Электрические рецепторы рыб
Электрические рецепторы рыб
По степени развития электрической чувствительности рыб можно подразделить на две группы. К первой относятся почти все виды, имеющие электрогенераторные органы (исключение составляют электрический сом и звездочеты, у которых электрорецепторов нет); ко второй — не имеющие электрических органов (кроме некоторых скатов, осетровых, сомовых и акуловых).
Тем не менее рыбы, у которых отсутствуют электрорецепторы, обладают повышенной электрической чувствительностью по сравнению с другими позвоночными животными. Благодаря ей рыбы в процессе эволюции выработали особые безусловнорефлекторные двигательные реакции на электрические поля — так называемые электротаксисы.
Восприятие рыбами слабых электрических полей
В 1917 г. американские ученые Г. Паркер и В. Гензен изучали чувствительность американского сомика к различным раздражителям. Воздействуя на рыб палочками, изготовленными из стекла, дерева и металла, они обнаружили, что сомики чувствуют приближение металлической палочки на расстоянии нескольких сантиметров, а на стеклянную реагируют только при ее прикосновении. Если поверхность соприкосновения металлической палочки с водой составляла 5—6 см2, рыбы уплывали, а при 0,9—2,8 см2 — они подплывали и «клевали» место контакта металла с водой. На электрически изолированные от воды (покрытые слоем парафина) металлические палочки рыбы реагировали так же, как и на стеклянные,— лишь в момент прикосновения На основании этих наблюдений было сделано предположение, что реакцию сомиков вызывали микротоки, возникающие в результате контакта металла с водой (гальванический эффект).
Дальнейшие эксперименты подтвердили этот вывод. Создавая в воде с помощью электродов и гальванического элемента электрическое поле постоянного тока, Паркер и Гензев обнаружили у рыб такие же реакции, какие вызывали металлические палочки. Токи менее 0,99 мкА вызывали «клев» электродов, больше 1,47 мкА — отрицательную реакцию.
Высокую чувствительность некоторых сомовых рыб к электрическим полям отмечали японские ученые С. Кокубо и К. Узука. При дальнейших исследованиях было установлено, что эти рыбы обладают специализированными электрическими рецепторами.
Иной механизм восприятия электрических полей у рыб, не имеющих электрорецепторов. Электрический ток — универсальный раздражитель нервных и мышечных клеток. Поэтому рыбы реагируют на них при воздействии непосредственно на нервно-мышечные структуры или рецепторы, предназначенные для восприятия неэлектрических раздражителей.
Чувствительность таких рыб к электрическому току исследовалась при изучении их реакций на сильные электрические поля. Ее порог определялся по значению напряжения, при котором наступала первичная двигательная реакция. Однако выяснилось, что этот метод не дает точных данных. Например, у морской лисицы, обладающей чувствительностью в 0,01 мкВ на 1 см, первичная двигательная реакция не совпадает с порогом восприятия электрического поля и проявляется только при напряжении в десятки — сотни вольт. Таким образом, не исключено, что рыбы, не имеющие электрорецепторов, чувствуют более слабые электрические поля.
Следующий этап изучения электрической чувствительности рыб начался с 1958 г., когда Г. Лиссман обнаружил у слабоэлектрических рыб — гимнарха и гимнотуса — особые высокочувствительные электрорецепторы. Стало очевидным, что некоторые рыбы с их помощью ориентируются в окружающей среде. Рыб, имеющих электрорецепторы, стали исследовать многие ученые.
Датские ученые С. Дийкграф и А. Кальмджин в 1962 г. установили, что высокой чувствительностью к электрическим полям переменного тока обладают обыкновенные скаты и акулы. Напряженность поля в десятые доли микровольта на 1 см оказалась достаточной, чтобы вызвать у акул рефлекс смыкания век (моргание глазами) и изменить у морской лисицы ритм дыхания. Выяснилось, что высокая чувствительность этих рыб к электрическим полям обусловлена наличием у них специфических рецепторов — так называемых ампул Лоренцини. При нарушении иннервации их чувствительность понижалась в несколько десятков раз. Ученые пришли к выводу, что ампулы Лоренцини являются у акул и скатов электрорецепторами.
