Родственный отбор
Родственный отбор
Кооперация и альтруизм — краеугольные камни социального поведения Homo sapiens. Вряд ли нужно объяснять, что без кооперации (сотрудничества, взаимовыгодного поведения) устойчивое существование социума у приматов невозможно. Очевидно и то, что наши предки эволюционировали именно как социальные, общественные животные. Социальная жизнь часто предполагает и некоторую долю альтруизма, жертвенности. Это не абсолютное правило, но в эволюции гоминид оно, по всей видимости, соблюдалось неукоснительно. Уже 1,7 млн лет назад ранние Homo заботились о беззубых стариках (см. главу "Очеловечивание" в кн. 1). А ведь это альтруизм — кормить беспомощного старика, вместо того чтобы съесть эту пищу самим или отдать своим детям.
Поэтому мы не поймем эволюцию человека, пока не разберемся в эволюционных механизмах, ведущих к развитию кооперативного и альтруистического поведения. Это тем более важно, что до сих пор еще приходится иногда слышать от далеких от биологии людей, что эволюция якобы не может объяснить альтруизм. На самом деле эволюция превосходно его объясняет. Каким образом — в этом мы сейчас попробуем разобраться.
Изучение эволюции альтруизма и кооперации — это одно из тех направлений, двигаясь по которым биология — естественная наука — вторгается на территорию, где до сих пор безраздельно хозяйничали философы, теологи и гуманитарии. Неудивительно, что вокруг эволюционной этики (так называют это научное направление) кипят страсти. Об этих страстях мы говорить не будем, потому что они кипят за пределами естественных наук. Пусть себе кипят. Нас, биологов, интересует другое. Нас интересует, почему, с одной стороны, большинство живых существ ведут себя эгоистично, но при этом немало есть и таких, кто совершает альтруистические поступки, то есть жертвует собой ради других.
Альтруизм у дрожжей? Да как вы смеете!
Часто приходится слышать, что такие понятия, как альтруизм (а еще мышление, желание, планирование, понимание, любовь и многие-многие другие), применимы только к человеку и больше ни к кому. По-моему, это просто дело привычки и общественной договоренности. Главное — чтобы было понятно, о чем речь. Слова для людей, а не люди для слов. Было время, даже слово "поведение" кое-кто считал допустимым применять только к людям. Ну как же: ведь только люди могут вести себя осознанно, осмысленно, то есть по-настоящему. А остальные живые существа себя не осознают, личности у них нет, и поэтому никак "себя вести" они не могут. Сейчас такие рассуждения выглядят историческим курьезом — все давно привыкли, что у нечеловеческих животных тоже есть поведение. Иначе можно договориться до того, что только люди могут есть: ведь мы едим осознанно, а все остальные — неосознанно.
Употребление слова "альтруизм" по отношению к нечеловеческим живым существам имеет в биологической литературе давнюю историю. Но все-таки многие еще не успели к этому привыкнуть. Поэтому нужно немного разобраться с определениями, тем более что общепринятый смысл понятия "альтруизм" в применении к людям (в этике) и другим живым существам (в биологии) не совсем одинаков.
АЛЬТРУИЗМ (ЛАТ. ALTER — ДРУГОЙ) В ЭТИКЕ — нравственный принцип, предписывающий бескорыстные действия, направленные на благо других людей; способность приносить свою выгоду в жертву ради общего блага.
АЛЬТРУИЗМ В БИОЛОГИИ: поведение, ведущее к повышению приспособленности (репродуктивного успеха) других особей в ущерб своим собственным шансам на успешное размножение.
В этих определениях много общего. В обоих случаях имеется в виду принесение индивидом в жертву своих интересов ("выгоды") ради кого-то другого. Однако интересы могут быть разные. В случае с человеком подразумевается, что у индивида есть какие-то цели, которые он в той или иной степени осознает (и может быть, даже сам себе их поставил, хотя что это значит — "сам поставил" — вопрос очень каверзный; см. раздел "Свободная воля" в главе "В поисках душевной грани"). Причем здесь имеются в виду не любые цели, а корыстные, эгоистические, связанные с личной выгодой (благами, удовольствиями). И наш индивид вдруг берет и отказывается от осуществления своей корыстной цели — жертвует своей "выгодой" ради кого-то другого. Вот вам и альтруизм.
