Ген изменяется
Ген изменяется
Изменение не наследственное для нас не существенно.
Ч. Дарвин

Вначале был термин
С первых лет нашего века, в пору возникновения классической генетики, исследователей интересовали вопросы: насколько стабильны гены? Какими факторами можно их изменять и можно ли это делать направленно? Иными словами, речь шла о разгадке тайны изменчивости — второго фактора эволюции из указанных Дарвином.
Остановимся на крайней точке зрения, впервые высказанной еще А. Вейсманом и вслед за ним Дж. Лотси. Согласно этой концепции, изменчивость определяется в основном генетическими рекомбинациями — перетасовками весьма стабильных, практически неизменяемых наследственных задатков.
Сторонников этой концепции у нас порой огулом зачисляли в идеалисты и реакционеры, причем обвинения были скорее эмоциональными, чем убедительными; никто из них не считал ген сверхстабильным образованием, наподобие кусочка платины (ведь тогда уже было известно о распаде самих атомов!). Дело в другом: первые генетики часто полагали, что условия, при которых ген изменяется, настолько жесткие, что не могут быть совместимы с жизнью. Поэтому эволюция использует старые гены в новых комбинациях, а астрономические цифры возможных комбинаций позволяют надеяться, что в ближайшие миллиард миллиардов лет процесс эволюции не кончится.
Парадоксально, но сторонники стабильности гена были не так уж неправы, хотя ген оказался изменяющимся.
Да, Вейсман прав: эволюция гораздо чаще использует старые гены, чем создает новые. Исключение составляют лишь микроорганизмы с гаплоидным, то есть одинарным, набором генов. И все-таки новые аллели возникают непрерывно. Стабильность гена отнюдь не абсолютна.
В этом можно убедиться, наблюдая возникновение форм с наследственно измененными признаками. Такие изменения издавна называли мутациями.
Сейчас уже трудно установить, кто был автором этого термина, возникшего в XVII–XVIII веках. То ли гейдельбергский аптекарь Шпренгер, описавший появление странного растения чистотела с разрезными листьями, то ли французский садовод Дюшен, нашедший землянику с простыми, а не тройчатыми листьями, то ли ботаник М. Адансон, описавший ряд изменений у льнянки, ячменя, пролески. Впрочем, латынь тогда была у ученых в большой чести (ботаники и по сие время дают описания — диагнозы новых видов на языке Вергилия и Цицерона). А мутация (mutation) по-латыни и будет просто — изменение, так что первые ученые, употреблявшие это слово, по-видимому, и не собирались придумывать новый термин, а лишь констатировали факт появления новых признаков. Воскресил слово «мутация» и наполнил его новым содержанием голландский ученый Гуго Де Фриз, один из переоткрывателей менделизма.
Началось с того, что Де Фриз задался целью проверить Дарвина — воспроизвести процесс видообразования в эксперименте. Сначала он высевал семена, собранные с растений больных, уродливых, выросших в неблагоприятных условиях существования, но не добился успеха. Лишь в 1886 году он открыл объект, позволивший сделать важные выводы.
Есть такое растение — ламаркова энотера, или ослинник. Это близкий родственник всем известного иван-чая из семейства кипрейных, завезенный из Америки сорняк (хотя окультуренную крупноцветную форму его иногда разводят в палисадниках). Я не видел энотеры до недавнего времени, но встретив ее в одичавшем парке под Киевом, мгновенно узнал по многочисленным изображениям. Еще бы — трудно найти растение, из-за которого в научных спорах было бы сломано больше копий! Ведь из-за него пытались опровергать дарвинизм.

Энотера (она же ночная свечка и ослинник) — самое знаменитое растение в генетике.
Проходя мимо заросшего энотерой картофельного поля близ деревни Гилверсум, Де Фриз заметил, что в массу обычных растений вкраплены резко отличающиеся от них экземпляры. Собрав семена, он посеял их в опытном саду и делал это на протяжении семнадцати лет, исследовав более пятнадцати тысяч растений! Вначале были обнаружены три резко отличающиеся формы; потом Де Фриз стал описывать их десятками. Многие из них были весьма стабильными. Мутация изменяла не один признак, а все растение и стойко наследовалась без расщепления: комплекс признаков переходил по наследству как единое целое.
