Великий синтез
Великий синтез
Как связать эволюцию с генетикой?.. Можно ли подойти к вопросам изменчивости, борьбы за существование, отбора — словом, дарвинизма, исходя не из тех совершенно бесформенных, расплывчатых, неопределенных воззрений на наследственность, которые только и существовали во время Дарвина и ею непосредственных преемников, а из твердых законов генетики?
С. С. Четвериков

Квадрат суммы двух чисел, или уравнение Харди — Вейнберга
В двадцатых годах нашего столетия в биологии сложилось странное положение. Уже два десятка лет стремительно развивалась новая наука — генетика. Два фактора эволюции по Дарвину — наследственность и изменчивость — изучались на строго научной основе. Умозрительные гипотезы сменялись точными экспериментами, интуитивные догадки — алгебраическими выкладками. И весь этот могучий поток мысли и деятельности протекал мимо той отрасли биологии, помочь которой он должен был в первую очередь — мимо теории эволюции. Более того, находились люди, которые обвиняли генетику в том, что она якобы не согласуется с дарвинизмом. С другой стороны, наиболее рьяные последователи и продолжатели Менделя полагали, что дарвинизм устарел.
Этот мнимый конфликт, отголоски которого и сейчас можно встретить в литературе, был разрешен еще 40 лет назад замечательным советским биологом Сергеем Сергеевичем Четвериковым. Именно он положил начало новой, так называемой синтетической эволюционной теории, объединившей дарвинизм и генетику.
История нашей генетики изобилует драматическими положениями и яркими, выдающимися личностями. Но и на этом, богатом героями и событиями фоне, фигура Четверикова выделяется особенно ярко.
В бессмертие он вошел как генетик — и немногие знают, что прежде всего он был энтомологом — специалистом по чешуекрылым. Этакий кузен Бенедикт из романа Жюля Верна, ученый не от мира сего — и в то же время член Московского стачечного комитета в период революции 1905 года. Мало кто работал так напряженно и плодотворно, как Четвериков, однако в силу многих внешних обстоятельств, и главным образом — из-за своей исключительной добросовестности (хочется сказать — излишней, если бы это качество могло быть излишним) печатался он немного. Известно лишь около 30 его работ. В наше время «информационного взрыва», когда научные статьи порой не читают, а считают, иной аспирант имеет больше публикаций. Но одна лишь работа Четверикова — «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926) перевешивает по значению сотни, если не тысячи, подобных скороспелых трудов. День ее опубликования — один из немногих «звездных часов» науки. Недаром доклад Четверикова на V Международном генетическом конгрессе в Берлине в 1927 году был встречен восторженно. Докладчика чествовали, как триумфатора, а потом, как это ни странно, забыли на ряд десятилетий. Лишь недавно, уже после смерти Четверикова, последовавшей в 1959 году, его имя всплыло из безвестности, работа 1926 года была переведена в ряде стран, и теперь от нее выводится новая наука — эволюционная и популяционная генетика. Как говорится, неисповедимы пути господни, но судьбы научных открытий порой еще более непредсказуемы.
Что же все-таки сделал Сергей Сергеевич Четвериков?..
Работа Четверикова имела предшественников. Еще в 1904 году К. Пирсон математически строго показал, что наследуемость мутаций по менделевскому принципу снимает опаснейшее для дарвинизма возражение Дженкина. В самом деле, ведь один ген не «разводится» другим. В гетерозиготном состоянии они сосуществуют, оставаясь раздельными. Мутация не «заболачивается», как писал Дженкин, скрещиванием, и новый признак всегда имеет шанс проявиться в фенотипе в неизмененном состоянии.
Через четыре года после работы Пирсона английский математик Г. Харди и независимо от него немецкий генетик В. Вейнберг сформулировали положение, которое сейчас именуется законом Харди — Вейнберга и описывает частоту встречаемости генов в равновесной популяции свободно скрещивающихся организмов. (Под равновесной популяцией подразумевается сообщество особей, внутри которого соотношение генов не изменяется, находится в равновесии.)
До сих пор я делал воистину героические попытки избежать математических формул. К сожалению, здесь этого сделать уже нельзя. Читателям, не жалующим математику, можно сказать в оправдание и утешение, что мы постараемся ограничиться азами, известными из курса средней школы.
Допустим, если частота встречаемости доминантного гена А в популяции равна q, а рецессивного а, соответственно, 1 — q, то соотношение гомо- и гетерозигот будет следующим:
q2AA +2q(1 — q)Aa + (1 — q)2аа.
Это известная из школьной алгебры формула квадрата суммы двух чисел. Если q = 0,5, как в случае первого поколения менделевских гибридов, то во втором поколении мы имеем следующее соотношение:
0,25AA + 0,5Aа + 0,25aа,
а так как Аа в случае полной доминантности фенотипически выражены как A, мы получаем менделевское расщепление — 0,75:0,25, то есть 3:1.
Физики-теоретики обычно допускают при выводе формул ряд облегчающих им работу предположений (их называют условиями, требованиями или ограничениями). В результате получаются абстракции вроде «идеального газа» и «идеальной жидкости». Закон Харди — Вейнберга также описывает некую абстрактную «идеальную» популяцию со свойствами, для реальных популяций невозможными:
1. Численность такой популяции должна равняться бесконечности. Это, впрочем, не столь жесткое требование, как кажется на первый взгляд. Статистики сплошь и рядом используют при описании частоты встречаемости какого-либо объекта так называемую кривую нормального распределения (кривую Гаусса), хотя, строго говоря, она годится лишь для бесконечно большой выборки. При этом в расчеты вносится ошибка, которая так и называется — ошибка выборки.
Понятие выборки требует пояснения. Дело в том, что статистик почти всегда не может исследовать изучаемые объекты все до единого (так называемую генеральную совокупность). Допустим, вы работаете дегустатором и должны оценить, каким сортом выпустить партию из тысячи банок консервов. Разумеется, вы не можете перепробовать их все — ведь их должны есть другие, в противном случае ваша работа потеряет смысл. Поэтому приходится судить об исследуемой совокупности по незначительной части ее; риск ошибки остается, но вы можете его уменьшить, увеличивая выборку.
