Естественный отбор
Естественный отбор
Я не усматриваю предела деятельности этой силы, медленно и прекрасно приспособляющей каждую форму к самым сложным жизненным отношениям.
Ч. Дарвин

Осы, бабочки и дарвинизм
В предыдущих главах мы неоднократно говорили о естественном отборе. Это и понятно: вся сущность теории Дарвина основана на широчайшем распространении в природе селективных процессов. Однако следует эту проблему рассмотреть подробнее — слишком уж она важна. Остановимся в первую очередь на доказательствах существования отбора в природе и его творческой роли. Как я уже имел случай упомянуть, большинство современных критиков дарвинизма отказывают отбору в творческой роли, считая его в лучшем случае могильщиком, уничтожающим ненужное. Так, современный антидарвинист А. Вандель пишет: «Во всех городах имеется служба канализации, отбор играет аналогичную роль в органической эволюции. Его специфическая функция заключается в устранении чудовищ, неудачников, немощных стариков».
Другие авторы (например, известный палеонтолог О. Шиндевольф) в принципе считают, что отбор может быть творческим началом в микроэволюции — возникновении новых форм, подвидов и видов. Однако макроэволюция — становление крупных систематических единиц — типов, классов, отрядов, семейств — управляется иными законами (о которых, правда, говорят сбивчиво и смутно).
Но, как справедливо писал Кольцов: «…для нас, верящих в неизменность закона постоянства энергии, термин „творить“ может иметь только одно значение: из многих комбинаций выбирать только одну. Поэтому я считаю, что мы и теперь, как 50 лет назад, имеем право спокойно утверждать: „естественный отбор творит новые формы“».
Рассмотрим несколько примеров селективных процессов в природе.
Быть может, одно из важнейших доказательств теории естественного отбора — существование приспособительных, покровительственных окрасок и форм у животных и растений. Как уже упоминалось раньше, ламаркизм полностью бессилен истолковать, например, белую окраску белого медведя и зайца-беляка, отпугивающие птиц окраски многих насекомых, подражание безвредных животных хорошо защищенным и многое другое. Выход, однако, был найден: если факты нельзя истолковать, их надлежит отрицать. Известный этолог Н. Тинберген так писал о подобных ниспровергателях: «Нельзя без удивления читать некоторые из этих псевдокритических писаний, и особенно содержащиеся в них язвительные насмешки по адресу „кабинетной позиции“ тех, кто верит в покровительственную окраску. Удивление испытываешь потому, что именно эта критика исходила из кабинетов, а те, кто верил — были естествоиспытателями».
Образцом «псевдокритического писания» является, в частности, книга Ф. Гейкертингера, в которой сконцентрированы все доводы антидарвинистов против дарвиновского истолкования этой проблемы. Почтенный австрийский энтомолог одним махом «закрывает» такие явления в природе, как криптизм (имитация фона местообитания), мимезия (имитация формы субстрата) и мимикрия (имитация защищенного животного). Одновременно достается и предостерегающей окраске, а так как он не заинтересован в объективном изложении фактов, победа остается на его стороне.
Стоит подробнее остановиться на феномене так называемого «индустриального меланизма» (меланос — по-гречески черный), ибо история его изучения — блестящий пример торжества дарвиновского объяснения криптизма.
По всей Европе и Азии, от Англии до Японии, обитает бабочка — березовая пяденица. Обычная ее форма белого цвета с мелкими черными пятнышками на крыльях и теле. Такие бабочки очень трудно различимы на своем субстрате — березовых стволах, покрытых лишайниками. Но уже в 1850 году в окрестностях английского города Манчестера была поймана березовая пяденица необычайной расцветки — черная с редкими белыми пятнышками, как бы негатив типичной. Само по себе явление меланизма не такое уж редкое. Ученых удивило другое: с каждым годом темные бабочки стали попадаться все чаще и чаще. Черная форма (карбонария) явно вытесняла типичную из прежних мест обитания. Впоследствии была описана и другая форма (островная, или инсулярия), более темная, чем типичная, но светлее карбонарии. Удалось установить, что окраска карбонарии обуславливается одним доминантным геном.
Четвериков знал об этом явлении и пророчески писал, что темная форма «каким-то, нам точно неизвестным образом, является лучше приспособленной к условиям своего существования, чем типичная». Далее он прогнозировал дальнейшее вытеснение основной формы вплоть до самых восточных границ ареала, до тех пор, пока белая форма не останется лишь на изолированных Японских островах как островной подвид. Такие случаи уже наблюдались: у сумеречной бабочки — хмелевого тонкопряда имеется четко выраженный половой диморфизм (самец со снежно-белыми крыльями), однако на Шотландских островах сохранилась примитивная форма, у которой самец и самка по окраске не различаются.
Теперь, когда мы знаем, к каким условиям лучше приспособлены темные формы березовой пяденицы, нам остается только желать, чтобы прогноз Четверикова не оправдался; более того, всеми силами противодействовать его осуществлению. Оказалось, что темные формы на своей родине — Британских островах — распределены неравномерно. Почти 100 % инсулярий и карбонарий приурочены к индустриальным районам, где стволы берез покрыты сажей и каменноугольной копотью, а лишайники погибли. Темных пядениц нет или еще мало в сельских районах Северной Шотландии, Корнуэлле, Уэльсе и Ирландии. Та же картина наблюдается и в континентальной Европе. Глядя на карту распространения бабочек-меланистов, мы можем судить о распределении предприятий тяжелой промышленности!
Ген меланизма делает бабочку на стадии гусеницы более устойчивой ко всякого рода индустриальным ядам, но это не главная причина победного шествия карбонарий и инсулярий. Английский исследователь Б. Кетлуэлл подметил, что в «закопченном» лесу белые формы видны с далекого расстояния и должны уничтожаться птицами. Наоборот, в местностях, не подверженных влиянию индустриализации, меланисты более заметны, чем типичные бабочки.
Выводы Кетлуэлла оспаривались теми, кто считал, что птицы не охотятся за сидящими бабочками. Поэтому Кетлуэлл обратился за помощью к одному из основателей этологии — Тинбергену. Прямые наблюдения, документированные киносъемкой, с непреложной убедительностью показали, что в чистом лесу птицами (мухоловками, овсянками, поползнями и горихвостками) уничтожаются в первую очередь темные формы, а в загрязненном — светлые.