Это определение (как и любое другое) можно при желании свести к абсурду и разрушить всю его слабенькую логику, если начать детализировать и докапываться: а что такое благо, а что такое выгода, а что такое удовольствие, а не получает ли человек удовольствие, совершая добрый поступок, а если получает, то это уже не альтруизм, и т. д.
Центральную роль в принятии решений у нас все-таки играют подкорковые структуры с их контурами боли и награды, кнута и пряника. Кора больших полушарий выполняет сложные калькуляции, строит модели, прогнозирует последствия возможных поступков, сравнивает их с последствиями альтернативных вариантов поведения. Но эмоциональная оценка результатов всех этих вычислений зависит от подкорковых структур. Это они в конечном счете определяют, что делать следует, а что нет. Какой из вариантов даст в итоге больше пряников-эндорфинов. Хвостатое ядро капризно виляет хвостиком: хочу — не хочу, буду — не буду. Если вам хочется совершить добрый поступок (и получить на свою голову все его предполагаемые последствия) сильнее, чем хочется его не совершать (тоже со всеми вытекающими), то вы его совершите. Если мысленная модель планируемого доброго поступка активирует нейронную сеть "кнута" сильнее, чем нейронную сеть "пряника", то вы его не совершите.
Так что при желании можно привести аргументы в поддержку тезиса, что альтруизма вообще не бывает: что хотим, то и делаем. Можно порассуждать о том, что моральные нормы, законы, системы запретов и табу как раз для того и выработаны обществом, чтобы наши хвостатые ядра чаще склоняли хвост в сторону поступков, полезных окружающим. Но лучше не будем погружаться в эти демагогические пучины. Остановимся на том, что большинству людей интуитивно понятно, что такое "эгоистический интерес", когда дело касается человека.
Перейдем к биологическому определению. Какие цели, какие интересы могут быть у дрожжевой клетки? Строго говоря, никаких интересов у нее нет и целей тоже. Но нам удобно думать, что у нее есть одна, совершенно определенная цель: размножиться, растиражировать свои гены в следующих поколениях. Нам удобно приписать ей такую цель, потому что если приглядеться внимательно к любому живому существу, то мы увидим, что оно замечательно приспособлено для выполнения именно этого действия — размножения, тиражирования своих генов.
Так получается само собой, потому что так работает естественный отбор. Если случайная мутация повышает эффективность тиражирования данного фрагмента ДНК, то (как можно догадаться) данный фрагмент ДНК будет тиражироваться эффективнее. Причем, что важно, тиражироваться он будет вместе с этой самой мутацией, которая повысила эффективность его размножения. Число копий гена с такой мутацией будет расти. Если другая мутация снизит эффективность размножения своего кусочка ДНК, этот кусочек — вместе с этой мутацией — будет размножаться хуже, и число его копий в этом мире будет снижаться.
В результате мир неуклонно наполняется фрагментами ДНК, все более эффективно себя тиражирующими. Ни у кого нет никаких целей. Однако выглядит все так, как будто у фрагментов ДНК (генов) есть цель: размножаться как можно эффективнее. На самом деле они просто автоматически, в силу слепых законов природы, накапливают изменения, повышающие эффективность их размножения. Но нам — в силу некоторых особенностей нашей психики — удобно рассматривать этот процесс как целенаправленный.
В действительности он просто направленный, без "целе-". Естественный отбор придает ему направленность в сторону роста эффективности размножения генов. А цель ему приписываем мы. Нам так удобно, потому что мы так устроены: мы любим приписывать цели направленным процессам, результаты которых можно в какой-то мере предвидеть. Или объектам, от которых можно ожидать более-менее определенных действий.
Философ Дэниел Деннетт объяснил, почему это нам удобно, в книге The Intentional Stance ("Интенциональная установка"). Это ценная психическая адаптация, удобное "срезание угла", кратчайший путь к практичному выводу, лихой прыжок через длинную цепочку "лишних" логических рассуждений — нейронных калькуляций. Если в джунглях на нас бросается хищник, глупо размышлять о природе его поведенческих реакций, о его рецепторах, лимбической системе и гормональном статусе. "Он хочет меня сожрать!" — вот как должен осмыслить поведение хищника жизнеспособный, адаптированный к реалиям этого мира двуногий примат. В некоторых языках даже будущее время формируется на основе глагола "хотеть": говорят не "поезд придет через пять минут", а "поезд хочет прийти через пять минут".