В случае с отношением генов ошибка, отклонение от заданной уравнением величины характеризует саму популяцию. На конечной круглой Земле нет популяций бесконечно больших. Численность стада саранчи или большого стада сельди можно без особого греха приравнять к бесконечности. А вот популяция, например, зубров в заповеднике или окуней в маленьком замкнутом пруду — отнюдь не бесконечность. В дальнейшем мы убедимся, насколько важны для эволюционного процесса подобные отклонения.
2. Скрещивание особей внутри популяции полностью свободно, без всяких ограничений (вероятность скрещивания каждого члена популяции с любым другим одинакова). Такие популяции называются панмиксными (свободно смешивающимися). Это условие гораздо более спорно. Человек, например, свободнее всех животных перемещается по планете, однако вероятность того, что житель Москвы женится на москвичке, наверняка гораздо выше, чем вероятность брака москвича с жительницей Иркутска или Караганды. В природе же возможность скрещивания особей из разных частей достаточно большой популяции практически равна нулю. При этом не нужно путать два понятия — популяцию и вид. Один и тот же вид может быть распространен чрезвычайно широко, практически всесветно. Я помню, как был поражен, увидев одну из банальнейших наших стрекоз — желтую стрекозу, порхающую над болотцем на коралловом острове архипелага Гилберта. Но весь этот огромный ареал распадается на ряд изолированных полностью и частично друг от друга популяций. Размеры их могут быть весьма малыми, но даже в самой маленькой популяции панмиксность не абсолютна. Так что второе ограничение серьезнее первого.
3. В описываемую уравнением Харди — Вейнберга популяцию не должны попадать новые гены из смежных популяций; она должна быть абсолютно изолированной.
Такие условия осуществляются в природе в весьма редких случаях. Известны реликтовые популяции, оторванные от основного ареала вида; порой только они и остаются в убежищах — рефугиумах, когда вид повсеместно вымирает. Бывает и наоборот — недавно возникший вид, не успев расселиться, какое-то время оказывается единой популяцией. Новым генам в таких случаях просто неоткуда попадать. Чаще изоляция не бывает абсолютной. Она нарушается главным образом миграцией подрастающего молодняка у животных, заносом семян или пыльцы у растений. Степень изоляции может быть весьма различной. Крысы, например, живут семьями, происходящими от одной родительской пары, и загрызают любого забредшего к ним чужака. Австралийские сороки гнездятся замкнутыми общинами; внутри общины самцы спариваются со всеми самками, самки откладывают яйца в общее гнездо и насиживают их по очереди, но залетные особи в общину не принимаются (предел панмиксности и изоляции одновременно!). А вот гориллы, по наблюдениям американского зоолога Г. Шаллера, кочуют по горным лесам экваториальной Африки семьями, возглавляемыми взрослыми, поседевшими самцами, которые вполне терпимо относятся к визитам гостей из других групп.
4. Новые аллели генов не должны возникать и внутри такой популяции. В ней не должно возникать новых мутаций! Ясно, что такое условие, пожалуй, чересчур жестковато. Все же, если мы исследуем распространение в популяции лишь одного гена в двух аллельных формах, принять его можно. Ведь частота повторного возникновения одной и той же мутации чрезвычайно низка; при ограниченной численности популяции можно принять, что каждая аллельная форма гена возникает только один раз. Разумеется, это допущение годится лишь для короткого ряда поколений (вероятность смерти человека от удара молнии ничтожна, но проживи он несколько тысяч лет — и эта смерть постигнет его со 100-процентной вероятностью).
5. Пятое ограничение кажется весьма существенным. Именно для того, чтобы распределение генов в популяции соответствовало уравнению Харди — Вейнберга, естественный отбор по признакам, определяемым доминантным и рецессивным аллелем, должен отсутствовать. Мутации должны быть нейтральными, иными словами, гомозиготы АА и аа и гетерозиготы Аа должны иметь одинаковую жизнеспособность и плодовитость, одинаковую вероятность оставить потомство.
О возможности существования нейтральных мутаций и их роли в эволюции мы успеем поговорить в другом месте. Пока же нам остается признать, что идеальная популяция Харди — Вейнберга вряд ли когда-либо существовала. В нашем мире нет идеала — все популяции конечны, большая часть их недостаточно хорошо изолирована от соседних и в то же время каждая из них недостаточно однородна внутри себя, все гены мутируют, и многие мутации имеют селективное значение.
Однако применение формулы Харди — Вейнберга к реальным популяциям вполне допустимо (разумеется, с учетом возможных отклонений за счет ограниченности популяции, частичной изоляции внутри нее, заноса новых генов, возникновения новых мутаций и действия отбора). Здесь возникает полная аналогия с физическими моделями вроде идеальной жидкости, успешно «работающими» в границе своих возможностей. Пользуясь этой формулой, можно рассчитать, например, как будет убывать в популяции частота встречаемости летального рецессивного гена, вызывающего 100-процентную смертность гомозигот аа. Оказывается, что даже при столь жестком отборе смертоносный рецессивный ген сохраняется сотни поколений; выщепляясь постепенно из гетерозигот. Иное дело — доминантные летальные мутации. Они из популяции исчезают, естественно, на протяжении одного поколения — ведь они не могут скрыться в гетерозиготе! Исключения из этого правила редки. Такова, например, наследуемая доминантно хорея Гентингтона — тяжелейшее заболевание нервной системы человека, которое может тянуться многие годы, заканчивается мучительной смертью, но проявляется лишь в зрелом возрасте (30–45 лет), уже после того, как носитель этого гена передал его своим детям. Такие случаи, быть может, встречаются чаще, чем это кажется. Есть оригинальная гипотеза, объясняющая старение наличием в геноме отвечающих за это доминантных генов. Мы стареем и умираем, чтобы освободить место для подрастающих поколений, ибо без смены поколений не было бы и эволюции.
Обсуждение этого вопроса завело бы нас слишком далеко. Поэтому вернемся к синтезу менделевской генетики и классического дарвинизма. Четвериков в 1926 году показал, что именно те ограничения, которые отличают харди-вейнберговскую популяцию от реальной, и делают возможным процесс эволюции. Рассмотрим же факторы эволюции по Четверикову.
