «всяческих случайностей». Оно проникло даже в научно-фантастическую литературу: в одном (не из лучших) рассказе люди далекого будущего усердно помогают эволюции на других планетах – ведь природных мутаций недостаточно! Автор не ведал, что представление, которое он принял за научную истину, самое фантастическое в его рассказе.
Рассмотрим этот вопрос подробнее. Да, мутации наблюдаются в природе довольно редко. Чтобы повысить их частоту, селекционеры применяют мутагены, лучи Рентгена и кобальтовые пушки. И все-таки вспомним длинный список факторов, повреждающих ДНК, приведенный в начале этой главы. Если принять, что изменения идут только в результате отщепления пуриновых оснований, при случайных изменениях температуры и pH, то простой расчет показывает: даже при этом клетки каждого человека теряли бы 50 тысяч нуклеотидов в сутки! За 70 лет жизни человек потерял бы 40 процентов всех пуриновых оснований. Ясно, что мы пришли к абсурду. При таком темпе он просто не дожил бы до 70 лет, а погиб еще в утробе матери. На деле люди порой вдвое перекрывают этот возраст и умирают отнюдь не из-за недостатка пуринов.
Следует прийти к выводу, что генетические программы заложенные в каждом из нас, чрезвычайно помехоустойчивы. Стабильность их определяется отнюдь не химической инертностью, а целым рядом особенностей структуры и специальными системами, которые отфильтровывают помехи или корректируют опечатки – как вам угодно – на каждом новом поколении клеток.
Первый из механизмов борьбы с помехами, самый простой, но не самый эффективный, – повторение, дупликация. Вспомним, что у каждого из нас в клетках по два генных набора, от отца и от матери.
Если в одном из них произошла мутация по важному гену, мы можем этого и не заметить: ген второго набора компенсирует мутацию и она в фенотипе не проявится, но только до тех пор, пока оба одинаково поврежденных гена не встретятся в одной оплодотворенной яйцеклетке. Вероятность этого повышается при близкородственном скрещивании. Недаром законодательства всех стран запрещают браки между близкими родственниками.
У полиплоидов геном напоминает тот разговор двух судовых радистов, о котором я уже вспоминал. Однако этот механизм очень уж прямолинеен и ведет к большим затратам и осложнениям. Практичнее дублировать не весь геном, а лишь наиболее важные его части. Так оно и есть, важнейшие последовательности ДНК представлены у высших организмов многими десятками, а то и сотнями копий, например те гены, в которых кодируются ядерные белки – гистоны. Примечательно, что многократно дублированы в основном не структурные гены, кодирующие белки, а регуляторные. Структурные гены в основной своей массе уникальны. Это имеет глубокий смысл: изменение каждого белка имеет шанс проявиться в фенотипе, чтобы пройти оценку на полезность в горниле эволюционных процессов. А вот регуляторные системы обязательно должны сработать, предоставить этот шанс структурному гену. Поэтому они должны быть многократно дублированы.
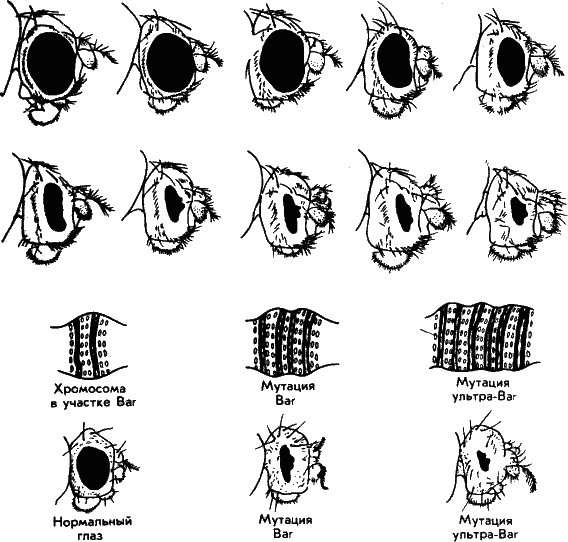
Рис. 34. Наверху – некоторые мутации глаз плодовой мушки дрозофилы. Генетикам очень повезло, что у нее, как и других двукрылых в клетках слюнных желез имеются гигантские хромосомы. Они поперечно исчерчены; каждая полоска в первом приближении соответствует одному гену. Удалось показать, что мутация Ваг, более чем вдвое снижающая число фасеток в глазу, обусловлена удвоением, дупликацией одного гена. Если же ген утраивается (мутация ультра-Ваг) фасетки практически редуцируются.