Нам удобно использовать интенциональную установку, потому что у нас великолепно развита "теория ума". У нас есть хорошая мысленная модель себя, и нам легко судить о внешних объектах по себе. А мы-то сами постоянно занимаемся планированием и ставим себе цели (по крайней мере так нам кажется).
Раз нам это удобно, то давайте так и поступим. Будем считать понарошку, что у генов и организмов есть цель — как можно эффективнее размножиться. Пусть это будет их "корыстным интересом". А когда организм вдруг начинает вести себя в ущерб своему корыстному интересу, да притом еще так, что это идет на пользу интересам другого организма, мы будем называть такое поведение альтруистическим. Вот и ладно, вот и договорились.
Итак, перед нами стоят два основных вопроса. С одной стороны, ясно, что многие жизненные задачи легче решать совместными усилиями, чем в одиночку. Почему же тогда биосфера так и не превратилась в царство всеобщей дружбы и взаимопомощи? Это первый вопрос.
Второй вопрос противоположен первому. Как вообще может в ходе эволюции возникнуть альтруизм, если движущей силой эволюции является естественный отбор — процесс, как представляется на первый взгляд, абсолютно эгоистический?
Все дело в том, что этот "первый взгляд" — неправильный. Ошибка здесь в смешении уровней, на которых мы рассматриваем эволюцию. Эволюцию можно рассматривать на разных уровнях: генов, индивидов, групп, популяций, экосистем, всей биосферы. На каждом уровне свои закономерности и правила. На уровне генов в основе эволюции лежит конкуренция разных вариантов (аллелей) одного и того же гена за доминирование в генофонде популяции. На генном уровне никакого альтруизма нет и быть не может. Ген всегда эгоистичен. Если появится "добрый" аллель, который в ущерб себе позволит размножаться другому аллелю, то этот альтруистический аллель будет вытеснен из генофонда и просто исчезнет.
Но если мы переведем взгляд с уровня генов на уровень организмов, то картина будет уже другой. Потому что интересы гена не всегда совпадают с интересами организма. Ген, или, точнее, аллель, — это не единичный объект, он присутствует в генофонде в виде множества одинаковых копий. "Интерес" у всех этих копий один и тот же. Ведь они — просто молекулы, и они абсолютно идентичны. И им, и нам, и естественному отбору совершенно все равно, какая именно из одинаковых молекул размножится, а какая нет. Важен только суммарный итог: сколько копий аллеля было и сколько их стало.
Организм, напротив, — это единичный объект, и в его геноме могут присутствовать, говоря упрощенно, только одна или две копии интересующего нас аллеля.
Иногда эгоистичному гену выгодно пожертвовать одной-двумя своими копиями для того, чтобы обеспечить преимущество остальным своим копиям, которые заключены в других организмах.
К этой мысли биологи стали подходить уже в 30-е годы прошлого века. Важный вклад в понимание эволюции альтруизма внесли Рональд Фишер (Гениальный биолог-теоретик, один из создателей генетической теории эволюции и современной статистики. У него были свои недостатки: рассказывают, что он заставлял жену рожать ребенка за ребенком, потому что считал, что у него шикарные гены, которые необходимо растиражировать. Фишер был поклонником евгеники), Джон Холдейн (Тоже великий биолог. Прославился своими афоризмами. Однажды в ответ на заявление некой дамы, которая не могла поверить, что "из одноклеточного организма даже за миллиарды лет может развиться такое сложнейшее существо, как человек", Холдейн сказал: "Мадам, вы сами это проделали. И у вас ушло на это всего девять месяцев". В другой раз на вопрос одного богослова, какая черта личности Создателя наиболее ярко проявилась в его Творении, Холдейн ответил: "Необычайная любовь к жукам" (жуки — самая разнообразная группа живых существ, их известно более 300 000 видов)) и Уильям Гамильтон (Тоже гений. Возможно, величайший биолог-теоретик XX века. Разработал и математически обосновал теорию родственного отбора, эволюционную теорию старения и много других замечательных идей).
