Ген существует
Ген существует
…хромосомная теория наследств ценности вошла в золотой фонд науки человечества.
В. С. Немчинов
Три к одному
В шестидесятых годах прошлого века в августинском монастыре города Брюнна тогдашней Австрийской империи (ныне город Брно в Чехословакии) проживал странный монах Грегор (в миру — Иоганн) Мендель. Исполнением своих прямых монашеских обязанностей он явно тяготился, зато охотно преподавал в реальном училище, а в свободное время со страстью изучал природу во многих ее проявлениях. Он разводил пчел и выращивал растения в крохотном огороде (7?35 м) под окнами своей кельи, вел метеорологические наблюдения, переписывался со многими видными учеными Европы и живо интересовался последними новинками научной мысли, в том числе нашумевшей книгой некоего эсквайра Дарвина.
Удивляться нечему — рясу Мендель надел лишь для того, чтобы получить образование и верный кусок хлеба, не ведая, что монашеский сан через сто лет некоторые будут ставить ему в вину и даже отрицать на этом основании его безупречные опыты.
А опыты были воистину замечательными. Теперь они изучаются в школах и известны почти всем. Но на всякий случай напомним: Мендель установил, что признаки организмов (в данном случае — обычного гороха) можно разделить на две группы — доминантные и рецессивные[6]. Проявлялись они при скрещивании. Например, желтый цвет семядолей горошин подавлял зеленый цвет. Гибриды первого поколения все имели желтые горошины.
Однако Мендель на этом не остановился и высеял гибридные горошины снова. Странное дело: во втором поколении рецессивный признак — зеленый цвет горошин — вновь возник в среднем в 25 % случаев. На 152 824 желтых горошины оказалось 50 676 зеленых (отношение 3,004:0,996). Гениальность Менделя заключалась в том, что он истолковал полученный факт абсолютно правильно. Наследственные факторы организмов, как он называл гены, распределяются в потомстве случайно. Один ген организм получал от отца, другой — от матери.
Каковы же возможные сочетания генов (в данном случае — зеленых и желтых горошин)?
1. Зеленый + желтый.
2. Желтый + зеленый.
3. Желтый + желтый.
4. Зеленый + зеленый.
В любой из пар, кроме последней, доминантный фактор подавлял рецессивный, и отношение их в потомстве получалось 3:1. Совершенно аналогичный результат мы получили бы, подбрасывая сразу две монеты (договорившись, что выигрывает тот игрок, у которого хотя бы одна монета ляжет орлом кверху). В 25 % выпадает два орла, в 50 % — один орел и одна решка и лишь в 25 % — две решки. Эта аналогия имеет глубокий смысл — она указывает, что отношение 3:1 должно получиться лишь при достаточном числе бросков, то есть гибридных семян. Только в этом случае закономерность выступает из хаоса случайного.
Историки науки, после того как работа Менделя в 1900 году была переоткрыта, вспомнили, что подобные результаты получали многие исследователи, в том числе и сам Дарвин. Опишем для примера его опыт.
Дарвин скрестил обычный львиный зев с разновидностью, имевшей на верхушке радиально-симметричный, так называемый пелорический цветок. Потомство первого поколения оказалось все с обычными цветами, но во втором «из 127 проростков 88 имели цветы обычной формы, 2 — промежуточной между пелорической и нормальной и 37 — были пелорическими».
С учетом случайного отклонения отношение признаков было близко к 3:1. В этом нетрудно убедиться, подсчитав вероятную ошибку m по формуле
m = ± q(n — q)/n,
где n — сумма всех гибридов, a q — число, которое можно ожидать из соотношения 3:1. Почему же Дарвин не опередил Менделя?
Я не думаю, что причиной была пресловутая дарвиновская нелюбовь к математике. Дело гораздо сложнее. Предшественники Менделя пытались разгадать наследуемость всех признаков организма сразу, не абстрагируясь до судьбы отдельных. В результате первое поколение гибридов им казалось смешанным — ведь в нем проявлялись доминантные гены и со стороны отца, и со стороны матери. Отсюда прямая дорога к некоей слитной наследственности, о которой шла речь в предыдущей главе.