Этого мало. На заре эволюции, в эпоху становления генетического кода, триплеты ДНК кодировали аминокислоты, по всей вероятности, не столь жестко как сейчас. В результате возник весьма помехоустойчивый механизм кодирования аминокислот тройками нуклеотидов, особенности которого описаны советским биофизиком М. В. Волькенштейном. Можно сравнить принципы построения генетического кода с пишущей машинкой Остапа Бендера. Как вы, конечно, помните, машинка конторы по заготовке рогов и копыт имела кавказский акцент: у нее не было буквы «е», и ее пришлось заменять буквой «э» (…приложэниэ. Бэз приложэний). Почему же великий комбинатор заменил «е» буквой «э», а не какой– либо другой? Это всем понятно: подбиралась буква, наиболее близкая по звучанию. Другая бы искажала смысл слова.
Нечто подобное происходит при мутациях структурных генов. Генетический код устроен таким образом, что во многих случаях смысл триплета не изменяется (кодируется та же аминокислота) или изменяется незначительно. А что значит – незначительно? Снова грамматическая аналогия. Буквы бывают гласные и согласные. Аминокислоты и остатки их, слагающие белки, делятся на полярные и неполярные.
Отличаются они по взаимодействию с молекулами воды – полярные аминокислоты, такие, как лизии, гистидин, аргинин и другие, притягивают молекулы H2O, «смачиваются» водою. Неполярные аминокислоты, такие, как глицин или же аланин, более гидрофобны, они отталкивают молекулы воды и охотнее взаимодействуют друг с другом.
Чередованием полярных и неполярных аминокислот в белке определяется его третичная структура. Неполярные остатки слипаются друг с другом полярные взаимодействуют с водой и в результате образуется сложная форма, порой напоминающая творение скульптора-абстракциониста. С той лишь разницей, что она содержательна: строго соответствует выполняемой этим белком функции.
А что будет, если в результате мутации полярный остаток в белке сменится на неполярный (или наоборот)? Форма молекулы может измениться настолько, что белок не сможет выполнять свою функцию. Гемоглобин не будет связывать кислород, фермент не сможет ускорять химическую реакцию, и организм, носитель данной мутации, скорее всего, погибнет.
Тем, что это происходит относительно редко, мы обязаны хитроумному устройству генетического кода. Еще тогда, когда между аминокислотами в белке и тройками нуклеотидов в нуклеиновой кислоте не было жесткого соответствия (орфография еще не установилась!), эволюция отобрала наиболее помехоустойчивый вариант. Прочие конкуренции с ним не выдержали и сошли с жизненной арены.
Как справляется с помехами генетический код? Возьмем для примера какую-нибудь аминокислоту. Аланин в матричной РНК может кодироваться четырьмя символами:
ГЦУ, ГЦЦ, ГЦА, ГЦГ. Нетрудно сообразить, что любая замена третьей «буквы» в кодоне не изменит его смысла, в белок включится тот же аланин. Но есть и другие аминокислоты, кодируемые меньшим числом триплетов (например, аспарагиновая. и глутаминовая). Оказалось, что за исключением тех случаев, когда в результате мутации возникает бессмысленный кодон, полярность аминокислотного остатка не меняется. Значит, не изменится существенно и форма белковой молекулы, и ее пригодность для выполнения функции. Несмотря на ошибку, «прочесть» инструкцию можно. Чем не машинка Остапа Бендера?