К каким же выводам пришел Де Фриз? Прежде всего, он заключил, что Дарвин неправ. Видообразование — не постепенный процесс приспособления к условиям внешней среды, а скачкообразное порождение одним видом другого. Иными словами, в процессе эволюции скачком изменяются не гены, а сами виды. Новый вид является из недр другого, как Афина-Паллада из головы Зевса, вооруженный против всех превратностей внешней среды. Роль естественного отбора при этом сводится к уничтожению неудачных мутантов.
Де Фриз не знал, что до него сходные идеи развивал талантливый русский ботаник и убежденный антидарвинист С. И. Коржинский. Собрав много фактов спонтанной (внезапной, скачкообразной) изменчивости, Коржинский построил теорию гетерогенезиса (рождения разнообразия), во всех деталях аналогичную мутационной теории Де Фриза. Разумеется, оба — и Де Фриз и Коржинский — были неправы. Нет таких мутаций, которые бы сразу порождали вид, а отрицание роли отбора в возникновении целесообразности сразу отбрасывает нас к Ламарку и Фоме Аквинскому. И все же основные положения Де Фриза оказались совершенно правильными:
1. Мутации возникают скачкообразно, без переходов.
2. Раз возникшая мутация устойчива, в отличие от фенотипических изменений.
3. Мутации возникают ненаправленно; одна и та же мутация может возникать повторно.
Эти выводы Де Фриза положили начало исследованиям изменчивости генотипов; ошибки же были исправлены временем. Сам термин «мутация» оказался чересчур широким, описывающим целую группу явлений. Теперь генетики различают в наследственной изменчивости следующие категории:
1. Геномные мутации, когда изменяется число хромосом в наборе.
2. Хромосомные перестройки, в процессе которых или изменяется последовательность генов внутри одной хромосомы, или негомологичные хромосомы обмениваются частями. В ту же категорию относят случаи, когда кусок хромосомы удваивается или, наоборот, теряется.
3. Точковые, или генные, мутации, изменяющие структуру отдельных генов и невидимые в микроскоп.
О каждой из этих категорий и роли их в эволюции следует поговорить подробнее.
Число хромосом — паспорт вида
Итак, в настоящее время мутациями называют изменения свойств и признаков организма, вызванные изменением структуры генома — совокупности хромосом клетки. Эти изменения иногда можно наблюдать в клетке в оптический микроскоп. В первую очередь наблюдаются так называемые геномные мутации, при которых умножается набор хромосом, свойственный виду. Такое явление называют автополиплоидией.
Число хромосом, на которые разделяется генный материал ядра при делении клеток, весьма варьирует у разных видов — от 2 хромосом у лошадиной аскариды до 1260 у тропического папоротника ужовника из Индии. Есть данные, что у одноклеточных организмов — радиолярий число хромосом достигает 1600. Дальнейшее умножение генного материала приводит к потере митоза и возникновению амитоза — простого деления ядра без скрупулезно точного механизма расхождения хромосом.
Наиболее широко полиплоидия представлена у растений. Она встречается у низших растений — водорослей, у грибов. Хорошо изучен процесс полиплоидизации у высших, цветковых растений. Многие роды и виды растений представляют полиплоидные ряды с числом хромосом n[7], 2n, 3n, 4n — до 308n у некоторых форм шелковицы и 265n — у мятлика.
С полиплоидными рядами связан один парадокс эволюции. Так, если гаплоидное число хромосом у растения n, а диплоидное, соответственно, 2n, то тетраплоидная форма (4n), возникнув в течение одного поколения, сразу поведет себя как новый вид, генетически изолированный от материнской формы. Это вполне понятно: при скрещивании образуется стерильный триплоид, так как одна гамета имеет хромосомный набор n, вторая 2n, а оплодотворенная яйцеклетка 3n. Три, как известно, на два не делится. Поэтому процесс редукционного деления ядра у триплоидов (мейоз, см. стр. 19) приведет к неравномерному расхождению генного материала по гаметам и бесплодию. Многие специалисты так и полагают, что в данном случае скачком возникает новый вид. Вряд ли это правильно — генетическая информация у полиплоидных форм та же, изменяется только ее количество. Генетическая изоляция, неспособность скрещиваться, — мы будем это неоднократно подчеркивать — необходимый, но еще недостаточный критерий вида.