Второй, не менее важный закон Менделя касается гибридов, родители которых различаются по ряду признаков. Оказывается, эти признаки распределяются в потомстве независимо друг от друга, и комбинации их чисто случайны (в опыте Менделя, например, желтый и зеленый цвет горошин и морщинистая и гладкая их поверхность).
Законы Менделя далеко не всем могли понравиться — именно из-за четкого признания случайности в процессе перераспределения наследственных факторов. Поэтому после возрождения менделизма в 1900 году Де Фризом, Корренсом и Чермаком в биологии вспыхнули жаркие споры. Любопытная закономерность: самые непримиримые противники менделевских законов, если они были честными учеными, в конце концов становились их сторонниками и продолжателями дела Менделя.
Вот пример, малоизвестный и убедительный. Наш великий садовод И. В. Мичурин в 1915 году писал: «В последнее время наши неофиты дела гибридизации как-то особенно назойливо стараются нам навязать этот гороховый закон — создание австрийского монаха». Однако Мичурин был настоящим ученым и решил проверить законы Менделя на гибридах обычной яблони с яблоней Недзвецкого. Последний вид — очень удачный партнер для скрещивания: у него красные плоды, красная кора и даже хлорофилл листьев замаскирован красным антоциановым пигментом. Уже в 1917 году Мичурин описал первые подобные опыты, где были получены четкие числовые расщепления, а двадцать лет спустя отмечал: «В законе Менделя я нисколько не отвергаю его достоинств» — и рекомендовал для показательных скрещиваний, демонстрирующих применимость менделизма в генетике плодовых, именно яблоню Недзвецкого, а также краснолистную грушу, сливу и орех. Поэтому было бы кощунством утверждать, что Мичурин до конца дней отвергал менделизм. Все обстояло как раз наоборот.
Колоссально много для развития учения о наследственности сделал американец Томас Гент Морган, который вначале был столь же ярым противником Менделя и решил его опровергнуть на другом объекте — кроликах. Однако попечители Колумбийского университета, где работал Морган, сочли кроликов чересчур дорогими; Морган не пал духом и использовал для опытов крошечную плодовую мушку — дрозофилу. Дрозофила стала классическим объектом генетики — редчайший случай, когда скупость снабженцев оказала науке неоценимую услугу! Дело в том, что за год можно получить 25 поколений дрозофил и разместить на одном лабораторном столе их многотысячные «стада». Помимо этого, в работе с дрозофилой есть какая-то непонятная, но властная притягательность. Я помню, как мне в свое время пришлось поставить на ней несколько опытов — я без особой нужды для основной работы затянул их на добрые полгода и прервал с великим сожалением. Это, как говорят на лабораторном жаргоне, болтливый объект — так много из него познаешь нового.
Рассказать о всех открытиях Моргана и созданной им научной школы здесь невозможно: для этого потребовалась бы книга по меньшей мере такого же объема. Да в этом и нет нужды — в последнее время вышло достаточно хороших популярных книг, посвященных успехам генетики. Мы рассмотрим лишь те открытия, которые имеют первостепенное значение для эволюционной теории.
Морган и его не менее прославившиеся ученики — А. Стертевант, Г. Мёллер и К. Бриджес в 1909–1911 годах доказали, что второе правило Менделя не абсолютно. Полной независимости в распределении наследственных задатков нет: они распределены по группам сцепления, иными словами, — могут передаваться потомству лишь совместно. У дрозофилы четыре таких группы — столько же, сколько пар хромосом в клетках тела, и столько же, сколько хромосом получает мужская или женская половая клетка — гамета. Гениальное предвидение Августа Вейсмана, в XIX веке связавшего наследственность с хромосомами, полностью подтвердилось.