В тех случаях, когда район обитания пяденицы только начинает подвергаться индустриальному загрязнению, типичная форма вытесняется сначала инсулярией и лишь потом — более темной карбонарий. Доминантность генов меланизма по мере того, как человек загрязнял вокруг себя среду, увеличивалась. Сто лет назад пяденицы-меланисты, гомозиготные по этому признаку, имели больше белых отметин, чем гетерозиготы сегодня.
Так было расшифровано явление «индустриального меланизма». Хотя непредубежденные исследователи нашли уже многие десятки примеров, доказывающих приспособительность криптических окрасок, случай с березовой пяденицей, когда человек сам стал свидетелем эволюции и, в некотором смысле, ее виновником, наверное, самый убедительный.
«Индустриальный меланизм» обнаружен и у других насекомых. Любопытно, что там, где заводы и фабрики переводятся с каменного угля и мазута на электроэнергию и газ, он исчезает. В окрестностях Бирмингема численность темной формы двухточечной божьей коровки вслед за очищением воздуха сократилась вдвое. Причина этого ясна: с исчезновением загрязненности черные формы — меланисты становятся лучше видны врагам, в первую очередь — птицам.
Изящный и достоверный опыт, доказывающий селективное значение покровительственной, защитной окраски — криптизма — проделал Ф. Самнер. Всем известный, вошедший в пословицы хамелеон — отнюдь не единственное животное, способное менять окраску в зависимости от фона обитания. Многие рыбы, например, наделены этой способностью, хотя и меняются не так быстро. Рыболовы, наверное, знают, что в реках со светлым, песчаным дном рыбы светлее, чем особи того же вида в реках с дном темным, илистым.
Самнер выдерживал мелких пресноводных рыбок — гамбузий в аквариумах с черным и белым дном и через несколько часов, когда они изменяли окраску сообразно с фоном, пересаживал светлых и темных поровну в бассейны (также светлые или темные), куда выпускал хищников. В качестве хищников выступали хищные рыбы, пингвины и цапли. Селективное значение окраски проявилось в опытах Самнера очень четко: хищники уничтожали в первую очередь рыб, цвет которых контрастировал с фоном обитания.
Эффективность защитной окраски продемонстрирована также в опытах А. Неснолы, который привязывал зеленых и бурых богомолов на разные растения. Зеленые богомолы на бурых растениях быстро уничтожались птицами, а те особи, окраска которых соответствовала фону, оставались невредимыми. Опыт Чеснолы проверил в усовершенствованном варианте М. М. Беляев. Оказалось, что защитная окраска, как и всякое приспособление, не абсолютна: одни птицы не замечают криптически окрашенных насекомых, а другие, например вороны, склевывают их на любом фоне.
Подчеркнем еще одно обстоятельство. Криптическая окраска приносила бы не много пользы, если бы не дополнялась специфическими чертами поведения ее обладателей. Белый медведь, подползая к тюленю, старается прикрыть лапой глаза и черный нос — единственные демаскирующие детали строения. Разноцветные ящерицы анолисы, будучи потревоженными, разбегаются и затаиваются на фоне разного цвета: серые на сером, зеленые на зеленом. Зеленая форма богомола, по наблюдениям Беляева, предпочитает зеленые же, не выгоревшие растения. Таких примеров можно привести гораздо больше. Самое любопытное, что черная и светлая формы березовой пяденицы также выбирают фон. Кетлуэлл проделал такой опыт: он выкрасил внутреннюю поверхность бочонка черными и белыми полосами, а затем выпускал туда бабочек разного цвета — около 70 % бабочек предпочитало садиться именно на те полосы, которые соответствовали их же цвету.
По всей Европе широко распространена наземная улитка цепея (близкая родственница виноградной улитки). Уже давно было подмечено, что колонии улиток этого вида из разных мест отличаются по цвету раковины. Иногда в популяции преобладают особи, имеющие светло-желтую или розоватую раковину, иногда особи с бурой раковиной. Раковина может быть полосатой или же не иметь полос. Высказывались предположения, что мы имеем здесь дело с проявлением дрейфа генов. Действительно, малочисленные популяции улиток с крайне ограниченной способностью к передвижению — как будто бы благодатный материал для обнаружения генетико-автоматических процессов.
Однако дело обстоит гораздо сложнее. Английские исследователи Кэйн и Шеппард показали, что популяции с преобладанием полосатых и светлоокрашенных особей обитают в кустарниках и траве. На однотонном темном фоне (почва в темных буковых лесах) доминируют бурые и неполосатые улитки. Окраска раковин оказалась криптической. Главный фактор отбора здесь, по-видимому, птицы. Это удалось доказать, исследуя «столовые» певчих дроздов. Дрозды разбивают раковины улиток о камни, поэтому у скоплений камней всегда находится много обломков раковин съеденных моллюсков; в них преобладали останки особей, контрастировавших с фоном обитания.
Примечательно, что отбор в данном случае не доводит дело до конца, в противном случае в траве обитали бы только светлые и полосатые, а в лесу только однотонные и бурые улитки. Дело в том, что фон обитания разный в разные сезоны года. Ранней весной, когда трава еще не выросла, улитки с однотонными раковинами имеют селективное преимущество даже на лугу. Кроме того, хищники не единственный фактор отбора. Полиморфизм в популяциях улиток поддерживается совместным селективным действием биотических факторов (птицы) и абиотических (главным образом температура и влажность). Есть опыты, показывающие, что формы с однотонной и светлой раковиной более устойчивы к колебаниям температур.

Предостерегающая окраска: вверху — бабочка-пестрянка в полете и сидящая (содержит в крови синильную кислоту); внизу — клопы, несъедобные из-за дурного запаха и вкуса.
Говоря об опытах Чеснолы и Беляева, мы уже упоминали, что популяции богомола полиморфны: в них встречаются как ярко-зеленые, так и бурые формы. В засушливых местностях, где растения летом выгорают, преобладают бурые богомолы, а там, где трава остается зеленой — зеленые. В окрестностях Карадага, где Беляев ставил сбои опыты, подходящим по фону субстратом для зеленой формы к концу лета оставались фактически одни ползучие плети диких каперцев. Есть четкая корреляция между уровнем осадков в данной местности и процентом бурых форм: с приближением к среднеазиатским пустыням зеленая форма практически выпадает. Вспомните уже рассмотренный нами пример из генетики человека — возрастание процента группы крови В по мере приближения к древним очагам оспы!