Теория, которую они построили, называется теорией родственного отбора. Суть ее образно выразил Холдейн, который однажды сказал: "Я бы отдал жизнь за двух братьев или восьмерых кузенов". Что он имел при этом в виду, можно понять из формулы, которая вошла в науку под названием "правило Гамильтона".
Вот эта формула. "Ген альтруизма" (точнее, аллель, способствующий альтруистическому поведению) будет поддержан отбором и распространится в популяции, если
RB > С,
где R — степень генетического родства жертвователя и "принимающего жертву" (на самом деле родство важно не само по себе, а только как фактор, определяющий вероятность того, что у "принимающего" имеется тот же самый аллель альтруизма, что и у жертвователя); В — репродуктивное преимущество, полученное адресатом альтруистического акта; С — репродуктивный ущерб, нанесенный "жертвователем" самому себе. Репродуктивный выигрыш или ущерб можно измерять, например, числом оставленных или не оставленных потомков.
С учетом того, что от акта альтруизма может выиграть не одна, а много особей, формулу можно модифицировать следующим образом:
NRB > С,
где N — число принимающих жертву.
Обратите внимание, что правило Гамильтона не вводит никаких дополнительных сущностей, не требует специальных допущений и даже не нуждается в экспериментальной проверке. Оно чисто логически выводится из определений величин R, В, С и N — точно так же, как геометрические теоремы выводятся из аксиом. Если NRB > С, "аллель альтруизма" будет совершенно автоматически увеличивать свою частоту в генофонде популяции.
Посмотрим, как это работает, что называется, на пальцах. Допустим, наш аллель альтруизма заставляет своего носителя пожертвовать жизнью, если эта жертва спасает жизнь трех его родных братьев (с двумя Холдейн погорячился: два брата — это шило на мыло; почему, будет объяснено ниже). Допустим, это действие совершается в молодом возрасте, когда никто из участников еще не имеет детей, и допустим, что в среднем каждый самец в этой популяции оставляет троих детей.
Подсчитаем значения переменных.
Число адресатов (N) равно трем (три брата).
Родство (R) равно 0,5. Жертвователь получил свой аллель альтруизма от одного из родителей. С каждым из братьев он имеет в среднем 50 % общих генов. В данном случае "общих" означает "идентичных по происхождению", то есть представляющих собой точные копии одного и того же родительского гена. Поэтому вероятность того, что данный брат имеет копию того же самого аллеля альтруизма, равна в среднем 0,5.
Репродуктивный выигрыш (В) равен трем. Брат оставит троих потомков, если выживет, и ни одного, если погибнет.
Репродуктивный ущерб (С) тоже равен трем. Совершив альтруистический акт, жертвователь погибает и не оставляет троих детей, которых он оставил бы, не совершив самопожертвования.
Подставив эти числа в неравенство, получаем выражение: NRB = 4,5 > 3. Неравенство истинно, значит, аллель альтруизма при данных условиях должен распространяться.
Проверим, так ли это.
Если жертва будет принесена, спасенные братья оставят по три потомка. Каждый брат имеет аллель альтруизма с вероятностью 0,5. Каждому из их детей этот аллель, если он у них есть, достанется тоже с вероятностью 0,5. Всего, таким образом, получится девять потомков, каждый из которых имеет аллель альтруизма с вероятностью 0,25. В среднем в следующее поколение перейдет 9 х 0,25 = 2,25 копии аллеля альтруизма.
Если жертва не будет принесена, в следующее поколение перейдет в среднем С х 0,5 = 1,5 копий аллеля. Мораль: аллелю выгодно, чтобы жертвователь совершил альтруистический акт. Благодаря этому акту аллель передаст в следующее поколение не 1,5, а 2,25 своей копии. Следовательно, аллель альтруизма с течением времени будет наращивать свою частоту, вытесняя из генофонда конкурирующий "аллель эгоизма". Подставив двух братьев вместо трех, получим равенство: 1,5 = 1,5.