В дальнейших экспериментах исследовалась чувствительность акул и скатов к электрическим полям естественного происхождения. Реакция рыб отмечалась с помощью электрокардиограммы, снимаемой с двух электродов, помещенных в перикард сердца Если рыбы воспринимали электрические поля, мышечный потенциал сердца значительно изменялся — уменьшались его амплитуда и частота следования импульсов. Оказалось, что в благоприятных условиях акулы и скаты чувствительны к электрическим полям напряженностью в сотые доли микровольта на сантиметр, т е. примерно в 10 раз более чувствительны к воздействию электрических полей, чем показывали опыты, проводившиеся другими способами.
При изучении ориентации слабоэлектрических рыб Лиссман установил, что некоторые из них воспринимают исключительно слабые электрические токи, возникающие в теле в результате индукции при движении в магнитном поле. Эта способность позволяет рыбам ориентироваться в таких полях. Гимнотуса, например, можно было приучить кормиться в участке аквариума, вблизи которого находился небольшой магнит. У гнатонемуса и гимнарха движение магнита вне аквариума на некотором расстоянии вызывало характерную двигательную реакцию.
Высокая чувствительность к электрическим полям позволяет слабоэлектрическим рыбам находить и различать в воде объекты, определять соленость воды, использовать разряды других рыб в межвидовых и внутривидовых отношениях. В основном мир ощущений у этих рыб электрический.
У разных видов рыб реакция на внешние электрические поля варьирует в очень широких пределах. Наибольшей чувствительностью обладают представители слабоэлектрических видов: у гимнарха она достигает 0,01 мкВ, стернарха — 0,02 и обыкновенных скатов — 1,2 мкВ на 1 см, а у некоторых сильноэлектрических рыб она значительно меньше: около 1 мВ на 1 см.
Чрезвычайно варьирует чувствительность неэлектрических видов. У некоторых сомовых и осетровых, имеющих специальные электрические рецепторы, она составляет десятки и сотни микровольт на 1 см, а у остальных неэлектрических рыб — десятки милливольт на 1 см.
Чувствительность отдельных участков тела рыб к электрическому току зависит от плотности расположения на них электрических рецепторов. У большинства рыб, имеющих электрические органы, например у гимнарха, такие рецепторы наиболее густо распределены на голове, спине и брюхе, у скатов — на брюхе. Для всех рыб характерно, что там, где находятся электрические органы, электрические рецепторы полностью отсутствуют.
Многочисленные исследования показали, что почти у всех слабоэлектрических и сильноэлектрических рыб специализированными электрорецепторами являются производные органов чувств боковой линии. Наиболее исследована электрорецепторная функция ампул Лоренцини. Этим образованиям присвоено имя итальянского ученого, описавшего их в 1678 г. Согласно его взглядам, эти ампулы — слизистые железы, находящиеся в коже рыб. Они относятся к системе органов чувств боковой линии и встречаются не только у хрящевых рыб (акул и скатов), но и у некоторых костистых рыб (морского тропического сома).
Ампулы Лоренцини (рис. 14) представляют собой довольно длинные (до нескольких сантиметров) трубочки-каналы, заполненные желеобразным веществом и заканчивающиеся на поверхности тела порами; внутри тела они образуют характерное расширение, в котором находятся чувствительные клетки. К каждой ампуле подходит несколько нервных веточек — обычно шесть, но иногда гораздо больше. У мраморного электрического ската их количество колеблется от 26 до 32.
Функциональное назначение ампул Лоренцини долгое время оставалось неясным. Их считали органами, выделяющими слизь, а в конце XIX в.— рецепторами гидростатического давления, предназначенными для ощущения глубины погружения рыбы. Это мнение подтверждалось отсутствием у скатов и акул плавательного пузыря — гидростатического органа костистых рыб. Правда, позднее выяснилось, что у морского тропического сома, обладающего плавательным пузырем, имеются и ампулы Лоренцини.
Электрофизиологические исследования ампул не внесли ясность в этот вопрос. Одни исследователи считали, что функция ампул терморецепторная, категорически отвергая их механорецепторную роль. Другие, наоборот, утверждали, что это механорецепторные органы, воспринимающие разницу давления внутри и снаружи ампул. Было также выдвинуто предположение об их хеморецепторной функции. Обнаружилась высокая чувствительность ампул к изменению концентрации солей в морской воде Появилась гипотеза об электрической чувствительности этих органов.