Далее, само правило сцепления оказалось неабсолютным. Гены могли переноситься из одной хромосомы в другую. Для тех, кто забыл школьную биологию или учился в период с 1948 по 1964 годы, напомним, что при делении клетки, приводящем к образованию гаметы (мейоз), хромосомы не делятся. Парные, гомологичные хромосомы выстраиваются друг против друга, а затем расходятся по клеткам. В результате яйцеклетка и спермий имеют уменьшенное вдвое число хромосом — у человека — 23, у дрозофилы — 4 (так называемый гаплоидный набор). Лишь слияние гамет, приводящее к возникновению оплодотворенной яйцеклетки — зиготы, восстанавливает прежний, диплоидный набор, характерный для вида.
Группа Моргана показала, что, выстроившись попарно, гомологичные хромосомы при мейозе могут обменяться частями. Это явление было названо кроссинговером (перекрестом). Оказалось, что цитологи уже давно наблюдали кроссинговер в клетках многих организмов.
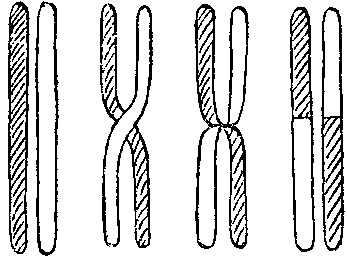
Схема кроссинговера.
Анализ кроссинговера показал, что гены в хромосомах расположены линейно в одномерной последовательности, как буквы в строке. Это позволило построить хромосомные карты генных локусов (локус — по-латыни просто место, в генетике — место, где расположен в хромосоме ген. Каждый локус может быть занят аллелем — формой какого-либо гена — например, аллелем желтой или зеленой окраски семядолей гороха). Чем дальше в хромосоме отстоят друг от друга локусы, тем больше вероятность того, что они расстанутся при кроссинговере.
Для эволюции кроссинговер имеет первостепенное значение. Возьмем для примера менделевский горох. Допустим, что ген желтой окраски горошин и ген, обуславливающий развитие длинных корней, находятся в одной хромосоме. Не будь кроссинговера, получение гороха с зелеными горошинами и длинными корнями было бы невозможным. Кроссинговер делает сцепление между генами неабсолютным: в результате его оба признака могут оказаться в одной хромосоме. Еще более важен для возникновения новых форм эффект неравного кроссинговера, открытый в 1925 году учеником Моргана Стертевантом. Стертевант изучал ген, вызывающий появление у дрозофилы полосковидных глаз, и установил, что возможны случаи не абсолютно точного обмена частями хромосом. После такого неравного кроссинговера в одной хромосоме оказывается два одинаковых локуса, а в другой — ни одного. Один и тот же локус, представленный двумя аллелями, становится двумя самостоятельными одноаллельными локусами. У многих организмов одинаковые гены многократно дублированы: в хромосоме имеется не один локус, а целая цепочка их. В таких случаях неравный кроссинговер — явление весьма частое: одна хромосома получает львиную долю локусов, другая — меньше их число. Идет как бы переселение генов из одной гомологичной хромосомы в другую. Однако возможен и обратный процесс выравнивания числа локусов: кроссинговер в этом случае восстанавливает равенство.
Первые успехи генетики позволили понять механизм комбинаторной, или комбинативной, изменчивости, имеющей огромное значение в эволюции. При мейозе отцовские и материнские хромосомы, образующие пары, расходятся по гаметам случайно. Каково же число возможных сочетаний пар хромосом? Как показывает математика, оно равняется 2 в степени, равной числу пар гомологичных хромосом. У кролика, например, оно равно 222 = 4 385 000, у дрозофилы 24 = 16. Остается еще раз восхвалить скупость попечителей Колумбийского университета. Используй Морган кроликов вместо дрозофилы, он мог бы просто утонуть в море генетических рекомбинаций, и группы сцепления были бы открыты позже.