Как возникает разнообразие
Из предыдущих глав вы уже знаете, что гены не абсолютно стабильны. Меняется по целому ряду причин структура отдельных генов, их местоположение в хромосомах, и сам хромосомный набор претерпевает всевозможные изменения — вплоть до многократного умножения. Судьба этих изменений оказывается различной.
Многие из них приводят к появлению нежизнеспособных форм, погибающих на той или иной стадии развития. Другие как бы воздвигают вокруг носителей таких мутаций «китайскую стену» нескрещиваемости. В большинстве случаев подобные отщепенцы сходят с жизненной арены, оставляя бесплодное потомство. Наконец, есть и такие, которые включаются в генный фонд и передаются из поколения в поколение. Они претерпевают при каждом мейозе все перипетии генетической рекомбинации — и в конечном счете рано или поздно, оказываясь в гомозиготном состоянии, проявляются в фенотипе. Вид на протяжении своей истории непрерывно обогащается мутантными аллелями. По образному выражению Четверикова, вид, как губка, всасывает мутации (он, правда, предпочитал другой термин — геновариации, в практике не прижившийся).
Чем же, однако, объяснить тот факт, что виды на протяжении многих поколений остаются стабильными, сохраняют, как говорят, «дикий тип»? Напомним, что большинство мутаций рецессивно. Каждая мутация — явление достаточно редкое; лишь после того, как она размножится в последующих поколениях потомков, вероятность встречи двух одинаковых генов в зиготе возрастет. Селекционер может ускорить этот процесс: практически все породы домашних животных и сорта культурных растений гомозиготны по разным рецессивным генам. Скрестив два разных сорта их, мы нередко получаем в потомстве возврат к прежнему «дикому типу». Это знал и Дарвин, скрещивавший голубей разных пород и получавший в потомстве «сизарей». Механизм здесь тот же, что и в случае супрессорных мутаций, о которых мы уже упоминали, — дефект одного генома исправляется другим.
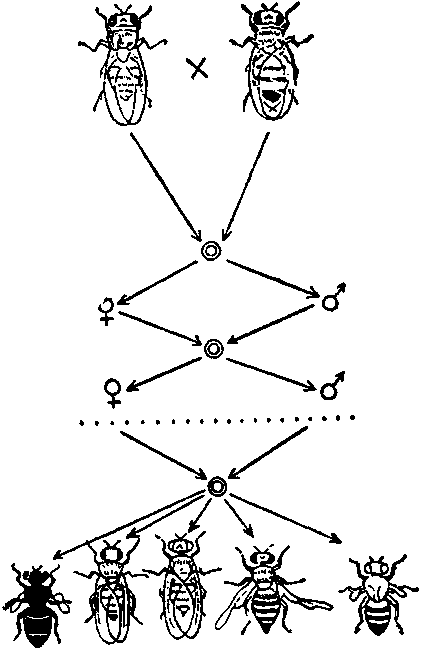
Схема опыта С. С. Четверикова. В потомстве одной дрозофилы после ряда поколений близкородственного разведения выщепляются рецессивные мутантные гомозиготы, до того в скрытой виде находящиеся в однообразном, «диком типе».
При внешнем фенотипическом однообразии геномы видов настолько обогащены скрытым разнообразием, что, остановись мутационный процесс, стань гены абсолютно стабильными, эволюция все равно не прекратится. Реализовать в фенотипах уже накопленное генетическое разнообразие в сколько-нибудь обозримые сроки невозможно.
Из построений Четверикова вытекает важное следствие. Наибольшее значение для эволюции имеют лишь те изменения генетического материала, которые, находясь в рецессиве, не проявляются в фенотипе. Не подвергаясь действию отбора, они могут накопиться в популяциях в достаточном количестве. Из этого правила есть, разумеется, исключения. Бактерии и сине-зеленые водоросли, то есть организмы с неоформленным ядром, не имеющие фазы мейоза, гаплоидны. Они имеют только один набор генов, поэтому каждая мутация немедленно проявляется в фенотипе, у них нет запасов генетической изменчивости. Образно выражаясь, они не помнят прошлого и не готовятся к будущему, живя сегодняшним днем. Бурный темп их размножения позволяет им выстоять в жизненной борьбе. Однако они, приспособляясь, не эволюционируют, оставаясь и в наши дни практически теми же, как и в то время, когда были единственными обитателями нашей планеты. Недаром и бактерии, и даже вирусы выработали свои, оригинальные механизмы генетической рекомбинации, которым, однако, далеко до мейоза и полового процесса высших организмов.
Не каждая мутация, вновь возникнув, может закрепиться в потомстве. Ведь не каждый ее носитель доживает до половозрелости. Существует, помимо накопления мутаций, обратный процесс их потери, обеднения генного фонда. Вероятность потери мутации можно рассчитать: если мы примем, что из потомства одной самки выживают двое, то есть численность популяции не увеличивается и не уменьшается (то, что немцы называют система двух детей — Zweikinder-system), то примерно 37 шансов из ста за то, что эта мутация будет потеряна[9]. В создании генного фонда популяции участвуют две противоборствующие тенденции.
Можно ли утверждать, что мутационный процесс наряду с генетической рекомбинацией сможет обеспечить нормальный ход эволюции, то есть возникновение новых видов, приспособленных к окружающей среде? Напомним, что аналогичные идеи развивал Де Фриз. Однако считать возникновение генетического разнообразия единственным двигателем эволюции нельзя. Мутационный процесс равнодушен к судьбам организмов, геномы которых он изменяет. Приспособленность к условиям внешней среды он создать не может.
«Волны жизни», «горлышко бутылки» и «принцип основателя»
Наряду с мутационным процессом второй фактор эволюции по Четверикову — колебания численности популяций, которые он образно называл «волнами жизни».
Каждый из нас наблюдал «волны жизни» воочию. Ранней весной редко можно увидеть случайную перезимовавшую между оконными рамами муху, а в августе мы страдаем от их назойливости. Год на год не приходится — в одно лето нет житья от комаров, а в другое они редки. Вспышки численности некоторых видов имеют воистину планетарный характер. Таковы, например, массовые миграции саранчи в годы ее усиленного размножения, годы «мышиной напасти», путешествия многочисленных стад полярных пеструшек — леммингов.