Итак, природа давным-давно использует такие выдумки, человека, как защитную форму-хаки, белый халат лыжника и пятнистый комбинезон десантника. Подчеркнем, что возникнуть защитные окраски могли только в результате отбора, проводимого хищниками. К окраске может прибавляться и форма — известны насекомые, подражающие по форме сухим сучкам, листьям, птичьему помету и иным малосъедобным предметам (этот феномен называется мимезией).
Но так бывает отнюдь не всегда. Многие животные не только не прячутся, а, наоборот, громко заявляют о своем существовании. Окраска у них яркая, «кричащая», они не делают никаких попыток скрываться. Причина ясна: они несъедобны из-за противного вкуса или запаха, или же ядовиты, вооружены колючками или жалом. Такая окраска называется предостерегающей или апосематической.

В начале опыта стрекоза раскачивается на ниточке перед глазами голодной жабы. (Жабы ловят только движущихся насекомых.)

Жаба жадно хватает приманку; изо рта торчат только брюшко и крылья жертвы.

Затем жабе предлагается муха-ктырь, обладающая большим внешним сходством со шмелем, но не способная жалить.

Жаба, которая никогда не имела дела со шмелем, пожирает муху так же охотно, как стрекозу.
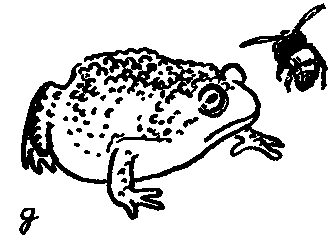
Теперь перед жабой раскачивается шмель. Как и предыдущие приманки, насекомое не способно улететь, но может жалить.
Так и случилось! Жаба кидается на приманку, но получает яростный укол в язык и выплевывает шмеля.

Когда жабе преподносят следующего шмеля, она не бросается на него, а пригибает голову к земле, хорошо помня полученный урок.

Теперь земноводному предлагают муху-ктыря. Жаба отказывается и от этой приманки, принимая муху за шмеля: предостерегающая окраска мухи действует вполне эффективно.

Чтобы доказать, что жаба отвергает эти приманки не потому, что она стала всего остерегаться или просто насытилась, животному предлагают стрекозу.

Жаба хватает стрекозу так же охотно, как и в начале опыта. Итак, она еще голодна, но научилась отличать съедобную пищу от несъедобной.
Большой популярностью у малосъедобных животных пользуется комбинация черного и желтого цветов. Черно-желтая «тельняшка» осы хорошо известна всем. Не менее бросаются в глаза черно-желтые гусеницы бабочек-медведиц, из-за отвратительного вкуса избегаемые птицами. Эта же окраска встречается и У позвоночных животных. Окраска одного из немногих сомов, перешедших к жизни в море — плотозуса, так же как и наших амурских сомиков-касаток, представляет комбинацию черных пятен и полос на желтом фоне (эти рыбы вооружены ядовитыми колючками, уколы которых вызывают сильную боль). Ядовитая слизь хорошо защищает черно-желтую пятнистую саламандру. Но еще лучше защищена от врагов пятнистая древесница — самая ядовитая лягушка земного шара, кожными выделениями которой индейцы Колумбии отравляли наконечники стрел. Яд древесницы влияет на центральную нервную систему и так силен, что стрела с этим ядом действенна не только против крупных обезьян, но и ягуаров. Расцветка древесницы — белые, желтые или оранжевые пятна по темно-каштановому или черному фону. Вспомним, наконец, единственных ядовитых ящериц — ядо-зубов. Они тоже расписаны черными полосами и пятнами по ярко-желтому или оранжевому фону.
Здесь так же, как и в случае криптической окраски, природа предвосхитила многие человеческие выдумки. Везде, где требуется обратить на что-то внимание, применяются комбинации белого, черного, красного и желтого цветов. Полосатый черно-белый шлагбаум, красно-желтый трилистник знака радиации, дорожные знаки и кричащие цвета реклам — все это уже не ново под луной.
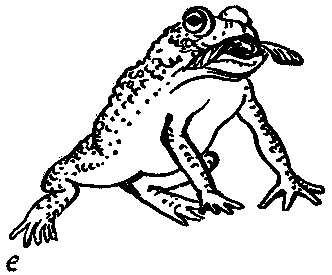
Гейкертингер и другие антидарвинисты пытались показать, что апосематическая окраска действенна лишь потому, что птицы и другие хищники избегают ярких и непривычных сочетаний цветов. Что это не так, показывают прямые опыты: молодые птицы, ящерицы и земноводные, никогда прежде не сталкивавшиеся с апосематическими животными, быстро, в прямом смысле слова «на горьком опыте» обучаются избегать их. Для хамелеона, например, одной попытки поймать пчелу более чем достаточно. Тинберген в одной из своих книг, переведенной на русский язык, приводит забавный рассказ в картинках-фотографиях: жаба с аппетитом съедает стрекозу, потом пчеловидную муху, затем ей предлагают шмеля. Убедившись в несъедобности последнего, она избегает и муху-пчеловидку, но по-прежнему ест стрекоз. Все наукообразные измышления противников теории естественного отбора развенчиваются этим комиксом!
Наказанный за жадность хищник хорошо помнит урок (лягушки и жабы не менее 10 дней), причем действенность его зависит от защищенности жертвы. Любопытно еще одно свойство апосематических животных — почти все они необычайно живучи и им не так-то просто нанести смертельное ранение. Бабочку-данаиду, выделяющую зловонную жидкость из сочленений ног и усиков, трудно раздавить пальцами, а живучесть хорька, контрастно окрашенного и выделяющего при опасности отвратительно пахнущий секрет пахучих желез, вошла в пословицу. Говорят, что американские скунсы еще более живучи. Такая стойкость к повреждениям вполне понятна — животному нужно отразить первую атаку хищника, а на вторую тот уже не осмелится.
Реакция «избегания» возникает тем быстрее, чем лучше защищена жертва. Были поставлены любопытные опыты, где «хищниками» были цыплята, а «жертвой» — подкрашенная вода с добавкой веществ горького вкуса. Оказалось, что цыплята отлично обучаются отличать «защищенную жертву», причем чем более горькая была вода, тем более тонкие оттенки цвета они различали.

Иногда криптическая окраска сочетается с ложной предостерегающей (демонстративной). Вверху — глазчатый бражник в спокойном состоянии. Если его потревожить, бражник открывает глазчатые пятна на второй паре крыльев.
