7. Парадокс Симпсона
7. Парадокс Симпсона
Еще одна замечательная хитрость такого рода называется парадоксом Симпсона. Суть этого парадокса в том, что при соблюдении определенного набора условий частота встречаемости альтруистов в группе популяций будет расти, несмотря на то, что внутри каждой отдельной популяции эта частота неуклонно снижается.
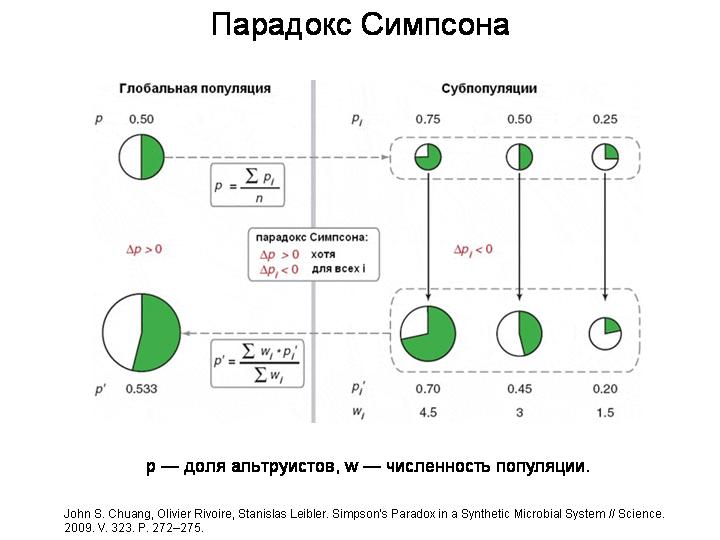
На этом слайде показан гипотетический пример действия «парадокса Симпсона». В исходной популяции было 50 % альтруистов и 50 % эгоистов (кружок слева вверху). Эта популяция разделилась на три субпопуляции с разным соотношением альтруистов и эгоистов (три маленьких кружка справа вверху). В ходе роста каждой из трех субпопуляций альтруисты оказались в проигрыше — их процент снизился во всех трех случаях. Однако те субпопуляции, в которых изначально было больше альтруистов, выросли сильнее благодаря тому, что они имели в своем распоряжении больше «общественно-полезного продукта», производимого альтруистами (три кружка справа внизу). В результате, если сложить вместе три выросших субпопуляции, мы увидим, что «глобальный» процент альтруистов вырос (большой кружок слева внизу).
О том, что такой механизм в принципе возможен, в свое время говорили Холдейн и Гамильтон, которых я уже упоминал как создателей теории родственного отбора. Однако получить экспериментальные доказательства действенности парадокса Симпсона удалось лишь недавно.
Это было трудно сделать, потому что в каждом конкретном случае, когда мы видим распространение «генов альтруизма» в популяции, очень нелегко доказать, что здесь не замешаны какие-то другие, неизвестные нам выгоды, сопряженные с альтруизмом у данного вида организмов.
Чтобы выяснить, может ли «парадокс Симпсона» в одиночку обеспечить процветание альтруистов, американские биологи создали интересную живую модель из двух штаммов генетически модифицированных кишечных палочек.
В геном первого из двух штаммов («альтруисты») был добавлен ген фермента, синтезирующего сигнальное вещество N-ацил-гомосерин-лактон, используемое некоторыми микробами для химического «общения» друг с другом. Кроме того, в геном обоих штаммов был добавлен ген фермента, обеспечивающего устойчивость к антибиотику хлорамфениколу. К этому гену «приделали» такой промотор (регуляторную последовательность), который активизирует работу гена только в том случае, если в клетку извне поступает вышеупомянутое сигнальное вещество.
«Эгоисты» ничем не отличались от альтруистов, кроме того, что у них не было гена, необходимого для синтеза сигнального вещества.