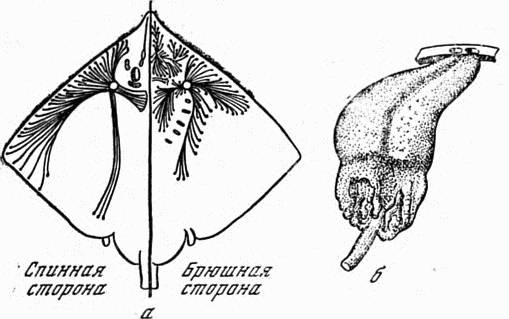
Рис. 14. Ампулы Лоренцини а — расположение ампул Лоренцини на теле морской лисицы (точками на концах трубочек отмечены места, где ампулы открываются наружу), б — две ампулы Лоренцини у акулы (сверху отверстия на коже)
Однако только последующее изучение поведения рыб, в том числе эксперименты Дийкграфа и Кальмджина, о которых уже упоминалось выше, позволили выяснить электрорецепторное назначение ампул Лоренцини. Специальные электрофизиологические исследования подтвердили высокую чувствительность ампул к электрическим полям — приблизительно 0,1 мкВ на 1 см. Если один электрод (зонд) помещался в проток ампулы, а другой — на тело рыбы, то для «срабатывания» ампулы было достаточно действия электрического тока силой всего в 0,005 мА. Если же зонд смещали в сторону от поры ампулы всего на 0,5 мм, то для достижения такого же результата силу тока приходилось заметно увеличивать. Это свидетельствует о том, что электрический ток в основном протекает по трубочке ампулы.
Описываемое явление хорошо согласуется с морфологическими и биофизическими свойствами ампул Лоренцини. Установлено, что электрическое сопротивление стенок ампулы в 160 тыс. раз больше сопротивления желе, заполняющего ее. Электропроводность морской воды и желе в протоках ампулы приблизительно одинакова, остальных частей тела рыбы — примерно в 2 раза меньше. Доказано, что протоки ампул Лоренцини являются хорошо изолированными проводниками, по которым электрический ток передается с небольшими потерями. В связи со специфическим строением ампулы электрические разряды высокой частоты затухают в ее канале быстрее, чем разряды низкой частоты. Поэтому длинные ампулы Лоренцини могут воспринимать в основном низкочастотные разряды, а короткие — высокочастотные Таким образом, ампулы Лоренцини представляют собой электрорецепторы. Однако они чувствительны и к другим раздражителям, прежде всего к механическим воздействиям. Возможно, что эти ампулы являются не только электрорецепторами, но и механорецепторами.
Как уже говорилось, высокой чувствительностью к электрическому току обладают, кроме акул и скатов, слабоэлектрические рыбы, например гнатонемус, гимнарх. У этих рыб имеются электрические рецепторы различных типов, образованные подобно ампулам Лоренцини, из органов чувств системы боковой линии. Наиболее чувствительны к электрическому раздражению ампулярные рецепторы: электрорецепторы гимнарха воспринимают напряженность электрического поля в 0,01 мкВ на 1 см.
Все исследованные электрические рецепторы слабоэлектрических рыб спонтанно генерируют импульсы определенной частоты — осцилляции Они не зависят от разрядов электрических органов рыб и сохраняются даже в том случае, если кусочки кожи, на которых расположены рецепторы, изолированы. Однако по мере приближения к рецепторам объектов, создающих внешнее электрическое поле, частота осцилляций изменяется.
Функции электрических рецепторов слабоэлектрических рыб различны. Одни непрерывно работают как гальванометры (как бы замеряя величину тока); другие же воспринимают только изменение напряжения внешнего электрического поля, определяя его фазу, т. е. работают аналогично осциллографу.
Каким же образом осуществляется анализ информации, поступающей к рыбам извне и воспринимаемой их электрическими рецепторами? Известно, что органами боковой линии рыб управляют мозжечок и продолговатый мозг. Именно в них осуществляется анализ электрической информации. В связи с этим мозжечок у некоторых слабоэлектрических рыб исключительно сильно развит Так, у мормируса относительная величина мозжечка больше, чем у любых других позвоночных животных. Он наиболее развит у сомовых рыб, которые, как и слабоэлектрические рыбы, обладают электрорецепторами. Степень развития «электрических» долей мозга слабоэлектрических и неэлектрических рыб хорошо видна на рис. 15.