По-иному защищает так называемая ложная предостерегающая или демонстративная окраска. Организмы, окрашенные так, обычно вполне съедобны. Хороший пример демонстративной окраски — яркие глазчатые пятна на крыльях многих бабочек из разных семейств. В наших широтах наиболее распространены окрашенные так дневной павлиний глаз — родственник крапивницы и сумеречная бабочка глазчатый бражник. Как правило, они сидят, если их не тревожить, так, чтобы глазчатые пятна были не видны. Достаточно их побеспокоить, как они раскрывают крылья, и открываются пятна, похожие на глаза крупного позвоночного. Это чистый блеф, однако, по наблюдениям Д. Блеста, птицы пугаются их и немедленно бросают вполне безобидную добычу. Но стоит стереть «глаза» с крыльев, как бабочки становятся беззащитными.
Да, но не имеем ли мы здесь дело с примером параллелизма изменчивости? Исследования показали, что это не так. Механизм образования глазчатых пятен разный у разных видов. Появление их контролируется разными генами. Это не параллелизм изменчивости, а приспособление, возникающее независимо, конвергентно. Подобным путем возникло сходство между акулой, дельфином и подводной лодкой…
Высшая ступень приспособительной окраски — мимикрия, когда организм «подражает», причем не только в окраске, но нередко и в форме, и в поведении организму другого вида, хорошо защищенному. Обычный пример — пчелы, осы и шмели, защищенные ядовитым жалом и противным вкусом внутренностей, с предупреждающей черно-желтой полосатой окраской и подражающие им в окраске безобидные мухи. Но история открытия мимикрии сложнее и насчитывает уже более века.
В 60-х годах прошлого столетия практически одновременно Г. Бэйтс (автор прекрасной книги «Натуралист на Амазонке»), его друг Уоллес, разделивший с Дарвином славу творца теории естественного отбора, и Р. Траймен, изучая бабочек Бразилии, Юго-Восточной Азии и Африки, обнаружили удивительное сходство в окраске, форме и поведении бабочек из неродственных групп.

Примеры мимикрии. Первый ряд — несъедобная бабочка из семейства геликонид; второй — мимикрирующая ее дизморфина; третий — слева шмель, справа муха журчалка шмелевидная; четвертый — слева оса, справа муха журчалка осовидная.
Причины этого «маскарада» вскоре были разгаданы. Мимикрия Бэйтса — это подражание съедобных бабочек тем видам, которые несъедобны или малосъедобны для птиц из-за ядовитости или неприятного вкуса или запаха. Для многих мимикрирующих бабочек уже расшифрованы генетические механизмы возникновения такой окраски.
Наряду с этим Ф. Мюллер открыл другой вид мимикрии, при которой один несъедобный вид подражает в окраске столь же несъедобному другому. Хищник, попробовавший особь одного вида, избегает затрагивать другой; в результате выигрывают оба. Например, гусеницы бабочки-медведицы покрыты чередующимися черными и желтыми полосами. Для большинства птиц они совершенно несъедобны из-за ядовитых волосков. Молодые птицы, попробовав однажды гусеницу медведицы, отказываются от любого насекомого, покрытого черно-желтыми полосами (например, от осы).
Шире всего мимикрия распространена в тропиках; у нас наиболее известно «подражание» мух пчелам и осам. Некоторые бабочки — бражники и стеклянницы также довольно точно имитируют перепончатокрылых. Мне вспоминается жук-дровосек, который настолько хорошо копировал крупного наездника, что с первого взгляда мне не удалось его узнать.
Есть поразительные примеры сходства совершенно неродственных организмов. В Далмации, например, в одном и том же биотопе — под камнями обитают многоножка и мокрица, покрытые точно такими же яркими пятнами, как и ядовитейший паук каракурт (по-тюркски «черная смерть»), известный жителям Средней Азии.
Шеппард приводит примеры мимикрии у птиц. Так, несъедобному и агрессивному, вроде нашей сороки, дронго часто сопутствует другая птица — черная мухоловка. Но, пожалуй, наиболее любопытный пример — мимикрия яиц кукушки. Европейская кукушка разделена на ряд форм, каждая из которых откладывает яйца в гнезда строго определенного вида птиц. По размерам, форме и цвету яйца каждой формы хорошо соответствуют яйцам хозяев гнезда (в противном случае птица может выбросить чужое яйцо или вообще покинуть гнездо и начать строить новое). Эту мимикрию также можно объяснить только жестким отбором, если, конечно, не считать, что кукушонок вылупляется из яйца пеночки.
Правда, есть нерешенный вопрос: каким образом закрепляется мимикрия в поколениях? Скрещиваются ли формы только внутри себя? На других видах птиц показано, впрочем, что цвет яйца определяется только генотипом самки, это признак, сцепленный с полом. У птиц, однако, нет женской половой хромосомы у (генотип самца хх, самки хо), но известно, что у-хромосома может возникать заново. Здесь орнитологам предстоит еще много работы. Беда в том, что хромосомные наборы птиц — неблагодарный объект для изучения. В них, как правило, много мелких трудноразличимых микрохромосом. Хотя вполне достоверных фактов существования мимикрии более чем достаточно, антидарвинисты неустанно продолжают отвергать ее. Расправляются с мимикрией довольно просто: приводится длинный список птиц, поедающих жалоносных перепончатокрылых, и делается вывод — окраска не приспособительна, ибо не защищает ни имитатора (муху), ни модель (осу). Хорошо, что подобная логика не так уж распространена, а то пришлось бы, следуя ей, отменить в армии защитную форму, потому что она не гарантирует солдату бессмертие.
Дело в том, что никакое приспособление не абсолютно, но если оно обеспечит обладателю хотя бы один лишний шанс в борьбе за существование, оно немедленно будет подхвачено естественным отбором. Защитная форма может спасти солдата от прицельного огня, но бессильна спасти от огня по площадям. Так же и в природе. Черно-желтая «тельняшка» осы не смущает орла-осоеда и золотистую щурку, но отпугивает других птиц. Яркая предупреждающая окраска бабочек-пестрянок, содержащих в гемолимфе сильнейший яд — синильную кислоту, предостерегает птиц, но бессильна против личинок наездников и паразитических мух. Великолепно замаскированные под сухие сучки гусеницы пяденицы незаметны для птиц, но служат объектом охоты для роющих ос.