Если мы учтем перетасовывающую деятельность кроссинговера, нам станет ясно: запасы комбинаторной изменчивости практически неисчерпаемы. Поэтому в природе не может быть двух размножающихся половым путем организмов с идентичными наборами генов, нет двух одинаковых особей. Исключение (подтверждающее правило!) — однояйцовые близнецы, черенки с одного куста, бактерии, полученные в потомстве от одной клетки.
Представим некий организм, у которого в хромосомах всего 1000 локусов (на деле даже у бактерий эта величина больше). Если каждый локус представлен серией из 10 аллелей, число возможных сочетании из них — 101000 (как указал английский генетик С. Райт, эта величина неизмеримо больше числа электронов во всей Вселенной). Разумеется, львиная доля подобных сочетаний была бы нежизнеспособной при любых изменениях среды. Но и того, что остается, более чем достаточно, чтобы служить «сырьем» для эволюционного процесса, поставляя отбору неисчерпаемый материал в течение практически бесконечного времени.
Гены влияют друг на друга
Генетические карты, построенные морганистами, выглядели на первый взгляд странно. Гены, определяющие развитие самых различных признаков, мирно уживались в хромосоме рядом, как бусинки на нитке. Могло сложиться впечатление (а у некоторых оно и сложилось), будто организм — не что иное, как мозаика признаков, каждый из которых независим от других и определяется одним геном.
Это наивное представление было быстро опровергнуто работами известного шведского генетика Г. Нильсон-Эле еще в 1908 году. Изучая гибриды между различными расами пшениц, Нильсон-Эле установил, что многие признаки, например окраска зерен, определяются не одним, а многими генами. В зависимости от сочетания таких полимерных генов признак может иметь разную интенсивность. Полимерия генов распространена чрезвычайно широко, что вполне объяснимо. Понятие «признак» весьма условно. Чем сложнее та особенность структуры, которую мы называем этим расплывчатым термином, тем большее количество генов ответственно за ее проявление.
Окраска каждого лепестка мака, например, контролируется по меньшей мере десятью генами. Не менее сложен генный контроль над окраской шерсти млекопитающих. Пигментация кожи у человека также полимерный признак, поэтому у супружеской пары — негра и белой женщины могут рождаться дети-мулаты с самыми разнообразными вариациями цвета кожи: от черной до почти белой.
Полимерия гена крайне важна в практическом отношении, потому что все хозяйственно значимые признаки, такие как яйценоскость кур, урожайность пшеницы, сахаристость свеклы, плодовитость и скороспелость животных и т. д., определяются полимерными генами.
Не менее существенное значение она имеет и для эволюции. Если признак определяется многими генами, он становится гораздо более стабильным, чем если бы определялся одним. Организм без полимерных генов был бы крайне неустойчивым; любая рекомбинация приводила бы к резкой изменчивости, что в большинстве случаев невыгодно. Эволюционировать такому виду было бы так же трудно, как нам написать картину, пользуясь только черной и белой красками, не смешивая их. Полимерия генов, определяющих количественные признаки, и дает всю гамму оттенков изменчивости. Скачки и метания взаимоисключающих одиночных аллелей заменяются плавными, постепенными переходами.
Важно для понимания процесса наследственности и явление плейотропии гена, то есть феномена, когда один и тот же ген определяет возникновение не одного, а многих признаков организма. Так, ген желтой окраски семядолей гороха определяет также окраску цветов и черешков листьев, что было подмечено еще Менделем. Дарвин также сообщал об удивительных случаях связи между, казалось бы, самыми разнородными признаками. По-видимому, генов, ответственных только за один наблюдаемый признак, нет.
В то же время совершенно неправильно считать, что плейотропия, как и полимерия, абсолютна (каждый ген определяет все признаки, и каждый признак определяется всеми генами). Такие виды вряд ли могли бы существовать и уж во всяком случае не смогли бы эволюционировать. Некоторая доля независимости в отношениях между признаками необходима, в противном случае вся система рушилась бы, как карточный домик, при изменении одного элемента. Поэтому в процессе эволюции складывается соотношение между плейотропией и полимерией генов, оптимальное для каждого вида.