У некоторых электрических и слабоэлектрических рыб, обладающих «электрическим мироощущением», относительно развиты боковые доли продолговатого мозга. Изучение этих отделов с точки зрения электрофизиологии позволило выявить несколько типов нейронов, по-разному реагирующих на внешнее электрическое поле, на движение в воде предметов с разной электропроводностью, а также различно адаптирующихся к этим раздражителям Оказалось, что в зависимости от движения объектов активность одних нейронов электрического центра повышалась, а других, наоборот, понижалась. Благодаря этому рыба получала информацию о положении и перемещении в воде разнообразных объектов.
Для передачи любой информации от воспринимающих элементов к анализатору необходим соответствующий код — система условных обозначений. У человека это слова, отдельные фразы, термины, буквы, цифры, знаки и т. д. У рыб подобная система неизмеримо проще, но тем не менее нервные волокна, передающие электрическую информацию от воспринимающих рецепторов в нервные центры, «кодируют» ее разнообразными способами. Это отчасти можно объяснить тем, что электрорецепторы работают в комплексе с электрическими органами.
При одновременной работе электрических органов и электрорецепторов кодирование информации происходит довольно сложным путем. Так, представители гимнотовидных рыб в основном используют два типа кодирования:
«числовое» и «вероятностное». У этих рыб с низкочастотной разрядной деятельностью (3—5 импульсов в секунду) нервные волокна как бы отвечают на каждый импульс цепочкой нервных импульсов. У электрического угря, например, эта цепочка состоит из 15 импульсов в секунду. Кодирование информации об изменении интенсивности внешнего электрического поля осуществляется путем изменения количества импульсов в цепочке — «числовое».
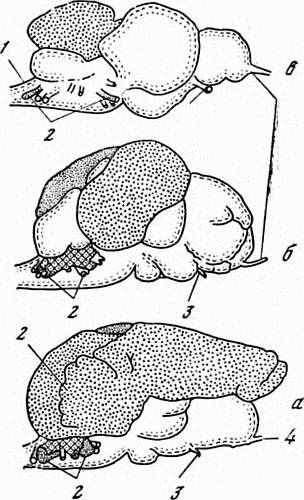
Рис. 15 «Электрические» зоны мозга слабоэлектрических рыб (заштрихованные участки)
а — мормирус,
б — гимнарх,
в — сом,
1 — позвоночный столб,
2 — нерв боковой линии,
3 — зрительный нерв
4 — обонятельный нерв
У гимнотовидных рыб с высокой частотой разрядной деятельности электрических органов обнаружен другой способ кодирования — «вероятностный». При каждом электрическом импульсе импульсы в нервном волокне либо отсутствуют вообще, либо возникает один импульс (у некоторых рыб может возникнуть несколько). Кодирование электрической информации осуществляется в результате возникновения нервного импульса в зависимости от изменения интенсивности внешнего электрического поля.
Есть еще один способ кодирования — «условно-частотный». Нервные волокна проявляют спонтанную активность, не согласованную с разрядами электрического органа. Если во внешнее электрическое поле попадают какие-либо объекты, нервные волокна определенным образом изменяют частоту следования разрядов электрического органа.
В последнее время в СССР и за рубежом были проведены многочисленные исследования, посвященные кодированию электрической информации нервными волокнами. Интерес к этому вопросу закономерен. Зная механизм кодирования, можно понять, как протекает в организме анализ поступающей извне информации и каким образом раздражители различаются во времени и пространстве.
Восприятие рыбами сильных электрических полей
Рыбы воспринимают электрическое поле постоянного тока обычно в виде ориентировочной двигательной реакции (они вздрагивают при включении и выключении тока). При увеличении напряженности поля наступает оборонительная реакция — так называемая стадия отпугивания: рыба сильно возбуждается и пытается выйти из зоны действия поля. Если напряженность увеличить еще больше, происходит анодная реакция. При дальнейшем повышении напряженности наблюдается электронаркоз: рыба теряет равновесие, подвижность и перестает реагировать на внешние раздражители Еще большее повышение напряженности электрического поля вызывает гибель рыбы.