В последнее время стали известны факты, свидетельствующие о «запаховой» и «звуковой» мимикрии. Цветы многих растений — стаппелии, многих ароидных и других пахнут падалью. На этот запах собирается немало мух; вышедшие из отложенных в цветы яиц личинки погибнут без пищи, но цветы будут опылены их обманутыми родителями. Другие цветы пахнут ванилью — и не случайно. Ванилин и родственные ему вещества — половые аттрактанты многих насекомых, привлекающие самцов к самкам. Изучение запаховой мимикрии долгое время тормозилось несовершенством обоняния человека. Лишь когда была применена новая техника (газовый хроматограф), открытия в этой области хлынули потоком. Мириам Ротшильд, известная специалистка в этой области, полагает, что запаховая мимикрия встречается по крайней мере не реже зрительной. Она же установила любопытный факт: некоторые насекомые «наследуют» ядовитые вещества от растений, которыми питались их личинки. Так, бабочка-данаида — данаус хризиппус (модель для многих имитаторов) на стадии гусеницы питается ядовитыми растениями из семейства ластовниковых.
Мы уже упоминали, что у орхидей рода Офрис к запаховой мимикрии прибавляется зрительная. Их цветы настолько напоминают самок пчел и ос, что самцы пытаются с ними спариваться и опыляют. Самки перепончатокрылых обычно вылетают позже самцов; к тому времени орхидеи уже бывают опылены. Любопытно, что один из таких видов стал в Англии самоопылителем. Жесткий отбор на форму цветка был снят, и в результате накопления неконтролируемых мутаций цветы стали весьма разнообразными по окраске и форме.
Известны и достоверные примеры звуковой мимикрии. Американские исследователи Брауэры слегка подрезали у мух-пчеловидок крылья, так что жужжание в полете изменяло тон. Жабы, прежде остерегавшиеся их, охотно начинали хватать мух, оперированных подобным образом.
Быть может, самый любопытный пример звуковой мимикрии приводит известный путешественник и исследователь Уссурийского края В. К. Арсеньев. Его проводник Дерсу Узала на охоте показал, как тигр подманивает оленя во время гона звуком, имитирующим рев самца («Амба постоянно так изюбра обмани, изюбрь теперь не понимает какой люди кричи»). Подчеркну, что путевые дневники Арсеньева — не «охотничьи рассказы», а безупречный по точности научный документ.
Противники теории естественного отбора усердно подчеркивали все несовершенство приспособительных окрасок и форм. Возможно ли, спрашивали они, чтобы полосатые злаковые мушки, размером от одного до трех миллиметров, успешно имитировали ос, или жуки — нахлебники муравейников — своих хозяев? Упрекая дарвинистов в антропоморфизме, они незаметно впали в него сами. Мимикрия жуков-мирмекофилов в основном запаховая, потому что муравьи «узнают» друг друга по запаху. Смойте с муравья запах, присущий только членам этого муравейника, и братья (а точнее, сестры, ведь рабочие муравьи — бесплодные самки) загрызут его, сочтя чужаком. Пропитайте этим запахом другое насекомое — и оно спокойно начнет жить в муравейнике, даже поедая личинок хозяев, как это делают гусеницы бабочек-голубянок. Оказывается, животные часто опознают другие объекты не по всем деталям структуры, а лишь по каким-либо четким опознавательным признакам — сигнатурам.
Принцип сигнатуры, как назвал его известный биокибернетик Г. Кастлер, широко распространен на всех уровнях живой природы — от молекулярного до организменного. Так, например, при синтезе белка соответствующие транспортные РНК «узнаются» по сигнатурным участкам — антикодонам, хотя бы эти РНК присоединялись не к тем аминокислотам, которые они обычно переносят. Антитела (защитные белки-глобулины) могут «не узнать» белок хозяина, что приводит к аллергии. Самцы бабочки грушевой сатурнии облепляют пропитанную запахом самки тряпку, не обращая внимания на саму самку, накрытую стеклянным колпаком. Самцы маленькой рыбки — колюшки, охраняя свой участок, бросаются на любой красный предмет — поэтому в аквариуме они могут «нападать» на пожарный автомобиль за окном.
Так же рьяно охраняют свой гнездовой участок чайки, накидываясь даже на чучело чужака, помещенное рядом с гнездом. Однако реакцию вызывает только голова: они атакуют отдельно выставленную голову, не реагируя на безголовое чучело. Певчие птицы часто стаями нападают на кукушку — покажите им одну голову кукушки — и реакция будет такой же.
Короче, если сигнатуры модели и имитатора совпадают, по всем другим признакам они могут быть не только несхожими, но и противоположными. Не исключена возможность, что мимикрия распространена в природе шире, чем это кажется человеческому глазу. Не следует дивиться «глупости» природы. Принцип сигнатуры порой доводится до абсурда и в человеческом обществе — вспомним тех бюрократов, которые за бумажкой не видят человека.
Итак, имитатору вполне достаточно походить на модель по тому признаку, по которому модельный объект имеет преимущество в отборе. Например, семена многих сорняков весьма точно мимикрируют семена культурных растений не по внешнему виду, а по аэродинамическим свойствам. Отбор здесь производят веялки, сортирующие семена воздушной струей. А что бы вы сказали о мимикрии, где имитатор — вирус, бактерия или паразитический червь, а модель — человек? Это так называемая молекулярная мимикрия, при которой внешние структурные белки и полисахариды паразитов становятся настолько похожими на белки хозяина, что перестают быть антигенами, то есть не вызывают появления антител. Единственно возможная причина возникновения мимикрии — жесточайший отбор паразитов в теле хозяина, осуществляемый защитными белками — иммуноглобулинами. Все другие попытки объяснения выглядели бы убого.
Мы можем заключить, что теория естественного отбора справляется с объяснением всех перечисленных нами замечательных фактов так же легко, как и сто лет назад, когда Бейтс и Уоллес обнаружили сходство в окраске между неродственными видами бабочек Южной Америки и Юго-Восточной Азии. Фактов, не согласующихся с ней, не найдено.
Идет ли эволюция скачками?
Факты, свидетельствующие о наличии и широком распространении в природе селективных процессов (отбора), настолько многочисленны, что одно их перечисление заняло бы книгу побольше этой. И тем не менее на многих исследователей временами находит как бы затмение, которое сводится к отрицанию творческой роли естественного отбора (при этом, разумеется, все эти факты — а имя им легион — игнорируются). Читая работы этих авторов или беседуя с ними лично, я всеми силами стараюсь понять, почему для них одиозно само понятие «отбор». К сожалению, мои попытки в этом направлении не увенчались успехом. Любопытно, что ни один из доводов, приводимых противниками дарвинизма, не выдерживает сколько-нибудь тщательного логического анализа и, однако, воспроизводится из поколения в поколение (причем каждый раз противники дарвинизма не ведают, что их доводы — лишь повторение давно опровергнутых). Плохое знание литературы нередко приводит к тому, что иной изобретает заново велосипед, причем с треугольными колесами.