В общем, из 526 возможных замещений (опять же не считая бессмысленных, обрывающих синтез белка), 364, более чем две трети, не меняют полярность аминокислотного остатка. Хуже, если мутация захватывает не один, а два, а то и три нуклеотида, но вероятность таких мутаций много меньше.
Но самое интересное я, как обычно, оставил под конец. Ведь и резервные гены, и особенности кодирования, снижающие эффект мутации, в конечном счете – пассивные средства борьбы с помехами. У генетических программ всех организмов, начиная с бактерий и кончая человеком, есть весьма действенные механизмы активной защиты. Такие процессы называются репарационными.
Репарация ДНК активно защищает генетическую программу клетки от повреждений. Рассмотрим один из ее механизмов на конкретном примере.
Ультрафиолетовые лучи – мощный мутагенный фактор. Наиболее часто они вызывают характерные мутации – сшивки двух соседних пиримидиновых оснований (Ц и Т). Такие сшитые основания не могут быть транскрибированы в мРНК, и процесс дупликации ДНК на них также кончается.

Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Аксиома первая
Аксиома первая Итак, в предыдущей главе мы пришли к выводу, что один из главных признаков живого – организация, структура. Аминокислоты в белках, внутриклеточные элементы, клетки в тканях и ткани в органах образуют упорядоченные структуры. Но все эти структуры, как уже
Аксиома вторая
Аксиома вторая И снова немного истории. В 1927 роду на III Всесоюзном съезде зоологов, анатомов и гистологов в Ленинграде наш блестящий биолог Николай Константинович Кольцов сделал доклад, в котором впервые была четко сформулирована вторая аксиома биологии. Принцип
Аксиома четвертая и последняя
Аксиома четвертая и последняя Из второго начала термодинамики, о котором говорилось в начале этих очерков, вытекает чрезвычайно важное следствие: нельзя получить энергию только отнимая у какого-либо тела тепло. При этом нужно нагревать какое-то другое тело.Помню, как
Часть третья. ИСЧЕЗАЮЩИЕ ДЖУНГЛИ
Часть третья. ИСЧЕЗАЮЩИЕ ДЖУНГЛИ Конечно, Бобру было б лучше всего Добыть подержанную кольчугу… «Охота Ворчуна» ПРИБЫТИЕ Я сидел под деревом, усыпанным огромными алыми цветами, и задумчиво потягивал пиво, когда послышался рокот моторной лодки. С высокой кручи
Третья часть
Третья часть В этой части мы рассмотрим вопросы, непосредственно не относящиеся к теме инстинктов (в узком смысле), но весьма тесно с нею связанные. Эти вопросы часто поднимаются в дебатах с противниками врождённости человеческого поведения, и от ясности ответов на них
Глава третья Выращивание потомства
Глава третья Выращивание потомства У голой обезьяны родительское бремя тяжелее, чем у любого другого животного. Если кто и может состязаться с нами в этом отношении, то в категориях интенсивности, но только не экстенсивности. Прежде чем оценить значение этой тенденции,
Глава третья О преломлении
Глава третья О преломлении Подобно тому, как явления отражения были объяснены волнами света, отраженного от поверхности гладких тел, подобно этому прозрачность и явления преломления мы объясним при помощи волн, распространяющихся внутри и через прозрачные тела, будь то
Книга третья
Книга третья Вопрос 29. Не являются ли лучи света очень малыми телами, испускаемыми светящимися веществами? Ибо такие тела будут проходить через однородные среды без загибания в тень, соответственно природе лучей света. Они могут иметь также различные свойства и способны
Глава третья
Глава третья Поощряемый Спенсером Кертисом Брауном, Даррел сел писать вторую книгу, посвященную путешествию в Британскую Гвиану. Спенсер весьма разумно посоветовал Джерри подготовить ее, не дожидаясь выхода «Перегруженного ковчега», дескать, если первая книга станет