Реакция рыб в электрических полях зависит от их ориентации в электрическом поле. Если рыба расположена головой к аноду, она возбуждается сильнее При постоянном повышении напряженности поля после первой стадии часто наблюдается анодная реакция — рыба движется к аноду.
Несколько иначе рыбы реагируют на электрические поля переменного тока. Первые две стадии примерно те же, но при дальнейшем повышении напряженности наступает стадия осциллотаксиса — рыба располагается поперек линий тока. Еще большее увеличение напряженности вызывает электронаркоз. Переменный ток вызывает у рыб более сильное возбуждение, чем постоянный. После его воздействия рыба долго не может прийти в нормальное состояние — она находится в состоянии своеобразного электрогипноза.
Еще более разнообразно и сложно поведение рыб в полях импульсного электрического тока. Реакции рыб зависят от амплитуды, частоты, формы и продолжительности импульсов. Различные виды рыб реагируют на импульсные поля неодинаково, однако и в этом случае стадии реакций у них такие же, как при воздействии полей постоянного тока.
Проявление отдельных стадий реакций рыб на различные электрические поля зависит от условий среды (электропроводности, температуры), а также от видовой чувствительности[5] рыб к току, их размеров, формы и физиологического состояния.
Итак, при действии на рыб сильных электрических полей можно выделить несколько типичных стадий изменения поведения: первичная пороговая реакция, возбуждение, анодная реакция и электрический наркоз (шок). Примерно по такой же схеме действует возрастающий электрический ток на любой нервно-мышечный аппарат. Это совпадение не случайно. Оно, несомненно, свидетельствует, что поведенческие реакции рыб на сильные электрические поля основаны на процессах в нервно-мышечных элементах. Сильные электрические поля воспринимаются этими элементами рыбы. В результате нарушения их нормальной работы и принудительного сокращения мускулатуры возникает та или иная реакция рыбы.
Однако механизм направленного движения рыб на анод не выяснен. Существуют только гипотезы. Остановимся на некоторых из них[6].
Первая гипотеза связана с представлением о перераспределении в теле рыб ионов под воздействием электрического поля. В обычном состоянии ионы не «рассортированы». Когда же на рыб начинает действовать электрическое поле, отрицательные ионы группируются в сторону положительного электрода. О перераспределении ионов в теле рыб говорят факты остаточного напряжения после снятия действия внешнего электрического поля. Накопление отрицательных ионов в голове рыбы заставляет рыб двигаться к аноду. В этой гипотезе не все ясно. Непонятно, например, почему рыба движется всегда головой к аноду. Очень часто она в начале действия поля головой повернута в противоположную сторону. В этом случае отрицательные ионы должны были бы накапливаться в хвосте, и рыба должна была бы двигаться хвостом к аноду. Между тем рыба всегда движется головой к аноду.
Согласно другим гипотезам, рыба — генератор биоэлектричества. Биоэлектрическое поле рыбы определенным образом взаимодействует с внешним электрическим полем. В результате у рыбы возникает анодная реакция.
Общие положения гипотез приемлемы. Однако есть моменты, с которыми нельзя согласиться. Так, предполагается, что ток в теле рыб (имеются в виду неэлектрические рыбы) течет от головы к хвосту. Между тем это положение не подтверждается экспериментально. Кроме того, согласно гипотезам, взаимодействие между полями начинается, когда рыба попадает в электрическое поле. При малом напряжении у рыб нет стремления изменить свое положение. Но с повышением напряженности внешнего поля это взаимодействие для рыбы становится ощутимым. В случае течения тока в противоположных направлениях результирующая сила тока уменьшается, и рыба для восстановления электрического равновесия меняет свое положение. Видимо, некоторое увеличение силы тока оказывает на рыбу действие, направляющее ее в сторону анода.
Согласно этим гипотезам, рыба движется к аноду, если напряженность внешнего электрического поля складывается с биоимпульсами ее мышц. В другом случае взаимодействие вызывает отрицательную реакцию.
Анодная реакция, как известно, начинается при очень высокой напряженности внешнего электрического поля — более 50 мВ на 1 см. Напряженность внешних электрических полей, вызывающих анодную реакцию, в 100—10 000 раз выше напряженности биоэлектрических полей. Почему же более слабые электрические поля не вызывают анодной реакции?