Быть может, одним из самых крупных таких «ниспровергателей» является известный немецкий генетик Р. Гольдшмидт. Его идеи можно выразить вкратце так: существуют два процесса — микроэволюция и макроэволюция, возникновение форм выше вида. Микроэволюция происходит внутри вида и управляется факторами, подробно рассмотренными нами ранее (отбор и дрейф генов, ограничение панмиксии и мутационный процесс). Иное дело — виды. Возникновение их идет скачками (сальтационный процесс: корень тот же, что и в слове сальто-мортале), путем образования некоторых мутаций, затрагивающих весь организм (вспомните Де Фриза!). Главная роль здесь отводится крупным хромосомным перестройкам, умножению генов и т. д. и т. д. Основное в этих построениях то, что выше вида эволюция идет не по воззрениям Дарвина и его последователей, начиная с Четверикова.
Нужно признать, что доводы последователей Гольдшмидта не очень убедительны. Для дрозофилы, например, известны мутации, которые изменяют признак, характеризующий отряд, а то и класс. Мутация «четырехкрылая» сразу выводит мутантную особь из отряда двукрылых (вместо жужжалец, характерных для мух и комаров, развивается вторая пара крыльев). Но от этого дрозофила не перестает быть таковой, так же как и человека с чешуйчатой кожей (такие мутации имеются) не надо спешить зачислять в класс рыб. Поэтому идея Гольдшмидта о том, что макроэволюция идет за счет возникновения «многообещающих монстров» (чудовищ), более чем спорна. Чем сильнее мутация перекраивает организм, тем меньше у последнего шансов в борьбе за существование. Эволюция обычно осуществляет принцип, выраженный в японской хокку:
Тихо, тихо ползи,
улитка, по склону Фудзи,
вверх, до самых высот!
Плейотропия гена, как упоминалось выше, не абсолютна. «Системные мутации» Гольдшмидта — это мутации со 100-процентным эффектом, затрагивающим все признаки фенотипа. А вероятность того, что при этом возникнет приспособленный к среде обитания вид, настолько ничтожна, что ее можно и не подсчитывать. Тут вспоминается герой английской сказки, который хотел со шкафа «впрыгнуть» в повешенные на стуле брюки. Разумнее брюки надевать обычным способом; логичнее допустить, что популяция предкового вида переходит в новый вид на протяжении ряда поколений путем формирования нового комплекса аллелей.
Сторонники Гольдшмидта (порой не ведающие, что являются таковыми) иногда приводят на первый взгляд убийственный довод: несмотря на существование внутривидовых форм, виды, по крайней мере за срок человеческого существования, остаются стабильными. Примером может служить хорошо известный всем окунь. Он распространен во всей Евразии и Северной Америке и везде остается окунем (американского иногда выделяют в подвид «желтый», но это явная натяжка. Рыболовы хорошо знают, что и у нас попадаются окуни с ярко-желтым брюхом). Отсюда вывод: вид — стабильная система, состоящая из отдельных, частично, а то и полностью изолированных друг от друга популяций.
Однако система — строго научное понятие, которое не надо употреблять всуе. Нечто подобное получилось с понятием обратной связи — мне приходилось слышать утверждения, что дарвинизм устарел, ибо существует обратная связь между видом и средой. Но ведь в машине обратную связь осуществляет человек; в природе ее может осуществлять или естественный отбор, или святой дух — третьего не дано. То же и с системой; под этим понятием мы подразумеваем совокупность элементов, объединяемых связями. А какие связи могут быть между популяциями? Или обмен генами — или телепатия. Однако обмен генами между американскими и европейскими популяциями невозможен, а в трансконтинентальную телепатию окуней как-то не хочется верить.
Все, что можно сказать по этому поводу, — это то, что есть виды лабильные, легко образующие внутривидовые группировки, порой дивергирующие до видового обособления, и есть виды стабильные, остающиеся в довольно широком диапазоне условий в одной ипостаси. Примером могут быть лососевые рыбы, относящиеся к роду гольцов. Обычный арктический голец полиморфен, настолько изменчив, что у одного из систематиков вырвалось признание: «Если вы начали выделять виды гольцов, вы уже не знаете, где вам остановиться». Другой вид, относимый к тому же роду, — кунджа — удивительно стабилен. Кунджа, где бы она ни жила, всегда остается кунджей — проходной рыбой с крупными желтовато-белыми пятнышками. Даже существование жилых, не выходящих из рек в море форм — вещь для лососевых обычная, для кунджи строго не доказана.
Но чем объяснить стабильность одних видов и лабильность других? Тут уже мы переходим от фактов к спекулятивным гипотезам. Одной из них я хочу с вами поделиться.
Среди генетиков долго бытовал взгляд на мутации как на явление неизбежное и неотвратимое. Это не совсем так: если мутация затронула только одну из цепей двойной спирали ДНК, существующие в каждой клетке специальные ферменты — репаразы спешат исправить ошибку, вырезая испорченный участок и пристраивая новый, комплементарный оставшейся цепи. Разумеется, они не всесильны (хотя американский биофизик Р. Сетлоу полагает, что они исправляют до 99 % вновь возникших мутаций). Во-первых, они с таким же успехом «узаконивают» ошибку, переделывая не изменившуюся, а оставшуюся прежней цепь. Во-вторых, они бессильны, когда мутация затрагивает обе цепи. Нельзя ли допустить, что повышенная изменчивость одних видов по отношению к другим объясняется тем, что у лабильных видов, в результате того же мутационного процесса, снижена активность репараз? Проверить это предположение методически трудно, но — в отличие от «макромутаций» Гольдшмидта — не невозможно.
Беда в том, что ретроспективно мы можем с тех или иных позиций объяснить какое-либо эволюционное явленна, но не можем объяснить, почему оно, несмотря на, казалось бы, благоприятствующие этому факторы, не произошло. Почему кунджа не образовала разнообразных форм? Почему щука на всем протяжении Евразии остается щукой, исключая сомнительный вид — щука амурская, а на американском континенте она вдруг проявляет вспышку изменчивости, образуя хорошо различимые виды? По-видимому, каждый подобный казус требует отдельного решения. Причины могут оказаться разными.