Некоторые из этих вспышек цикличны: они имеют сезонный, годовой или многолетний период (часто, хотя порой и без достаточного обоснования связываемый с солнечной активностью). Другие же апериодичны. Это вспышки численности организмов, попавших в новую среду обитания без достаточного количества врагов, эпидемии гриппа и иных заболеваний, с быстротой пожара распространяющиеся по планете нашествия вредителей из других стран.
Колорадский жук у себя на родине, в Северной Америке, никогда не достигал высокой численности, питаясь дикими растениями из семейства пасленовых. Печальную славу злостного вредителя он завоевал на европейских картофельных полях. То же можно сказать и об американской виноградной тле — филлоксере. Континент Евразии не остался, впрочем, перед Америкой в долгу, «наградив» ее непарным шелкопрядом, воробьями и длинным списком сорняков. Вспомним также нашествие кроликов и кактусов опунций на Австралию.
Отнюдь не всегда такие подъемы численности видов вредоносны. Бурное развитие в Каспийском море акклиматизированного там азовского червя — нереиса значительно увеличило кормовую базу рыб. Это в свою очередь компенсировало оскудение Каспия, наступившее в результате зарегулирования стока Волги. Не будь нереиса, осолонение Северного Каспия нанесло бы каспийским рыбам непоправимый ущерб. Никто не может возразить и против «нашествия» на нашу страну американской мускусной крысы — ондатры, расселившейся от Камчатки до Мурмана, от северной Сибири до Средней Азии (правда, по некоторым данным, она вытесняет ценного зверя — выхухоль). И все же испортить неразумным вселением сложившуюся структуру сообществ растений и животных гораздо легче, чем улучшить ее. Нелишне напомнить, что акклиматизации нереиса в Каспии предшествовала многолетняя работа. Сейчас с развитием авиации, облегчающей перевозки, порой случается и такое, что сначала акклиматизируют, а потом скромно уступают друг другу честь этого замысла. Дело доходит до курьезов, когда вместе с ценными объектами (а то и вместо них) завозят совсем нежелательные. Так, вместе с ценными растительноядными рыбами в Среднюю Азию завезли головастиков чернопятнистой лягушки с Дальнего Востока (уж этого-то можно было избежать!).
Много можно было бы рассказать интересного и о причинах таких колебаний численности, когда в период максимума она больше, чем в период минимума в миллионы раз. Обычные океанские волны никогда не достигают такой высоты, как «волны жизни»! Некоторые из них имеют характер автоколебательного процесса, как в генераторах радиоволн. Так, в период вспышки численности мышевидных грызунов прежде изолированные маленькие популяции сливаются в одну большую, возрастает вероятность контактов. Это приводит к быстрому распространению болезнетворных бактерий (например, чумы и туляремии), массовым эпизоотиям и резкому спаду численности. Гребень волны сменяется провалом, популяция колеблется, как маятник.
Однако сейчас нас интересует эволюционное значение таких колебаний. В первую очередь «волны жизни» резко расшатывают генофонд популяции, изменяя численность всех аллелей, разрушая сложившееся харди-вейнберговское отношение их. А это приводит в конце концов к изменению свойств популяции — так расшатывание выпадающего зуба облегчает процесс избавления от него. При спаде «волны жизни» численность одних мутаций возрастает, а значит, возрастает вероятность встречи рецессивов в гомозиготах; скрытые аллели проявляются в фенотипах и подпадают под действие естественного отбора. Новая «волна жизни» будет состоять из особей с измененным генофондом.
Другие мутации при спаде волны могут просто выпасть из генофонда. Этот эффект образно называют «горлышко бутылки» (или эффект бутылочного горлышка) — далеко не все аллели могут пройти к новым поколениям через минимум численности. Например, если при спаде численности лисиц из популяции исчезнут особи — носители генов черно-бурой окраски, чернобурок в этой популяции уже не будет.
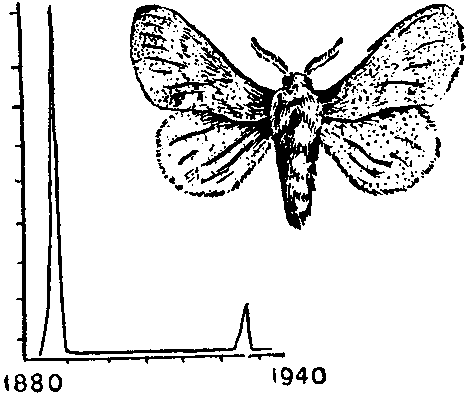
Пример «волн жизни». Колебания численности зимующих гусениц соснового коконопряда за 60 лет.
Не менее важно и то обстоятельство, что при подъеме «волны жизни» прежде изолированные мелкие популяции объединяются в единую, начинается интенсивный обмен генами. Ареал такой популяции расширяется, периферийные части попадают в новые условия, в которых они еще не обитали. Эти enfants perdu (потерянные дети — так во французской армии называли сторожевое охранение на передовой) часто становятся предками новых форм, подвидов и видов (разумеется, если им повезет в жизненной борьбе). При спаде волны они нередко оказываются отрезанными от основного ареала и продолжают свое существование с тем генофондом, который у них имеется. А он, как правило, неадекватен общему генофонду всей популяции. Следующий подъем численности восстанавливает прерванную связь, но соединившиеся популяции порой уже «не узнают» друг друга, они уже генетически изолированы.
Наиболее четко это проявляется в феномене, описанном Четвериковым, который много позже эволюционист-теоретик Э. Майр назвал «принципом основателя». Нередко новый ареал (например, отдаленный остров) заселяют случайно занесенные туда несколько единичных особей нового вида. Так, вся западноевропейская популяция ондатры ведет начало от всего лишь пяти особей, выпущенных в начале века вблизи Праги. Пределом начала популяции, по-видимому, может быть одна оплодотворенная самка. Естественно, она одна, и даже несколько особей, не может хранить весь генофонд родительской популяции. Эволюция этого вида на новом месте должна пойти по-другому. Наряду с изменившимися условиями существования «принцип основателя» приводит к формированию островных рас, подвидов и видов.