Взаимодействие между сильными внешними электрическими и биоэлектрическими полями рыб действительно существует. Но восприятие их рыбами принципиально иное, чем восприятие слабых полей. В сильных электрических полях реакции рыб по своей природе безусловно-рефлекторны. Слабые электрические поля рыбы могут использовать в целях ориентации и общения.
Эволюция электрических систем
Как уже отмечалось, из всех известных животных только некоторые виды рыб в ходе эволюции развили способность создавать сильные электрические поля и использовать их в специальных целях. Концевые двигательные пластинки мышц у этих рыб превратились в специализированные электрические генераторы — электрические пластинки.
Интересно, что электрические органы есть как у низших хрящевых рыб (скатов), так и у представителей отрядов и семейств костистых рыб У некоторых костистых и хрящевых рыб имеются также электрические рецепторы и соответствующие анализаторные системы. Однако прямой взаимосвязи в развитии электрических органов и электрических рецепторов не наблюдается Так, у некоторых видов рыб, имеющих электрические органы (электрический сом, звездочеты), нет электрических рецепторов; у других видов, не обладающих морфологически оформленными электрическими органами (некоторые из осетровых, сомовых и акул), такие рецепторы есть.
Каким же образом произошли электрические органы и электрические рецепторы рыб в процессе естественного отбора? Ответить на этот вопрос трудно. Прежде всего, почему электрическими органами обладают только рыбы? Это, казалось бы, можно объяснить их экологией — тем, что рыбы живут в среде, имеющей сравнительно высокую электропроводимость. Но в воде обитают не только рыбы, но и другие водные организмы (например, амфибии и беспозвоночные), у которых таких органов нет. Очевидно, эволюционные механизмы определяются не только условиями окружающей среды, но и предрасположенностью к соответствующей трансформации клеток и тканей.
Однако исходный материал, из которого развились электрические органы,— мышечная ткань — широко представлен в других классах животных. Если мы сравним мышцы рыб и мышцы, например, беспозвоночных и особенно амфибий, то не сможем определить причину, обусловившую преобразование мышечной ткани рыб в электрические органы. Концевые двигательные пластинки рыб, ставшие объектом трансформации, по своему морфологическому строению такие же, как у амфибий и водных беспозвоночных. Правда, мышечные клетки расположены в туловище и хвосте рыб в определенном порядке, благодаря чему их преобразование в серии электропластинок облегчается. Но в других мышцах рыб такое расположение клеток не встречается, хотя электрические органы ската произошли из жаберных мышц, американского звездочета — из глазных.
Единственное отличие мускулатуры рыб от мускулатуры амфибии и водных беспозвоночных — ее развитие (количественный признак). Мышечная ткань у амфибий сравнительно однородна и мало развита, а у водных беспозвоночных — еще менее развита. Образно говоря, у этих организмов отсутствует «лишняя» мышечная ткань, имеющаяся у рыб. Двигательные мышцы рыб выполняют не только двигательную функцию, они участвуют также в обмене веществ. Все сказанное свидетельствует о предрасположенности мышц рыб к генетическим преобразованиям, которой нет ни у беспозвоночных, ни у амфибий.
Рассмотрим происхождение электрических органов. Как уже отмечалось, они имеются и у хрящевых и у костистых рыб.
Если принять степень разнообразия электрических систем в различных группах за критерии длительности их развития, то наиболее древние по происхождению электрические системы имеются у гимнотовидных, затем у мормирообразных и, наконец, у скатов, хотя скаты — наиболее древняя группа среди этих групп. Поэтому можно предположить, что электрические органы скатов являются новообразованием. Кроме того, различен исходный мышечный субстрат, из которого развились электрические органы: у электрических скатов — это мышцы жаберного аппарата, у американского звездочета — глазные, у мормирообразных и обыкновенных скатов — хвостовые, у электрического угря — туловищные.
Как же объяснить это разнообразие в происхождении электрических органов?
Еще Ч. Дарвин признавал, что существование электрических органов даже у сравнительно небольшого числа разновидностей, известных в его время, обусловливает одну из «особых трудностей теории естественного отбора».
Исходя из теории естественного отбора полезных для вида признаков и свойств Дарвин объяснил существование электрических органов у электрических угря и ската тем, что они необходимы этим рыбам в агрессивно-оборонительных целях. Существование же электрических органов у слабоэлектрических рыб казалось необъяснимым, так как во времена Дарвина локационное и коммуникационное назначение этих органов было неизвестно.