Фенотип и генотип
Предтечей наступающей эры синтеза дарвинизма и менделевской генетики был датский исследователь В. Иоганнсен. Заслуги его перед генетикой огромны (кстати, он первый произнес слово «ген»). Как и Мендель, Иоганнсен не был ученым-профессионалом. В начале своей научной «карьеры» он был учеником аптекаря в датском городе Эльсиноре — городе Гамлета.
Иоганнсен изучал обыкновенную фасоль. Как правило, фасоль — строгий самоопылитель, поэтому от одной особи можно в больших количествах получить генетически однородное потомство — так называемую чистую линию. Иоганнсен вывел девятнадцать чистых линий фасоли, отличавшихся весом семян, и начал с ними экспериментировать.
Вес семян в каждой линии колебался вокруг некоторой средней величины, причем частота встречаемости разных вариантов описывалась колоколообразной кривой — так называемой кривой Гаусса, или просто гауссианой. Иоганнсен вел отбор в (пределах каждой линии на увеличение или уменьшение веса фасолин: его занимал вопрос — сместится ли в результате отбора среднее значение признака? Казалось бы, на этот вопрос сразу следовало дать положительный ответ: ведь земледельцы и животноводы испокон веков так делали, оставляя на племя наиболее ценные экземпляры, и почти всегда добивались успешных результатов. Достаточно сопоставить, например, вес и сахаристость корня дикой и домашней сахарной свеклы или яйценоскость банкивской куры и домашних леггорнов. Однако в науке не все очевидное достоверно. Отбор в пределах чистой линии сказался бессильным, вес семени по наследству не передавался. Если же вести отбор в свободно скрещивающейся популяции, происходит «…больший или меньший сдвиг — в направлении отбора — того среднего признака, вокруг которого, флуктуируя, варьируют соответствующие индивиды» (Иоганнсен). Вывод этот был чрезвычайно важен и полностью соответствовал дарвиновским представлениям о том, что отбор имеет дело с уже возникшей в популяции разнокачественностью особей. Все фасолины чистой линии были генетически однородны, так что отбирать было не из чего.
Но ведь по размеру и весу они были различными? Значит, генетическая однородность, «чистота крови», и внешняя разнородность мирно уживаются в одной чистой линии? Здесь мы переходим к важнейшему для современной биологии делению организма на генотип и фенотип.
Генотип — это набор генов организма, имеющаяся в нем генетическая программа развития. Фенотип — комплекс свойств и признаков организма, результат реализации генетической программы, это компромисс между требованиями генотипа и условиями внешней среды. Изменчивость внутри чистой линии фенотипическая, она ненаследственна. Внутрилинейные колебания веса фасолин столь же наследуются, как и вырезанный аппендикс, трудовые мозоли или умение играть на гитаре. Это не просто шутка: мы знаем, что способность к музыке наследуется (пример тому хотя бы семья Бахов, в течение ряда поколений давшая миру много известных и талантливых исполнителей и композиторов и одного гениального). Но даже Иоганну-Себастьяну приходилось учиться играть сначала.
Отрицание деления организма на генотип и фенотип неизбежно приводит к ламаркизму, так как при этом не проводится граница между наследственными изменениями генотипа и ненаследственными — фенотипа.
Подчеркнем, что генотип богаче фенотипа. Наследственные задатки организма в данных условиях среды каждый раз реализуются не полностью (хотя бы потому, что рецессивные аллели подавляются доминантными). К этому положению мы будем неоднократно возвращаться. Пока же отметим, что наличие в генотипе организма какого-нибудь гена отнюдь не свидетельствует, что определяемый им признак непременно возникнет в фенотипе.