В последнее время группа японских исследователей детально изучала популяции крыс на многих островах Океании. Эти грызуны — подлинный бич романтических атоллов — успели с момента вселения пройти большое расстояние по пути дивергенции (расщепления). Некоторые оказались уже генетически изолированными.
Однако колебания численности популяций сами по себе еще не могут привести к видообразованию, к становлению приспособленных к внешней среде форм. Они изменяют в популяции относительную численность аллелей; это несомненно, но несомненно и то, что эти изменения ненаправленны.
Следующий фактор эволюции по Четверикову имеет, по-видимому, для эволюции еще большее значение.
Изоляция — что это такое?
Как вы помните, одним из условий харди-вейнберговского равновесия генов была панмиксия — одинаковая вероятность скрещивания для всех членов популяции. Поэтому следующим «возмутителем спокойствия» является, по Четверикову, изоляция — ограничение панмиксии, происходящее от самых различных причин.
Какие же это причины? Об одной из них мы уже упоминали — когда при спаде «волны жизни» прежде единая популяция распадается на ряд мелких субпопуляций, изолированных друг от друга пространством. Такие осколки называют изолятами. Они еще более четко разделены, если между ними возникают преграды физического характера. Человекообразные обезьяны, например, не умеют плавать, поэтому для стада горилл любой достаточно большой ручей является непреодолимым препятствием. В последнее время важность пространственной изоляции, видимо, даже переоценивают, но об этом речь у нас пойдет впереди.
Чаще скрещиваемость в популяции ограничивается другими формами изоляции. Одна из них — экологическая изоляция, при которой снижается вероятность встречи готовых к размножению особей. Таковы сезонные расы растений, рыб и многих беспозвоночных — здесь изоляция наблюдается не в пространстве, а во времени. Есть и еще более удивительные примеры сосуществования в одной и той же популяции форм, размножающихся одновременно, но не скрещивающихся. У дрозофилы, например, существует мутация «желтая». Желтый самец у обычных серых самок имеет крайне мало шансов на успех, так как вибрирует крылышками в более медленном темпе, что снижает его привлекательность. Если же у серого самца удалить крылья, он вообще теряет способность стимулировать самок к спариванию. Наоборот, самки дрозофил с удаленными усиками-антеннами теряют свою разборчивость и спариваются с самцами других видов. Таких случаев описано немало для самых разных животных.
Даже если спаривание (или опыление у растений) произошло, оплодотворения нередко не происходит. Это так называемая физиологическая изоляция. У многих растений пыльца просто не может прорастать на рыльцах пестиков одних особей, но прорастает у других. Наконец, последний тип — собственно генетическая изоляция. Оплодотворение в данном случае происходит, но гибриды первого поколения нежизнеспособны или же стерильны из-за нарушений мейоза.
Большинство описанных выше форм изоляции преодолимо (например, искусственным осеменением). Генетическая же изоляция создает между формами уже непреодолимый барьер. Его, однако, не следует считать видовой границей, как это нередко делают. Многие виды не смешиваются с близкими видами только благодаря различиям в брачном поведении. Наоборот, сравнительно легко вывести форму, полностью генетически изолированную от исходной, но имеем ли мы право назвать ее новым видом? Полиплоидия у растений и хромосомные перестройки типа транслокаций у животных — самые верные способы генетической изоляции. Но, как мы уже говорили, изоляция — не само видообразование, а лишь служанка его. Услуги ее, однако, огромны. Скрещивание сглаживает различия внутри популяции, препятствует дивергенции прежде единого вида на внутривидовые группировки, которые в дальнейшем могли бы стать новыми видами. Напротив, изоляция закрепляет в небольших популяциях (субпопуляциях, или, как их порой называют, демах) различия. Без нее не было бы дивергенции — вся популяция если и изменялась бы, то целиком, один вид переходил бы в другой, не порождая новых. Это было бы видопревращение, а не видообразование.
Остановимся еще на одном вопросе — камне преткновения для многих: что же все-таки изоляция — причина или следствие генетических различий? Вдумчивый читатель без труда сообразит, что ответить на этот вопрос однозначно нельзя. Кролики, акклиматизированные на острове Мадейра, весьма существенно изменились и потеряли способность скрещиваться с европейскими. Здесь генетические различия явились побочным следствием накопления их за время раздельного существования популяций.
Бывают и другие случаи, когда ничтожное генетическое различие (например, изменение частоты брачного кваканья у лягушек или же изменение скорости вспышек у светляков) приводит к изоляции, а та в свою очередь раскалывает популяцию — вплоть до видового обособления.
Наконец, возникновение тетраплоида или транслоканта сразу делает невозможным скрещивание его с другими особями популяции. К этому же может привести появление достаточно большой инверсии. Тут есть одна особенность: возникновение хромосомных мутаций — вещь достаточно редкая, причем они появляются в отдельном организме. Такая изоляция чересчур уж хороша, чтобы стать важным путем эволюции. С ней справляются лишь обоеполые растения с их способностью к вегетативному размножению. Авторы, отстаивающие главную роль хромосомных перестроек в видообразовании, обычно начинают во здравие, а кончают за упокой. Сначала они, в согласии с постулатами генетики, принимают, что: а) все хромосомные перестройки единичны (отсюда вытекает, что единицей эволюции должна быть отдельная особь, а не популяция); б) эти перестройки создают абсолютный генетический барьер между изменившимся героем-одиночкой и серой массой прочей популяции. В ходе дальнейших рассуждений приходится принимать, что или перестройки происходят с частотой в тысячи раз большей, чем обнаруживаемая в природе и в эксперименте, или же они не обеспечивают абсолютной изоляции (чему опять-таки достаточных обоснований нет). На мой взгляд, проще признать вслед за Четвериковым, что сама по себе изоляция не создает вида, а, как правило, развивается постепенно, и эволюция идет в результате действия нескольких факторов. О последнем из них — естественном отборе — речь пойдет в следующем разделе.
Двигатель эволюции
Важнейший фактор эволюции — и здесь Четвериков целиком на стороне Дарвина — это естественный отбор. Все факторы, рассмотренные ранее, изменяют численность генов в популяции случайно, не направленно. Только отбор — процесс направленный. Именно он осуществляет обратную связь между условиями обитания и генофондом популяции, накладывает печать полезности на происходящие в популяции изменения.