Некоторые современники Дарвина рассматривали слабоэлектрических рыб как предшественников настоящих электрических. На ошибочности такой классификации еще тогда настаивал русский ученый А. И. Бабухин. По сравнению с сильноэлектрическими у слабоэлектрических костистых рыб наблюдается большое разнообразие анатомических и физиологических особенностей, что указывает на их более высокий уровень развития. Слабоэлектрические рыбы ориентируются благодаря развитию нервной системы, анализирующей информацию, поступающую от рецепторов и элементарных электрогенераторов. У слабоэлектрических рыб отдельная электропластинка генерирует более высокое напряжение, чем у сильноэлектрических.
Формирование электрических органов зависит не только от генетических механизмов, но и от условий окружающей среды. Морская вода благодаря содержанию в ней солей имеет сравнительно высокую электропроводимость, а пресная — гораздо более низкую. Известно, что по проводнику с высокой электропроводностью целесообразна передача тока большой силы, а по проводнику с низкой электропроводностью — тока высокого напряжения. Именно этот принцип определяет устройство электрических органов морских и пресноводных рыб. Электрические пластинки морских рыб соединены параллельно, благодаря чему их электрические органы генерируют импульсы большой силы тока, но невысокого напряжения Наоборот, электрические пластинки пресноводных рыб соединены последовательно, и их электрические органы генерируют импульсы высокого напряжения при небольшой силе тока. Именно в этих особенностях устройства и функционирования электрических органов проявилась адаптация рыб к окружающей среде.
Другое приспособление к условиям среды заключается в биологическом назначении электрических систем. В связи с этим в настоящее время рыб подразделяют на три группы: сильноэлектрические, слабоэлектрические первого и второго родов. Эта классификация в значительной мере основана на системном подходе, при котором анализируется развитие не только электрических органов, но и электрических сенсорных (чувствительных) систем. Такой подход позволяет рассматривать вопрос о происхождении электрических систем у рыб с большей точностью.
«Исходным материалом» в процессе развития сильно-и слабоэлектрических рыб первого рода могли быть неэлектрические виды рыб (слабоэлектрические рыбы второго рода), обладающие способностью генерировать слабые разряды с помощью нервно-мышечных структур. Таким образом, уже неэлектрические рыбы имеют потенциальную возможность развития этой способности для использования ее в специфических целях.
В некоторых межвидовых и внутривидовых актах поведения (особенно в стайном поведении) суммирование разрядов отдельных рыб, составляющих стаю, позволяет им пользоваться особой стайной формой электроориентации Суммированные разряды достигают величин, при которых они могут быть восприняты неспециализированными рецепторами (см. стр. 66).
Стайность — это наиболее древняя форма поведения, поэтому систему стайной электроориентации, по-видимому, можно считать первой электросистемой, возникшей у рыб. Очевидно, подобная система наиболее развилась у типично стайных рыб, обитающих в мутных неспокойных водах (например, у проходных каспийских сельдей) и в водах с низкой электропроводимостью. Возможно, что такая форма электроориентации способствовала появлению у отдельных стайных видов (некоторые гимнотовидные) электрорецепторов.
Сильноэлектрические виды рыб, как правило, используют свои разряды для нападения или обороны. Поэтому следует думать, что их электрические системы возникли у одиночных хищников, находившихся в условиях жесткой межвидовой конкуренции, или у хищников-засадчиков. Не исключено, что звездочеты в этом отношении представляют переходную ступень между сильноэлектрическими и неэлектрическими видами рыб. Рыбы этого семейства — хищники-засадчики. У них сформированы электрические органы, но электрорецепторов нет. Захват добычи звездочеты сопровождают характерной разрядной деятельностью, аналогичной той, какая присуща электрическому сому. Однако их разряды имеют небольшую величину и поэтому не могут парализовать добычу.
Разряды слабоэлектрических рыб первого рода в основном предназначены для локации. Естественно, что она может осуществляться только с помощью электрических рецепторов. У рыб они представляют собой производные органов чувств системы боковой линии. Эта система является механорецепторной. Ее рецепторы характеризуются полифункциональностью — они воспринимают давление, вибрации и, как считают некоторые исследователи, относительно сильные электрические поля.