Гипотезы о скачкообразном возникновении видов, а то и групп более высокого ранга неоднократно высказывались палеонтологами. Причины этого понятны. Новый, возникающий вид, как правило, имеет низкую численность и малый ареал. Если же он, к тому же, обитает в условиях, не способствующих захоронению органических остатков, мы из-за неполноты палеонтологической летописи упускаем момент его становления и находим в земных слоях уже «в готовом виде» полностью сформировавшимся. Однако известны и хорошие, так называемые филетические ряды, показывающие, что возникновение Ридов и родов идет постепенно. Как указывает известный американский палеонтолог Дж. Симпсон, такая постепенность — факт, независимо от того, всегда ли она наблюдается. Всесторонний анализ многих палеонтологических материалов позволил Симпсону прийти к выводу, что термины макро- и микроэволюция условны. Существует единый процесс эволюции, осуществляемый совокупностью факторов, о которых мы уже говорили. Это отбор, генетико-автоматические процессы, изоляция и последующий за ней инбридинг.
Прямолинейна ли эволюция?
Я уже упоминал в главе о прогрессе, что иногда стратегия игры Вид — Среда прямолинейна, поэтому эволюция приобретает вид направленного процесса. Такие, известные из палеонтологического материала факты время от времени вызывали к жизни разные модификации ортогенетических теорий, в конечном счете сводимые к «закону градаций» Ламарка.
Сторонников ортогенеза (прямой, направленной эволюции) далеко не всегда следует зачислять в идеалисты. Часто это прямолинейные последователи Демокрита, не желающие признавать эволюцию вероятностным процессом, отбором случайных мутаций. Причины ортогенеза (термин, предложенный еще в 1893 году В. Гааке и позже Г. Эймером) выдвигались разные: и направленное влияние среды, и особенности, заложенные в самих организмах. Однако все ортогенетики согласны в одном — в отрицании дарвиновского принципа отбора ненаправленных наследственных изменений. Некоторые из трактовок в настоящее время выглядят уже несколько наивно. Палеонтолог О. Абель, например, считал, что в эволюции действует некий аналог первого закона Ньютона («всякое тело находится в состоянии покоя или равномерного прямолинейного движения…»). Но ведь эволюционирующий вид — не брошенный камень и законам механики не подчиняется.
Другие прямо вводят в эволюцию некий нематериальный, порой с явно теологической окраской элемент, изгоняющий из нее неприличную случайность. Таким был известный французский антрополог Тейяр де Шарден (его живо написанная книга «Феномен человека» издавалась у нас). Оттого, что книги де Шардена были запрещены Ватиканом, материалистической его точка зрения не стала.
Крупнейший советский ихтиолог Лев Семенович Берг, о котором специалисты по рыбам до сих пор говорят с почтением (вполне им заслуженным), в 1922 году выпустил книгу «Номогенез, или эволюция на основе закономерностей». Название ее само говорит за себя — случайность из эволюции изгоняется, в ней царит закономерность, необходимость. Необходимость эту Берг видел в «химическом строении протоплазмы» и «направленном влиянии ландшафта». Вряд ли подобные объяснения сейчас кого-либо удовлетворят, они находятся на уровне натурфилософского «закона градаций» Ламарка. Я, например, не могу понять механизма действия ландшафта на вид. Да и никто не сможет.
Видный наш энтомолог А. А Любищев до конца жизни оставался противником дарвиновского истолкования эволюции; он иронически писал, что «естественный отбор в мировоззрении неодарвинистов исполняет обязанности всемогущего бога». Однако ирония — не лучший аргумент в научном споре. Любищев утверждал, что закон гомологических рядов Вавилова опровергает дарвинизм. Вряд ли это так — параллелизм изменчивости, как мы видели, вполне убедительно объясняется с позиций биохимической генетики, а молекулярная генетика во всех деталях это истолкование подтверждает. Вообще в науке любая теория может быть побеждена лишь новой теорией, лучше объясняющей факты. Теория естественного отбора до сих пор удивительно логично объясняла всю совокупность фактов, накопленных самыми разными отраслями биологии. Менять ее на смутные соображения о непознанных нами закономерностях эволюции было бы верхом неблагодарности.
Поэтому в настоящее время большинство эволюционистов считает ортогенез синонимом ортоселекции. Если отбор на протяжении филогенетического ряда сменяющих друг друга видов действует в одном направлении, возникает направленное изменение признака. Мы уже рассматривали ряд таких примеров — в частности, эволюцию мезозойских динозавров и эволюцию лошади. К примерам того же рода относится и увеличение в процессе эволюции размеров слонов, увеличение клыков у саблезубых тигров, объема мозга у предков человека и многое другое.
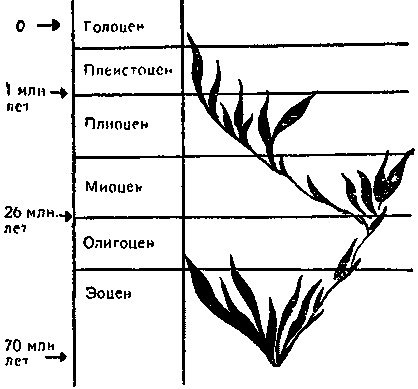
Эволюция лошади. На протяжении 70 миллионов лет произошли по меньшей мере три вспышки видообразования. Эволюция шла по разным направлениям, но игру со средой выиграл лишь род аквус (зебры, ослы, лошади).
Следует, впрочем, оговориться, что не один отбор приводит к направленному изменению признака. Увеличение процента носителей группы крови В от берегов Атлантики к центру Евразии, несомненно, действие отбора, осуществлявшегося возбудителем оспы. Оспа до открытия вакцины Дженнера была весьма жестким фактором отбора в человеческих популяциях. А вот потеря группы крови В американскими индейцами, скорее всего, следствие генетико-автоматических процессов, дрейфа генов. Советский антрополог Ю. Г. Рычков полагает, что такое подобие направленного изменения частоты генов, имитирующее действие отбора, встречается весьма часто (нужно лишь добавить, что дрейф направленно изменяет признаки, если в это не вмешивается отбор).