Один, без других факторов эволюции, отбор бессилен. Вспомните классический опыт Иоганнсена: в чистой линии, где все гены одинаковы, не из чего выбирать. Поэтому прежде всего для эволюции требуется мутационный процесс, поставляющий «сырой материал», а также свободное скрещивание, перебирающее этот материал во всевозможных комбинациях. Однако будь свободное скрещивание (панмиксия) абсолютным, не могли бы происходить дивергенция, распад прежде единого вида на новые, дочерние. Значит, те или иные формы изоляции для эволюции также необходимы. Наконец, будь популяция бесконечно большой, вероятность возникновения рецессивных гомозигот была бы ничтожной, и они не смогли бы попадать под действие отбора. Поэтому необходимы колебания численности конечной по объему популяции. Четыре фактора Четверикова можно сравнить с постулатами, лежащими в основе геометрии. Подобно тому, как из постулатов геометрии можно построить все ее теоремы, исходя из факторов Четверикова, можно описать все процессы видообразования. Этих факторов ровно столько, сколько необходимо для исчерпывающего объяснения процесса эволюции (как говорят математики — необходимо и достаточно).
Многие, если не все ошибочные тенденции в изучении и истолковании процесса эволюции основываются именно на том, что один из факторов превозносят в ущерб прочим. При этом почему-то больше всего достается отбору. Немало исследователей пыталось прикинуть: а нельзя ли исключить отбор из эволюционных факторов? Многим казалось, что им это удалось. Чаще всего отбору отказывают в творческой роли — это, мол, фактор консервативный, отметающий все ненужное, но бессильный сотворить что-либо новое.
Такая точка зрения отнюдь не нова. Вспомним, что аналогичные взгляды развивал еще в прошлом веке основоположник механоламаркизма Г. Спенсер. Действительно, отбор может сохранять в популяции прежнюю норму — наиболее приспособленную для данных условии комбинацию генов (так, на выставках собак получают первые места особи с наименьшими отклонениями от породного стандарта). Классический пример такого отбора приведен в старой работе американского биолога X. Бампуса, который подобрал после жестокой бури со снегом оглушенных и полузамерзших воробьев. Половина из них, несмотря на уход, погибла. Оказалось, что выжили в основном те особи, морфологические признаки которых больше всего приближались к средним, типичным для вида значениям. Это правило распространимо и на человека. Анализ огромного материала из картотек родильных домов убедительно свидетельствует о том, что наибольшая смертность в первые 28 дней жизни наблюдается среди детей, вес которых или много меньше, или же, наоборот, много больше среднего (три килограмма шестьсот граммов). Лишь большие успехи педиатрии в последнее время позволяют спасти значительное число таких детей.
Наш замечательный биолог-эволюционист И. И. Шмальгаузен, обобщив много наблюдений подобного рода, назвал такой тип отбора стабилизирующим. О нем мы еще будем говорить. Но есть и другой отбор — направленный, творческий. Такой отбор проводит селекционер, оставляя на племя организмы с максимальным развитием тех признаков, которые его интересуют (селекционер и означает — отборщик). Этому отбору подвергаются популяции диких растений и животных в периферийных частях ареала, где условия максимально отличаются от обычных. Фактически селекционер, отбирая, например, растения на засухоустойчивость или морозостойкость, почти не отличается от природы (за тем исключением, что, помимо выживаемости в крайних условиях, ему еще нужно получить хозяйственный эффект — зачем превращать пшеницу в кактус?).
Здесь-то творческая роль отбора проявляется во всей полноте. Как справедливо заметил один из виднейших продолжателей дела Четверикова английский генетик Р. Фишер, отбор — «процесс, благодаря которому вероятность некоего случайного события (удачной комбинации генов. — Б. М.), крайне малая, так увеличивается со временем, что крайне маловероятным становится не наличие, а отсутствие этого события». Творческий отбор, как и стабилизирующий, снижает разнообразие в популяции. Но если стабилизирующий сохраняет норму, то творческий — отклонения от нее.
Есть и третья форма отбора, именуемая дизруптивной (от английского to disrupt — разрывать, раскалывать). Такой отбор сохраняет не одну норму, а две или более. Дизруптивный отбор человек применял, создавая, например, из дикой банкивской курицы яйценоские и мясные породы, из дикой лошади — скакунов и тяжеловозов, из волка — болонок и догов. Естественный дизруптивный отбор приводит к дивергенции вида на два и более.
Иногда значение отбора стараются приуменьшить следующими соображениями. Представим относительную приспособленность генотипов в популяции в виде некоей карты, где максимумы приспособленности означают вершины, а минимумы — провалы. Генотип, поднявшийся на один такой пик, не может сменить его на другой, пусть даже повыше прежнего, — для этого ему потребовалось бы пойти против действия естественного отбора. Перескочить низину от пика до пика можно лишь случайным скачком. Английский генетик С. Райт уподобляет популяцию человеку, бредущему по пересеченной местности в тумане с целью забраться повыше. Если такой человек будет идти только вверх, то его путешествие кончится на первом же крохотном возвышении (максимальной адаптации — приспособления к внешним условиям популяция не достигнет).
Такое рассуждение выглядит убедительно; у него лишь один недостаток: оно неверно. Ведь карта «страны приспособленности» не остается стабильной! На деле вершины и низины даже на поверхности Земли изменяются, непрерывно меняясь местами. Комбинации генов, прежде приспособленные, с изменением условий вновь становятся неприспособленными, и стабилизирующий отбор сменяется творческим, а то и дизруптивным (если одна часть популяции будет подниматься на один пик, а другая — на другой). Адаптивное поле — не карта горной страны, скорее это толчея волн в центре тайфуна, в которой мечется кораблик эволюционирующей популяции. Это не исключает возможности существования таких условий, которые меняются крайне медленно. Вот тогда-то и возникают так называемые персистентные формы — «живые ископаемые», без видимых изменений переживающие целые геологические эпохи!
Короче — при объяснении целесообразности организмов без естественного отбора не обойтись. Но выполнить свою миссию отбор может лишь при наличии ряда условий, о которых речь пойдет дальше.