С точки зрения самого аллеля никакого альтруизма тут нет, один сплошной эгоизм. Аллель заставляет своих носителей — то есть организмы — жертвовать собой, но тем самым аллель блюдет свои корыстные интересы. Он жертвует небольшим числом своих копий, чтобы дать преимущество большему числу точно таких же своих копий. Естественный отбор — это автоматическое взвешивание суммы выигрышей и проигрышей для аллеля — для всех его копий вместе, и если выигрыши перевешивают, аллель распространяется. Вот, собственно и вся теория.
Глупая чайка
Правило Гамильтона обладает замечательной объясняющей и предсказательной силой. Например, оно помогает объяснить типичную сцену, которую можно наблюдать на берегах водоемов (я ее часто наблюдаю летом на Белом море). Поймал рыбак рыбку и начинает ее на берегу потрошить, бросая потроха в воду. Это замечает чайка, она прилетает и начинает хватать потроха из воды. Но она делает это не молча, а сначала издает несколько громких призывных криков. На эти крики быстро слетается еще десятка два чаек, которые тут же набрасываются на первую чайку и начинают отнимать у нее добычу. Та не отдает, отбивается, разыгрывается целый спектакль с вырыванием друг у друга из клюва рыбьих потрохов. Странное поведение! С одной стороны, почему бы чайке не есть молча? Зачем она позвала других, создав тем самым себе проблемы? Второй вопрос: если уж она их позвала, то почему тогда не хочет поделиться, а дерется и не отдает?
В этой сцене, как и во многих других ситуациях в живой природе, мы видим причудливое сочетание альтруистического и эгоистического поведения. Призывный пищевой крик чайки — типичный пример альтруизма. От этого крика чайка не получает никакой выгоды. Выигрыш достается другим чайкам: они получают шанс пообедать. Вторая часть сцены — драка. Здесь уже, конечно, мы видим лишь чистый эгоизм со стороны всех участников.
Разгадка — в правиле Гамильтона. Чайки на Белом море питаются в основном стайными рыбами, например селедкой. Если чайка заметила одну рыбку, то, скорее всего, рядом есть много других: на всех хватит. Это значит, что величина С — цена альтруистического акта — будет в среднем низкой. Величина В — выигрыш тех, кто прилетит на крик, — будет довольно большой: они пообедают. Поскольку рыба стайная, следующую стаю, возможно, придется долго ждать. Величина R (родство) тоже, скорее всего, будет высокой, потому что чайки гнездятся колониями, часто возвращаются на одно и то же место после зимовки, и поэтому, скорее всего, рядом с этой чайкой гнездятся ее родственники — родители, дети, братья и племянники.
Величина N — число чаек, которые услышат, прилетят и пообедают, тоже довольно высоко. Неравенство NRB > С выполняется. Вот и закрепились в популяции чаек мутации, способствующие "пищевому крику". А почему чайка не делится своей добычей, не отдает то, что уже схватила? Потому что в этом случае величина С больше: чайка остается без обеда. Величина N, напротив, оказывается меньше. Отдав свою добычу другой чайке, она накормит одну, а не целую стаю. Неравенство не выполняется, и мутации, склоняющие чайку поделиться добычей, не закрепляются, отсеиваются отбором.
Конечно, выгоднее всего для чайки (точнее, для ее генов) было бы научиться различать ситуацию, когда пищи много и хватит на всех, и когда пищи мало. В первом случае выгодно кричать, а во втором помалкивать. Но для таких калькуляций нужны мозги. А мозг, как мы знаем, дорогой орган. Отбор, как правило, пытается сэкономить на мозгах. К тому же мозги тяжелые. Чайкам надо летать, а не решать алгебраические задачи. Поэтому птица и не может сообразить, когда ей выгодно звать товарок, а когда нет, и ее поведение оказывается нелогичным. Не всегда, а только при недостатке рыбок.
Эволюция альтруизма особенно далеко зашла у перепончатокрылых насекомых: муравьев, пчел, ос, шмелей. У общественных перепончатокрылых большинство самок отказываются от собственного размножения, чтобы выкармливать сестер. Это высшее проявление альтруизма. Таких животных называют эусоциальными, то есть "истинно общественными". Но почему именно перепончатокрылые?