Таким образом, сигнальное вещество, выделяемое альтруистами, необходимо обоим штаммам для успешного роста в присутствии антибиотика. Выгода, получаемая обоими штаммами от сигнального вещества, одинакова, но альтруисты тратят ресурсы на его производство, а эгоисты живут на готовеньком.
Поскольку оба штамма были созданы искусственно и не имели никакой эволюционной истории, экспериментаторы знали наверняка, что никаких «тайных хитростей» во взаимоотношениях альтруистов с эгоистами в их модели нет, и альтруисты не получают от своего альтруизма никаких дополнительных выгод.
В среде с добавлением антибиотика чистые культуры эгоистов, как и следовало ожидать, росли хуже, чем чистые культуры альтруистов (поскольку в отсутствие сигнального вещества ген защиты от антибиотика у эгоистов оставался выключен). Однако они начинали расти лучше альтруистов, как только в среду добавляли либо живых альтруистов, либо очищенное сигнальное вещество. Альтруисты в смешанной культуре росли медленнее, потому что им приходилось тратить ресурсы на синтез сигнального вещества. Убедившись, что модельная система работает в соответствии с ожиданиями, исследователи приступили к моделированию «парадокса Симпсона».
Для этого они посадили в 12 пробирок со средой, содержащей антибиотик, смеси двух культур в разных пропорциях (0, 10, 20, 30, 40, 50, 60, 70, 80, 90, 95 и 100 % альтруистов соответственно), подождали 12 часов, а затем измерили численность бактерий и процент альтруистов в каждой пробирке. Оказалось, что во всех пробирках, кроме 1-й и 12-й, процент альтруистов значительно снизился. Таким образом, альтруисты во всех случаях проигрывали конкуренцию эгоистам. Однако размер тех популяций, где изначально было больше альтруистов, вырос значительно сильнее, чем тех, где преобладали эгоисты. Когда авторы суммировали численности микробов во всех 12 пробирках, то выяснилось, что общий процент альтруистов заметно вырос: парадокс Симпсона успешно «сработал».
Однако в природе никто не будет нарочно смешивать альтруистов с эгоистами в разных пропорциях и рассаживать их по пробиркам. Какой природный процесс может служить аналогом такой процедуры? Авторы показали, что эту роль могут играть «бутылочные горлышки» — периоды сильного сокращения численности популяции с последующим ее восстановлением. Это может происходить, например, при заселении новых субстратов очень небольшим числом микробов — «основателей». Если число основателей невелико, то среди них в силу простой случайности может оказаться повышенный процент альтруистов. Популяция, которую образует эта группа основателей, будет расти быстро, тогда как другие популяции, основанные группами микробов с преобладанием эгоистов, будут расти медленно. В итоге парадокс Симпсона обеспечит рост «глобальной» доли альтруистов в совокупности всех популяций.
Чтобы доказать действенность этого механизма, авторы смешали альтруистов с эгоистами в равной пропорции, сильно разбавили полученную культуру и стали ее высевать в пробирки порциями разного объема, с приблизительно известным числом микробов в каждой порции. Размер порций оказался главным фактором, от которого зависела дальнейшая судьба альтруистов. Как и следовало ожидать, когда порции были большими, парадокс Симпсона не проявился. В большой порции, то есть в большой выборке из исходной культуры, соотношение альтруистов и эгоистов по законам статистики не может сильно отличаться от исходного. Популяции, основанные этими выборками, растут примерно с одинаковой скоростью, и альтруисты оказываются в проигрыше не только в каждой популяции по отдельности, но и во всех популяциях в целом.
Однако если порции были настолько малы, что в каждой было всего несколько бактерий, то среди этих порций обязательно оказывались такие, в которых альтруисты преобладали. Такие группы основателей давали начало быстро растущим колониям, и за счет этого общий процент альтруистов в совокупности всех популяций увеличивался. В конкретных условиях данного эксперимента для проявления эффекта Симпсона необходимо, чтобы среднее число микробов в группе основателей было не больше 10.