Советский генетик Н. В. Тимофеев-Ресовский предложил оценивать фенотипическое проявление гена двумя параметрами. Их называют пенетрантностью и экспрессивностью. Пенетрантность — это процент особей — носителей данного гена, у которых он проявился в фенотипе, а экспрессивность — степень выражения этого признака. Например, у дрозофилы описан ген vestigial, изменяющий форму и размер крыльев. Действие его сильно зависит от температуры. Мушки с одинаковым генным набором, но выросшие при 24° и при 30°, имеют совершенно разные крылья. И экспрессивность и пенетрантность поддерживаются естественным отбором и отражают реакции сходных генотипов на среду.
Одна из разновидностей китайской примулы, если ее выращивать в оранжерее при температуре 30°, имеет белые цветы, а выращенная при 20°, она зацветет красными цветами. Есть такая порода кроликов — гималайские. При 30° шерсть у них совершенно белая. Если же они растут при 25°, на наиболее охлаждающихся частях тела — ушах, лапах, кончике носа — вырастает черная шерсть. Такая же шерсть отрастает на любом участке кожи, если прежнюю сбрить и на бритое место прикладывать, например, пузырь со льдом.

Два поросенка из одного помета, выращенные в разных условиях. Различия между ними свидетельствуют о том, что задатки, заложенные в генотипе, могут проявиться лишь в конкретных условиях.
У новозеландских овец известен рецессивный ген, вызывающий у гомозигот (то есть особей, унаследовавших его от отца и от матери) болезнь печени. Печень при этом наследственном заболевании не может перерабатывать продукты распада хлорофилла, в результате они, накапливаясь в коже, повышают ее чувствительность к солнечным лучам. Итог печальный — ягненок слепнет, а затем гибнет от экземы и воспаления кожи. Однако при желании такого ягненка можно спасти — нужно только не держать его на солнце или же не кормить зеленой травой. Ген тогда никак не проявится, и обнаружить его будет невозможно. Среда как бы включает один ген, подавляя другой. Все явления подобного рода именуются наследственной нормой реакции (то есть реакции гена на среду). Это понятие весьма важно в практическом отношении. Какими бы отличными генами ни обладал, например, поросенок, при плохом кормлении из него хорошей свиньи не получишь. Самый прославленный сорт пшеницы не дает высокого урожая без удобрений. Но зато в благоприятных условиях лучшие задатки генотипа развернутся в фенотипе полностью. А низкопродуктивные сорта и породы, какие бы райские условия им ни создавали, ничего хорошего не принесут.
Это, казалось бы, банальные истины. Увы, многие еще полагают, что дело не в наследственности, а в заботливом содержании. Такое смешение генотипа с фенотипом столь же наивно, как и вера в то, что, если дворняжку кормить до отвала мясом, она вырастет до размеров дога. При развитии организма среда действует лишь как проявитель — при любых концентрациях метола и гидрохинона незасвеченный негатив останется прозрачным.
Генетики после Иоганнсена хорошо усвоили разницу между геном и признаком. Не лишне отметить, что еще Мендель не ставил знака равенства между признаком и фактором, его определяющим. Забвение этой истины, периодически наступающее у некоторых исследователей, неизбежно приводило к возрождению ламаркизма в самых худших, вульгарных вариантах.
Поэтому надо различать генетику — науку о наследственности и феногенетику — науку о способах и путях, которыми наследственность реализуется в признаках организма. Один из животрепещущих вопросов феногенетики мы рассмотрим в следующем разделе.
Как слуга становится господином
Эта проблема весьма важна для эволюционной теории, но до конца еще не разработана. Почему, в силу каких особенностей одни гены доминантны, а другие рецессивны?