Гамильтон предположил, что дело тут в особенностях наследования пола. У перепончатокрылых самки имеют двойной набор хромосом, а самцы — одинарный. Из-за этого складывается парадоксальная ситуация: сестры оказываются более близкими родственницами, чем мать и дочь. У большинства животных сестры имеют 50 % общих (идентичных по происхождению) генов. Величина R в формуле Гамильтона равна 1/2. У перепончатокрылых сестры имеют 75 % общих генов (R = 3/4), потому что каждая сестра получает от отца не половину его хромосом, а весь геном полностью. Мать и дочь у перепончатокрылых имеют, как и у других животных, лишь 50 % общих генов. Вот и получается, что самкам перепончатокрылых при прочих равных выгоднее выращивать сестер, чем дочерей.
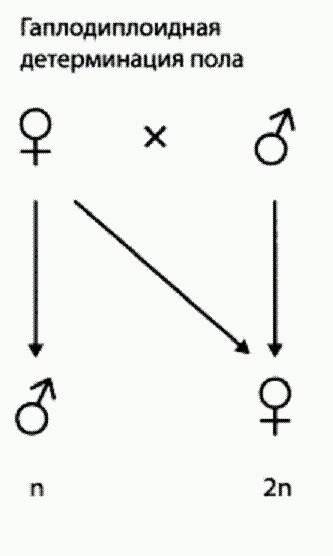
Механизм наследования пола у перепончатокрылых. Самка диплоидная, то есть имеет двойной набор хромосом (2n). Она может отложить неоплодотворенное яйцо с одинарным набором хромосом (п), из которого выведется гаплоидный самец. Если же яйцо будет оплодотворено, то его хромосомный набор будет двойным, и из него выведется самка. Самка получает половину хромосом от матери, половину — от отца. Самец получает от матери половину ее хромосом, а отца у него нет. Такой механизм наследования пола называется гаплодиплоидным.
В действительности все несколько сложнее. Кроме сестер есть еще и братья-трутни, которые имеют со своими сестрами лишь 25 % общих генов (если смотреть со стороны сестры) или 50 % (с точки зрения брата). Однако рабочие самки выращивают и братьев тоже (хоть и недолюбливают их). Мы не будем вдаваться в эту довольно сложную теоретическую область, тем более что приматы, которые нас интересуют, не являются гаплодиплоидами. Но у общественных перепончатокрылых есть (или было в эволюционном прошлом) еще одно важное свойство, резко повышающее вероятность развития альтруизма под действием родственного отбора. Это свойство — моногамия.
Потомки моногамных диплоидных родителей имеют в среднем по 50 % общих генов (R = 0,5). У потомков самки, спаривающейся со многими самцами, средняя величина R стремится к 0,25 (если самцов достаточно много). Для родственного отбора это очень серьезная разница. При R = 0,5 достаточно любого пустяка, чтобы склонить чашу весов в сторону предпочтения сестер и братьев. При R = 0,25 свои дети однозначно дороже. Очень важно, что моногамия свойственна термитам — второму отряду насекомых, в котором эусоциальность получила широкое распространение, причем без всякой гаплодиплоидности. У термитов работают не только самки, но и самцы (они диплоидные, как и их сестры).
Как мы помним, моногамия, вероятно, была свойственна древним гоминидам. Это могло стать мощным стимулом для развития под действием родственного отбора братской (и сестринской) взаимовыручки, внутрисемейной кооперации и альтруизма. И еще, конечно, отцовской любви, а заодно и преданности детей обоим родителям, а не только матери. Возможно, всю эту гамму альтруистических чувств родственный отбор смог поддержать у наших предков именно потому, что они были — хотя бы отчасти — моногамными.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Естественный отбор
Естественный отбор Итак, Дарвин показал, что основным средством создания всех пород домашних животных является искусственный отбор. Даже в то далёкое время, когда люди вели этот отбор, не ставя перед собой определённой цели, бессознательно, они достигали поразительных
Естественный отбор
Естественный отбор Я не усматриваю предела деятельности этой силы, медленно и прекрасно приспособляющей каждую форму к самым сложным жизненным отношениям. Ч. Дарвин Осы, бабочки и дарвинизмВ предыдущих главах мы неоднократно говорили о естественном отборе. Это и
6. ИСКУССТВЕННЫЙ ОТБОР
6. ИСКУССТВЕННЫЙ ОТБОР В эволюции животных действовали вполне понятные законы, построенные на изменчивости, отборе и наследуемости полезных признаков. Эти незатейливые принципы распространялись на предков человека вплоть до райской эпохи. Райский период эволюции
2. Родственный отбор
2. Родственный отбор «Я бы отдал жизнь за двух братьев или восьмерых кузенов» Дж. Холдейн Создатели теории родственного отбора: Рональд Фишер (1890–1962), Джон Холдейн (1892–1964), Уильям Гамильтон (1936–2000).К этой мысли биологи стали подходить уже в 30-е годы прошлого века.