Как же осуществлялось преобразование рецепторов боковой линии в электрические? Ответить на этот вопрос очень важно, так как именно развитие электрических рецепторов у какой-то предэлектрической рыбы повлекло за собой образование и развитие электрических органов.
В процессе эволюции, очевидно, была стадия, когда рецепторы боковой линии одновременно воспринимали как механические, так и электрические раздражители. Это подтверждают исследования, показывающие, что в ординарных рецепторах боковой линии неэлектрических рыб может возникать так называемый микрофонный эффект. Такая особенность свидетельствует о принципиальной возможности трансформации механочувствительных систем рыб в электрочувствительные.
Итак, первым шагом в процессе перехода от неэлектрических рыб к электрическим явилось приобретение отдельными видами рецепторов, чувствительных к электрическим раздражителям. На первой стадии эта чувствительность может быть неспецифической, как, например, У некоторых осетровых и сомовых, имеющих пассивные электрические рецепторы. Параллельно шло развитие электрических органов из мышечной ткани.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 13. Рецепторы и органы чувств
§ 13. Рецепторы и органы чувств Органы чувств по источникам воздействий можно разделить на эндогенные и экзогенные. Первые специализируются на рецепции внутренней среды и органов животного, а вторые информируют о внешней среде. Оба источника информации крайне важны для
4.2. Электрические явления в вестибулярной системе
4.2. Электрические явления в вестибулярной системе Даже в полном покое в волокнах вестибулярного нерва регистрируется спонтанная импульсация. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую сторону (детекция
ОПИАТНЫЕ РЕЦЕПТОРЫ И ОПИОИДЫ МОЗГА
ОПИАТНЫЕ РЕЦЕПТОРЫ И ОПИОИДЫ МОЗГА Морфин и кодеин – два активных ингредиента морфия. Морфин был очищен в XIX веке и стал широко применяться в медицине. Механизм действия морфина на мозг довольно хорошо изучен благодаря тому, что было синтезировано вещество налоксон,
Рецепторы гормонов
Рецепторы гормонов Биологическое действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней. Клетки, наиболее чувствительные к влиянию определенного гормона, называют клеткой-мишенью. Специфичность гормонов по отношению к клеткам-мишеням
Механизм передачи гормональных сигналов через мембранные рецепторы
Механизм передачи гормональных сигналов через мембранные рецепторы Гормоны (первичные посредники) связываются с рецепторами на поверхности клеточной мембраны и образуют комплекс гормон-рецептор. Этот комплекс трансформирует сигнал первичного посредника путем
Механизм передачи гормонального сигнала через внутриклеточные рецепторы
Механизм передачи гормонального сигнала через внутриклеточные рецепторы Передача сигнала гормонов с липофильными свойствами (стероидные гормоны) и тироксина возможна при прохождении их через плазматическую мембрану клеток-мишеней. Рецепторы гормонов находятся в
Передача сигналов через рецепторы, сопряженные с ионными каналами
Передача сигналов через рецепторы, сопряженные с ионными каналами Рецепторы, сопряженные с ионными каналами, являются интегральными мембранными белками, состоящими из нескольких субъединиц. Они действуют одновременно как ионные каналы и как рецепторы, которые
Как устроены электрические органы рыб?
Как устроены электрические органы рыб? Известно более 300 видов современных рыб, оснащенных электрическими органами – парными образованиями, способными генерировать электрические разряды, которые служат для защиты, нападения, внутривидовой сигнализации, ориентации и
Электрические и магнитные поля
Электрические и магнитные поля Из того перечня сил природы, которые могут участвовать в строительстве и осуществлении жизни тел организмов и формируют нас, людей (как личность — как сущность, которая имеет возможность хранить информацию и перерабатывать ее, а также
Электрические разряды рыб
Электрические разряды рыб По характеру генерируемых разрядов различают два типа рыб. К первому относят рыб, использующих электрические органы для обороны и нападения — разряды производятся только в ответ на стимуляцию или при встрече с жертвой. Это все
Электрические рыбы и бионика
Электрические рыбы и бионика Идеи различных изобретений человек, как правило, находил в окружающей его природе. Так, в первых проектах летательных аппаратов копировалось крыло птицы или летучей мыши. К изобретению самозатачивающихся инструментов привело исследование