Пока ясно одно: доминантность — не абсолютное, а относительное свойство аллеля. Напомним, что само понятие «доминантный» («доминус» — господин) говорит о том, что доминантным ген может быть лишь по отношению к другому гену. В свою очередь для третьего гена он может сам оказаться рецессивным. Степень доминантности может быть различной, иногда доминантность бывает неполная — в проявлении признака участвуют порой оба гена (тогда их называют кодоминантными). Например, хорошо известное садовое растение ночная красавица имеет две формы — красноцветковую и белоцветковую. Цветы гибридов розовые, но во втором поколении вновь происходит расщепление на белую и красную форму. Один и тот же признак может наследоваться как доминантно, так и рецессивно. Так складка во внутреннем углу глаза (эпикантус) у монгольской расы — признак доминантный, а у южноафриканских бушменов и готтентотов — рецессивный. Быть может, это свидетельствует о том, что эпикантус в том и в другом случае возник независимо. Но возможно и другое объяснение — эволюция доминантности в том и другом случае происходила по-разному.
Что же такое эволюция доминантности?
Вернемся более чем на 80 лет назад, когда И. В. Мичурин скрестил владимирскую вишню (дающую розовые плоды) с белоплодной черешней сорта Винклер белая. Гибридный сеянец дал белые плоды «с едва заметным розовым оттенком на световом бочку», — иными словами, белоплодность доминировала. Трудно сказать, как пошло бы дело дальше, оставайся гибрид на собственных корнях. Ведь и мы в детстве часто бываем светловолосыми, а потом нередко значительно темнеем. Маточное дерево гибрида, названного Красой Севера, замерзло в суровую зиму, но почки от него были привиты на обычную красную вишню. Результатом было появление розовоплодных гибридов. Заметьте, что это было все еще первое поколение гибридов, но в результате прививки розово- и белоплодность обменялись доминантностью. Так возник мичуринский метод ментора (Ментор — наставник сына Одиссея), основа которого заключалась в управлении доминантностью.
Обычно в таких случаях гибриды первого поколения прививались на растение-ментор. Подбор ментора — в высшей степени сложное дело, во многом зависящее от интуиции; если ментор оказывался удачным, пластические вещества подвоя, поступающие в привой, подавляли в последнем проявление нежелательных доминантных генов, и в фенотипе проявлялись рецессивные. После того как все качества привоя проявлялись и сорт оказывался пригодным, он размножался вегетативно.
Подчеркнем, что метод ментора ничего общего не имеет с так называемой «вегетативной гибридизацией» — здесь нет передачи признаков подвоя на привой, а лишь управление доминантностью генов привоя.
Ментор — не единственный и отнюдь не главный способ, при помощи которого Мичурин управлял доминантностью гибридов первого поколения. Главными способами были внешние условия — температура и влажность, свет и почва. Мичурин понимал, что возникновение доминантности — сложный и длительный исторический процесс, в котором первостепенную роль играет искусственный и естественный отбор, — недаром он указывал, что в гибридах доминируют прежде всего признаки диких видов («дичков»), затем старых, устоявшихся культурных сортов и, наконец, — молодых.
Уже в 30-х годах нашего века эволюция доминантности была переоткрыта английским генетиком Р. Фишером и другими учеными на иных объектах. Было выдвинуто сразу несколько гипотез, объяснявших механизм этого явления. Не вдаваясь в подробности, скажем лишь, что ни одна из них, взятая в отдельности, не объясняет эволюцию доминантности до конца. Вероятнее всего, в природе существует несколько механизмов, управляющих проявлением гена в признаке. Отбор, как и полагал Мичурин, играет в этом важную роль, что показали опыты английского генетика Э. Форда на бабочке — смородинной пяденице. Эта бабочка в норме имеет крылья с пятнами на белом фоне. Известна, однако, мутация с желтым фоном крыльев. У гетерозигот (гибридных форм с обеими аллелями в геноме) цвет фона крыльев промежуточный. Форд разделил экспериментальную популяцию на две части: в одной отбирались гетерозиготы наиболее светлые, а в другой — наиболее желтые. Через несколько поколений в первой части ген «желтого фона» стал полностью рецессивным, а во второй — полностью доминантным.
Значение подобных опытов станет ясным, если мы вспомним, что возникающие заново аллели обычно бывают рецессивными (признаки «дичков» доминируют над культурными). Жесткий отбор на выживание рецессивов должен привести — и приводит — к тому, что они становятся доминантными.