Бессознательный отбор.
Бессознательный отбор. В настоящее время выдающиеся животноводы пытаются путем методического отбора, преследующего определенную цель, произвести новую расу или подпороду, превосходящую все прочие в стране. Но для нашей цели важнее та форма отбора, которую можно назвать
Половой отбор.
Половой отбор. Так как при доместикации часто возникают у одного из полов особенности, наследственно связанные с этим полом, то, без сомнения, то же должно встречаться и в природе. Таким образом, возникает возможность модификации каждого из обоих полов путем
Половой отбор
Половой отбор Для того чтобы мы окончательно не деградировали и не вымерли, как мамонты, Ее Величество Природа придумала половой отбор, который невозможен при партеногенезе и гермафродитизме. Половой отбор заключается в выборе полового партнера по каким-либо внешним
ЕСТЕСТВЕННЫЙ ОТБОР
ЕСТЕСТВЕННЫЙ ОТБОР Естественным отбором считается процесс, который Дарвин назвал «борьба за существование», при котором наиболее приспособленные организмы выживают, а наименее приспособленные погибают. Согласно положениям дарвинизма, естественный отбор в популяции с
ИСКУССТВЕННЫЙ ОТБОР ЖИВОТНЫХ
ИСКУССТВЕННЫЙ ОТБОР ЖИВОТНЫХ На протяжении многих тысяч лет люди высоко ценили породистых животных. Такие животные получились в результате искусственного отбора. Главная цель искусственного отбора — вывести животных с определенными желаемыми признаками; скотоводы,
ИСКУССТВЕННЫЙ ОТБОР РАСТЕНИЙ
ИСКУССТВЕННЫЙ ОТБОР РАСТЕНИЙ Одним из пионеров селекции (искусственного отбора) растений был американец Лютер Бербанк. Он вывел многочисленные новые сорта растений. Пожалуй, самым известным его вкладом в селекцию растений является улучшение сортов картофеля. Однажды в
ОТБОР
ОТБОР Естественный отбор — это процесс, сокращающий в популяции пропорцию организмов, относительно неприспособленных к выживанию и увеличивающих пропорцию организмов с фенотипом, близким к оптимальному, для определенной среды и уровня конкуренции. Здесь важен высокий
ГДЕ ВЫБОР, ТАМ И ПОЛОВОЙ ОТБОР
ГДЕ ВЫБОР, ТАМ И ПОЛОВОЙ ОТБОР Биологическая цель соревнования и выбора состоит в том, чтобы в первую очередь обеспечить воспроизведение наиболее полноценных особей и воспрепятствовать размножению неполноценных. В инстинктивных программах самки заложено стремление
Половой отбор
Половой отбор Половой отбор представляет собой важную составляющую естественного отбора. Ч. Дарвин определял его как «форму естественного отбора, направленную на закрепление преимуществ особей, проявленных в период размножения». Ч. Дарвин сформулировал два основных
Естественный отбор
Естественный отбор Естественный отбор – важнейший фактор эволюции. Дарвинизм (а именно на базе дарвинизма построена СТЭ), как уже отмечалось выше, называют теорией естественного отбора.Кратким и удачным определением отбора может служить сформулированное И. Лернером:
Половой отбор
Половой отбор Большой интерес представляет выделенный еще Ч. Дарвином половой отбор, который сыграл заметную роль в эволюции человека. Дарвин различал два вида полового отбора, и его наблюдения подтвердились в дальнейших исследованиях. Первый вид – это состязание
Групповой отбор
Групповой отбор Подходя в основном нормально к эволюционной психологии, Дарвин уступил искушению, известному как групповой отбор. Рассмотрим его центральное объяснение эволюции морального чувства. В "Происхождении человека" он написал, что "повышение стандартов этики и