Отбор и браки между родственниками
Многие из тезисов Четверикова в наше время кажутся общеизвестными, тривиальными. Следует, однако, помнить, что он пришел к этим выводам в 1926 году. В ту пору уже были известны сотни мутаций дрозофилы — классического объекта тогдашней генетики, но все они были открыты при искусственном разведении в лабораториях и казались совершенно чуждыми природе, вроде декоративных пород собак — болонок и той-терьеров. Предположить, что нормальные, «дикие» мухи являются носителями этих генов в гетерозиготном состоянии было довольно смелым шагом. Естественно, Четвериков вместе со своими учениками — Б. Л. Астауровым, Н. В. Тимофеевым-Ресовским, С. М. Гершензоном, П. Ф. Рокицким, Д. Д. Ромашовым, Е. И. Балкашиной (все они впоследствии внесли огромный вклад в развитие разных отраслей генетики) — приступил к экспериментальной проверке гипотезы.
Основной метод, которым они пользовались, фактически заключался в моделировании эволюции по принципу, много позже названному «принципом основателя». Пойманные в природе мушки разделялись на размножавшиеся внутри себя чистые линии — в потомстве одной самки сестры скрещивались с братьями, внуки с внучками и так далее. Что же происходило в популяции?
Чем меньше численность популяции, тем вероятнее встреча двух одинаковых мутантных аллелей в генотипе и проявление их в фенотипе. Рецессивные аллели в многочисленной «дикой» популяции были подавлены доминантными. Но за однообразной гетерозиготной мае кой скрывалось не поддающееся учету разнообразие. Смелая гипотеза проявилась с блеском — однообразный «дикий» тип был насыщен мутациями (по выражению Четверикова — как губка водой). Все естественные популяции оказались гетерогенными.
Такое близкородственное скрещивание давно, с незапамятных времен известно людям. Его называют инбридингом или инцухтом (разведение в себе). Селекционеры нередко использовали его, чтобы закрепить в потомстве признаки выдающегося производителя. В то же время было хорошо известно, что инбридинг связан с возникновением у животных и растений вырождения, снижения жизненности и продуктивности. Дарвин всесторонне исследовал этот вопрос в труде «Действие перекрестного опыления и самоопыления в растительном мире». Но, каков механизм инбридинга, до развития менделевской генетики никто не знал. Ведь сказать, что инбридинг вреден, потому что снижает жизненность, — значит, лишь создать видимость объяснения. Генетика показала механизм инбридинга — переход в гомозиготное состояние рецессивных генов, многие из которых вредны, а то и летальны.
Вот хороший пример. В племенном животноводстве чрезвычайно высоко ценятся быки-производители. У прославленного быка голштинской породы Принца Адольфа к 1930 году было более двух тысяч сыновей, внуков и правнуков. Однако он имел рецессивный ген, носители которого в гомозиготном состоянии рождались без волосяного покрова и, естественно, погибали. Эти «лысые» телята принесли убытков на многие миллионы. Сейчас, когда развито искусственное осеменение и один и тот же бык способен фактически быть отцом всех телят в стране, подобный просчет может стоить много дороже.
Казалось бы, инбридинг в эволюции имеет лишь отрицательное значение. Однако это далеко не так. Вспомним мутацию фермента тирозиназы, вызывающего потемнение шерсти у животных. Есть рецессивные аллели, приводящие к образованию термолабильного — чувствительного к температуре фермента. При 37 °C темный пигмент — меланин в шерсти не образуется.
А теперь предположим, что какое-нибудь южное животное расселяется к северу, в места с суровыми зимами и снежным покровом. Мутанты по термолабильному ферменту на юге находились в гетерозиготном состоянии, а появляющиеся крайне редко светлые гомозиготы никаких преимуществ не имели. На границе ареала популяции малочисленны, и инбридинг приобретает тем большее значение, чем меньше популяция. Гомозигот становится все больше и больше; отраставшие у них летом, при высокой температуре, волосы оказываются слабо пигментированными. Такие светлоокрашенные особи, мало заметные зимой на фоне снежного покрова, получают преимущество в жизненной борьбе. Но под светлой шерстью, в условиях низкой температуры, у них вырастает темная, пигментированная (мутантный фермент начинает работать!) — и к началу лета, облиняв, они меняют окраску и снова становятся незаметными на темном фоне земли.
Вот так и возникала хорошо всем известная сезонная смена окраски меха (и оперения — у птиц), широко распространенная среди животных нашего климата. Мутационный процесс «поставил материал» — неустойчивый при высокой температуре фермент. Однако не будь изоляции, приведшей к инбридингу, и естественного отбора, оценившего результат, это несомненно ценное приспособление не смогло бы возникнуть, причем неоднократно, в разных группах неродственных организмов. Разумеется, все обстоит сложнее, чем мы изложили, окраска определяется совместным действием многих генов, но суть дела от этого не меняется.
В последнее время эффект инбридинга в малых популяциях стали даже переоценивать. Ход рассуждений здесь примерно такой: в каждом поколении число предков удваивается. Действительно, у каждого из нас один отец и одна мать, но два деда и две бабушки, четыре прадеда и четыре прабабушки и т. д. и т. д. Если принять, что человек как вид существует 500 тысяч лет (это очень умеренная цифра) и продолжительность поколения 25 лет, то число предков каждого человека должно выражаться сверхастрономической цифрой — 220 000. На Земле никогда не существовало столько людей. Поэтому мы должны поделиться друг с другом предками. Старая пословица: «среди четырех морей все люди братья» верна: все мы кровные родственники. Однако, экстраполируя родство, как говорится, к нулевой точке, нельзя впадать в абсурд и выводить человечество от единого предка — некоего обезьяньего Адама, возникшего в единственном экземпляре. Такой Адам не мог бы быть хранителем всех множественных аллелей (например, групп крови), роднящих нас с человекообразными обезьянами. Вспомните о «горлышке бутылки» — оно должно быть достаточно широким!
Популяция предкового вида, дающая начало новому, может быть небольшой, но это должна быть популяция, то есть группировка особей, а не герой-одиночка. В противном случае множественный аллелизм не мог бы унаследоваться при переходе от вида к виду.