Авторы также показали, что, повторив несколько раз эту последовательность действий (разбавление культуры, расселение маленькими группами в пробирки, рост, соединение популяций в одну, опять разбавление и т. д.) можно добиться сколь угодно высокого процента альтруистов в культуре.
Было выявлено и еще одно условие, необходимое для распространения «генов альтруизма» в модельной системе: смешанным популяциям нельзя позволять расти слишком долго. Разбавление и расселение нужно проводить до того, как популяции достигнут стабильного уровня численности, заселив всю питательную среду в пробирке, потому что тогда различия по уровню численности между популяциями сглаживаются, и парадокс Симпсона не может проявиться.
Таким образом, естественный отбор при соблюдении определенных условий может обеспечивать развитие альтруизма даже тогда, когда в каждой отдельно взятой популяции он благоприятствует эгоистам, а альтруистов обрекает на постепенное вымирание. Однако спектр условий, в которых может действовать парадокс Симпсона, довольно узок, и поэтому в природе он вряд ли играет очень большую роль.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Парадокс веществ-разобщителей
Парадокс веществ-разобщителей Тот факт, что окисление может быть отключено от фосфорилирования, впервые описан при изучении брожения. Если сбраживать сахар в среде, где фосфат (Н3РО4) заменен на арсенат (H3AsO4), то брожение идет с большей скоростью, но без образования АТФ.
Парадокс оленей: отец силен, а дочь слабая
Парадокс оленей: отец силен, а дочь слабая «Почти как отец» — подобная характеристика радует в детском возрасте, но для дочери, по мере взросления, такой отзыв становится не совсем приятен. Кому может понравиться женщина с широкими плечами, сильным подбородком, низким
2.3. Временной парадокс
2.3. Временной парадокс Каким образом результат (событие, которое наступит в будущем) может детерминировать текущую активность, быть её причиной? Решением этого «временного парадокса» была разработка представления об «информационном эквиваленте результата», о модели
Глава 96. Парадокс Ферми (XV)
Глава 96. Парадокс Ферми (XV) Сообразительному Читателю уже не нужны пояснения Автора относительно номера этой главы. Читатель уверен, что 96 — это своеобразная кода, возвращение к началу, символ описанных выше цифровых симметрий. Или попытка Автора придать своему нынешнему
Глава 5. Парадокс гениев
Глава 5. Парадокс гениев Гениальность – что это такое? Великий дар, заслуженная награда или же патология? А может быть, гениальный ум – это компенсация за физические недостатки или издержи развития? Как ни парадоксально, но многие гении имели серьезные проблемы со
Глава 96. Парадокс Ферми (XV)
Глава 96. Парадокс Ферми (XV) Сообразительному Читателю уже не нужны пояснения Автора относительно номера этой главы. Читатель уверен, что 96 – это своеобразная кода, возвращение к началу, символ описанных выше цифровых симметрий. Или попытка Автора придать своему
Глава 11 Эукариотический геном и парадокс значений С
Глава 11 Эукариотический геном и парадокс значений С У нас нет достаточных оснований допускать, что эволюция происходит прежде всего за счет увеличения числа генов у высших форм ... главную роль играет не число появляющихся новых генов, а их виды. Т. Морган. «Научные основы
Cамый большой парадокс лингвистики
Cамый большой парадокс лингвистики «…Никто поначалу не обращал внимания на этих пришельцев. Слишком хрупкие, чтобы в одиночку охотиться на крупную дичь. Слишком уязвимые, чтобы противостоять нам в единоборстве – их длинные шеи ломались под натиском наших мощных рук. Их
Глава 8 Парадокс измены
Глава 8 Парадокс измены Мы не удивимся, если окажется, что в этот момент вы чешете затылок и спрашиваете себя: «Раз мы так зависим друг от друга и до смерти боимся расстаться, почему же отношения заканчиваются? И как объяснить измену жены или мужа?» Эти два интереснейших