Начинает постепенно проясняться вопрос о том, что за вещества «включают» или «выключают» тот или иной аллель. А. Кюн и его сотрудники работали с гусеницами мелкой бабочки — амбарной огневки, а Г. Бидл и Б. Эфрусси — с личинками дрозофилы. Они осуществляли прививки на животном материале, пересаживая из одной гусеницы в другую скопления клеток — имагинальные диски, из которых развиваются части тела взрослого насекомого. Удалось выделить вещество, названное геногормоном, которое вызывало четкие изменения окраски глаз. Впоследствии геногормоны — производные каротиноидов — были получены для многих насекомых. Действие их сравнительно малоспецифично: и у мухи-дроэофилы и у бабочки-огневки, например, цвет глаз управляется одним геногормоном. Когда мы узнаем во всех тонкостях, какие вещества управляют доминантностью генов, мы сможем подавлять действие нежелательных генов у гибридов с небывалой до сего точностью и в буквальном смысле слова лепить новые растительные и животные формы…
Итак, классическая генетика, развивавшаяся бурными темпами с начала XX столетия, заложила прочный фундамент изучения одного из трех «китов» эволюции по Дарвину — наследственности. Параллельно шло интенсивное исследование антитезы наследственности — изменчивости.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Существует ли летняя спячка в мире животных?
Существует ли летняя спячка в мире животных? Интересное биологическое явление, вызываемое периодическими (или неожиданными) метеорологическими переменами, изменяющимися условиями жизни в летний сезон, представляет собой так называемая летняя спячка у животных,
ИСКОПАЕМЫХ ЧЕЛОВЕЧЕСКИХ КОСТЕЙ НЕ СУЩЕСТВУЕТ
ИСКОПАЕМЫХ ЧЕЛОВЕЧЕСКИХ КОСТЕЙ НЕ СУЩЕСТВУЕТ Известно, что человеческих костей среди ископаемых еще не найдено; это лишнее доказательство, что ископаемые расы не были вариациями, так как они не могли подвергнуться влиянию человека.Я говорю, что человеческих костей
Существует ли «демократия» в мире животных?
Существует ли «демократия» в мире животных? Наряду со строгой иерархией (и даже «диктатурой») в сообществах животных существует также «демократия». Установлено, что совместному решению большинства животные подчиняются охотнее, чем единоличному решению лидера.
СУЩЕСТВУЕТ ЛИ ОСОБЫЙ ОРГАН ЧУВСТВ, ПРИ ПОМОЩИ КОТОРОГО ГОЛУБИ ОПРЕДЕЛЯЮТ НАПРАВЛЕНИЕ?
СУЩЕСТВУЕТ ЛИ ОСОБЫЙ ОРГАН ЧУВСТВ, ПРИ ПОМОЩИ КОТОРОГО ГОЛУБИ ОПРЕДЕЛЯЮТ НАПРАВЛЕНИЕ? С каждым годом становится все очевиднее, насколько сложно найти объяснение навигационным способностям птиц, основываясь на общепринятых научных представлениях, — ив наше время эта
Бессонницы не существует
Бессонницы не существует Наша книга приближается к концу. Мы с вами познакомились с историей изучения сна, гипотезами и открытиями в этой области. Мы также рассмотрели почти все виды нарушений сна и бодрствования. Вместе с тем наше замечание насчет рождения новой
Глава 8 ВАВИЛОНСКОЕ СТОЛПОТВОРЕНИЕ Почему существует так много языков, как и откуда они появились
Глава 8 ВАВИЛОНСКОЕ СТОЛПОТВОРЕНИЕ Почему существует так много языков, как и откуда они появились На всей земле был один язык и одно наречие. Двинувшись с востока, они нашли в земле Сеннаар равнину и поселились там. И сказали друг другу: наделаем кирпичей и обожжем огнем. И