Дрейф генов, или эволюция без отбора
Мы уже упоминали о том, что относительная численность аллелей в популяции абсолютно точно описывалась бы уравнением Харди — Вейнберга, если бы численность самой популяции была бесконечной. Чем меньше популяция, тем больше отклонения, и если численность размножающихся особей в ней окажется ниже 500, начинаются любопытные явления. Описавшие их советские генетики Н. П. Дубинин и Д. Д. Ромашов назвали их генетико-автоматическими процессами. В том же 1932 году к сходным выводам за рубежом пришел С. Райт, который применил более короткий термин — дрейф генов. Название, как отмечал Э. Майр, не очень удачное. Под дрейфом мы привыкли понимать пассивное движение объекта в одну сторону, например, смещение заряженной частицы в электрическом поле. Здесь же картина больше напоминает броуново движение. Однако термин прижился главным образом из-за лаконичности, что немаловажно в наш суматошный век.
Суть дрейфа генов в следующем. Допустим, популяция состоит из 100 особей, и отношение аллелей А и а (доминантный и рецессивный) 1:1. Казалось бы, в ней и в следующем поколении должно быть 25 АА, 50 Аа и 25 аа. Но из-за случайных колебаний один из аллелей может вообще выпасть из популяции, она окажется по данному гену гомозиготной. А какой из аллелей выпадет — доминантный или рецессивный — это уже дело чистого случая.
Дальнейшие выкладки приводят к еще более любопытным заключениям. Если концентрация аллеля в популяции мала, у него возрастает вероятность быть «выметенным» из популяции генетико-автоматическими процессами. Более многочисленный аллель успешнее противостоит дрейфу. Так реализуется библейская несправедливость — имущему дастся, от неимущего отнимется. Нетрудно сообразить, что это открывает путь случайной дифференцировке популяций — вплоть до видового обособления.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
5. Синтез рибосомных РНК
5. Синтез рибосомных РНК В обычных клетках синтез трех видов рРНК (28S, 18S и малой 5S) координирован, т. е. на одну молекулу 28S образуется одна молекула 18S и одна молекула 5S. Синтез 28S и 18S происходит в виде одного большого, общего для них предшественника (пре-рРНК), который затем
6. Гормоны регулируют синтез желтка и белка
6. Гормоны регулируют синтез желтка и белка Мы уже говорили, что у позвоночных животных желток будущего яйца синтезируется в печени. Этот синтез стимулируется женскими половыми стероидными гормонами — эстрогенами (подробнее см. специальную главу). Один из таких гормонов
Глава 7 Прыжок в пучины времени. Трудности в изучении ископаемых. «Пилтдаунский человек». Великий охотник – миф. Наши видовые отличия. Десмонд Моррис и его «безволосая обезьяна». Гипотеза Элистера Харди о водных приматах.
Глава 7 Прыжок в пучины времени. Трудности в изучении ископаемых. «Пилтдаунский человек». Великий охотник – миф. Наши видовые отличия. Десмонд Моррис и его «безволосая обезьяна». Гипотеза Элистера Харди о водных приматах. Погружаясь в пучины времени, не рассчитывайте на
Глава 3. Великий симбиоз
Глава 3. Великий симбиоз Эволюция «с человеческим лицом» Типичным примером примитивизации дарвиновских идей является широко распространенное по сей день мнение о том, что в основе эволюции лежат исключительно эгоизм, конкуренция и безжалостная «борьба за
2.4. Конфронтация или новый синтез?
2.4. Конфронтация или новый синтез? Наиболее оправданной позицией многим эволюционистам уже давно представлялся синтез положений СТЭ с концепциями направленной эволюции и сальтационизма на основе достижений генетики. Разные авторы говорили, что пора переходить от
«Великий эволюционный эксперимент» в африканских озерах
«Великий эволюционный эксперимент» в африканских озерах Рыбы цихлиды больших африканских озер — превосходный объект для изучения симпатрического видообразования. Во-первых, это пресноводная группа, а значит, они развиваются в относительно замкнутых (по сравнению с
3. ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ
3. ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ Описанная ранее концепция «светлого пятна» исходит из того, что сознание определяется некоторым уровнем возбудимости мозговых структур. Однако можно предположить, что этого недостаточно и в действительности
Синтез гликогена (гликогеногенез)
Синтез гликогена (гликогеногенез) Гликоген синтезируется в период пищеварения (через 1–2 часа после приема углеводный пищи). Синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т. е. требует затрат энергии.Синтез гликогена включает 4
Синтез жирных кислот
Синтез жирных кислот Синтез жирных кислот происходит в основном в печени, в меньшей степени – в жировой ткани и лактирующей молочной железе. Гликолиз и последующее окислительное декарбоксилирование пирувата способствуют увеличению концентрации ацетил-КоА в матриксе
5.5. Альтернативные теории и синтез идей эволюционизма
5.5. Альтернативные теории и синтез идей эволюционизма В рамках научной методологии эволюционизму нет альтернативы, поскольку такой альтернативой может служить только креационизм. Однако сам эволюционизм не является однородным течением. Хотя после чтения популярных
10.4. Достоевский — великий сострадалец и печальник
10.4. Достоевский — великий сострадалец и печальник По-видимому, в предыдущем разделе были достаточно подробно документированы жестокость и садизм Достоевского, связь этих свойств с личностным комплексом эпилептоидности-эпилепсии. Можно говорить о прямых уликах. Тему
Глава 6. Великий парламент инстинктов
Глава 6. Великий парламент инстинктов Как все в единство сплетено, Одно в другом воплощено! Гёте Как мы видели в предыдущей главе, эволюционный процесс ритуализации создает по мере надобности новый, автономный инстинкт, вступающий как независимая сила в общую систему
«Великий грабеж зерна»
«Великий грабеж зерна» Госсекретарь Генри Киссинджер провел внутреннюю интригу во властных коридорах, чтобы перехватить управление американской политикой сельского хозяйства, традиционно бывшей областью американского Министерства сельского хозяйства. Киссинджер
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация В определении понятия «гормон» было указано несколько этапов распространения гормонального сигнала (рис. 2.6). Рис. 2.6. Этапы распространения гормонального