Сторонники ортогенеза нередко ссылаются на действительно любопытные факты, когда вымершие виды обретали признаки, на первый взгляд явно неприспособительные. С точки зрения ортогенеза эти факты объясняются просто: филогенетический ряд видов, развивающийся согласно какой-то заложенной свыше тенденции, заводит какой-либо признак за оптимум, что является причиной вымирания. Так бильярдный шар после чересчур (ильного удара может вылететь за борт. Излюбленный пример орто-генетиков — гигантский олень мегацерос, у которого рога были чуть ли не больше его самого. В ту же категорию относят клыки саблезубых тигров и причудливые образования — не то вооружение, не то украшение — многих динозавров, рога ископаемых копытных — титанотериев, чрезмерно развитые клыки поздних мамонтов и т. д. Логика проста: эти признаки не кажутся мне приспособительными, значит, естественный отбор не мог их создать.
Справедливости ради следует отметить, что ламаркизм, учение о врожденной приспособляемости, перед этими фактами на самом деле бессилен.
А дарвинизм? Прежде всего нужно показать, что эти гипертрофированные признаки (гиперморфоз, как называл это направление эволюции Шмальгаузен) действительно неприспособительны, неадаптивны. Многие животные, до сих пор благоденствующие, имеют столь же нелепые с человеческой точки зрения образования, и это не мешает им выстоять в борьбе за существование. Однако изменись резко условия жизни (измени среда стратегию), и они должны вымереть, если не успеют измениться сами.
Некоторые весьма любопытные соображения по этому поводу мы сейчас рассмотрим.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Естественный отбор
Естественный отбор Итак, Дарвин показал, что основным средством создания всех пород домашних животных является искусственный отбор. Даже в то далёкое время, когда люди вели этот отбор, не ставя перед собой определённой цели, бессознательно, они достигали поразительных
Естественный отбор
Естественный отбор Я не усматриваю предела деятельности этой силы, медленно и прекрасно приспособляющей каждую форму к самым сложным жизненным отношениям. Ч. Дарвин Осы, бабочки и дарвинизмВ предыдущих главах мы неоднократно говорили о естественном отборе. Это и
Естественный отбор в природе и в лаборатории
Естественный отбор в природе и в лаборатории Действие отбора изучают не только в лабораторных экспериментах, но и в ходе многолетних наблюдений в природе. Первый подход позволяет контролировать условия среды, выделяя из бесчисленного множества реальных жизненных
ЕСТЕСТВЕННЫЙ ОТБОР: НЕ ВСЕ В ЭВОЛЮЦИИ ДВИЖЕТСЯ ВПЕРЕД
ЕСТЕСТВЕННЫЙ ОТБОР: НЕ ВСЕ В ЭВОЛЮЦИИ ДВИЖЕТСЯ ВПЕРЕД Шок Дарвина Леонардо да Винчи (1452–1519) говорил: «В природе нет ошибок, но знай: ошибка есть в тебе». Слишком совершенным казалось этому гению все богатство и разнообразие форм в природе, чтобы хоть немного усомниться в
ЕСТЕСТВЕННЫЙ ОТБОР
ЕСТЕСТВЕННЫЙ ОТБОР Естественным отбором считается процесс, который Дарвин назвал «борьба за существование», при котором наиболее приспособленные организмы выживают, а наименее приспособленные погибают. Согласно положениям дарвинизма, естественный отбор в популяции с
ЕСТЕСТВЕННЫЙ ОТБОР ПОД ВЛИЯНИЕМ ХИЩНИКОВ
ЕСТЕСТВЕННЫЙ ОТБОР ПОД ВЛИЯНИЕМ ХИЩНИКОВ Чарлз Дарвин осознавал важность размера популяции при определении выживаемости индивидов, соревнующихся между собой за ограниченное число ресурсов удовлетворения основных потребностей, в особенности за пищу. В этом процессе
ЧТО МОЖЕТ ГРУППОВОЙ ЕСТЕСТВЕННЫЙ ОТБОР
ЧТО МОЖЕТ ГРУППОВОЙ ЕСТЕСТВЕННЫЙ ОТБОР Групповой брак приводит к близкородственному скрещиванию и через несколько поколений делает всех членов группы близкими по набору генов. В такой ситуации не столь важно, мое или твое потомство выжило, я или ты погиб преждевременно.
5.3. Естественный отбор и развитие этики сексуальных контактов
5.3. Естественный отбор и развитие этики сексуальных контактов Сила и продолжительность половой любви бывают такими, что невозможность обладания представляется обеим сторонам великим, если даже не величайшим несчастием; они идут на огромный риск, даже ставят на карту
7. ВОЙНЫ И ЕСТЕСТВЕННЫЙ ОТБОР
7. ВОЙНЫ И ЕСТЕСТВЕННЫЙ ОТБОР Высочайшее счастье для человека и его величайшая радость — разбить и уничтожить врага, стереть его с земли, забрать все, что он имел, заставить плакать его жен, скакать на его лучших и любимых лошадях и обладать его красивыми
Что может групповой естественный отбор
Что может групповой естественный отбор Групповой брак приводит к близкородственному скрещиванию и делает через несколько поколений всех членов группы близкими по набору генов. В такой ситуации не столь важно, чье — мое или твое — потомство выжило, я или ты погиб
9. Естественный отбор – главная движущая сила эволюции
9. Естественный отбор – главная движущая сила эволюции Вспомните!Какие виды отбора вы знаете?Назовите известные вам формы естественного отбора.Естественный отбор – это преимущественное выживание и размножение наиболее приспособленных особей каждого вида и гибель
Естественный отбор — быть сильнее своего животного начала
Естественный отбор — быть сильнее своего животного начала Нам же особенно важно, что это комендант своей силой заставляет тело следовать инстинктам. (Не пропустите этот момент!) То есть это комендант (его сила) определяет животное начало в теле. И с точки зрения физики
Глава 12 Естественный отбор: кто выживет?
Глава 12 Естественный отбор: кто выживет? Есть горстка ученых, пытающихся выявить признаки, общие для всех когда-либо существовавших человеческих культур и цивилизаций. Эти ученые роются в трудах этнографов и в статьях антропологов в поисках различий между племенами и
Естественный отбор — единственный направленный фактор эволюции
Естественный отбор — единственный направленный фактор эволюции Несомненно, важнейшим эволюционным фактором является естественный отбор. При определении естественного отбора Ч. Дарвин использовал понятие «выживание наиболее приспособленного». При этом имелось,
Естественный отбор и феногеография
Естественный отбор и феногеография Изучение естественного отбора — одна из важнейших задач в изучении микроэволюции. Без глубокого понимания действия этого единственного направленного эволюционного фактора ни о каком переходе к управляемой эволюции не может быть и