Однако фенотип полиплоидов нередко изменяется. Полиплоиды — часто крупные, а то и гигантские формы с увеличенными ядрами и клетками. Жизнеспособность и продуктивность их может быть гораздо выше, чем у обычных диплоидов, что нередко используется в селекции культурных растений. Известны высокопродуктивные полиплоиды картофеля и хлопчатника, льна и гречихи, пшеницы и овса, земляники и сахарного тростника — список этот можно многократно увеличить. Главный поставщик натурального каучука — гевея у себя на родине, в Бразилии, имела 36 хромосом (здесь и далее без оговорок приводятся числа для диплоидных наборов — 2n). В юго-восточной Азии культивируются 72-хромосомные формы гевеи.
Даже не образующие семян триплоиды высоко ценятся в хозяйстве — упомянем только гигантскую триплоидную осину, культурный бессемянный банан и ряд других форм, размножающихся вегетативно.
Было бы, однако, ошибкой утверждать, что полиплоидия всегда связана с увеличением мощности растения. По-видимому, есть некий оптимальный уровень полиплоидности, разный у разных видов, за которым начинается как бы разлад взаимоотношений ядра и цитоплазмы и, как следствие этого, — карликовость, снижение жизнеспособности и продуктивности.
В эволюции растений полиплоидия проявляется двояко — как тормоз и как стимулятор прогресса. Многие древние растения (такие как древовидные папоротники, магнолия, гигантская секвойя или необычайный ботанический монстр из пустыни Калахари — вельвичия удивительная) — полиплоиды. В данном случае избыточность генетической информации в полиплоидном наборе тормозит эволюцию. В других случаях полиплоиды более изменчивы, легче приспосабливаются к новым условиям, нередко заселяют места на границе ареала с крайними условиями существования. Больше всего полиплоидов в Арктике, в высокогорных районах и пустынях (на Шпицбергене, например, 80 % видов — полиплоиды).
Не менее широко распространена у растений аллополиплоидия — удвоение генного материала у межвидовых или межродовых гибридов. Обычно такие гибриды бесплодны, так как хромосомы одного вида не находят себе парных хромосом-гомологов при мейозе. Иное дело, если хромосомный набор гибрида удваивается (возникает амфидиплоид). Здесь нельзя не вспомнить пионерские работы замечательного советского генетика Г. Д. Карпеченко, получившего впервые межродовой гибрид капусты и редьки. Такие гибриды обычно не давали потомства, потому что образовывали гаметы с нарушенным числом хромосом. Полученный Карпеченко гибридный тетраплоид был вполне плодовитым и не скрещивался ни с кем из родителей — ни с редькой, ни с капустой. Формально он заслуживал выделения в новый род — редькокапуста — рафанобрассика.
По проложенному Карпеченко пути устремилось немало исследователей. При этом не только создавались новые виды (вернее, гибриды, могущие стать новыми видами после «пришлифовки» отбором к условиям внешней среды), но и ресинтезировались старые. Из скрещивания алычи с терном удалось, например, заново сконструировать сливу. Таким же путем был ресинтезирован табак и рапс, пикульник и разные виды пшениц.
Возникают ли в настоящее время подобные гибридные формы в природе?
Такие случаи были описаны. На юге Якутии, по опушкам хвойных лесов на протяжении более 300 км обнаружена популяция оригинального кустарника — рябинокизильника, возникшего в результате естественного скрещивания сибирской рябины с кизильником. Это уже, пожалуй, настоящий гибридный вид, захвативший немалую территорию и успешно отвоевывающий себе место под солнцем. Сходным путем в США возник гибридный вид из двух видов диких подсолнечников.
А встречается ли авто- и аллоплоидия в царстве животных, и если да, то какое значение она имеет для их эволюции? На этот вопрос ответить нелегко.
Возникновение полиплоидов у раздельнополых животных (как и двудомных растений) затрудняется существованием хромосомного механизма определения пола.
Вспомним, что, например, у человека 46 хромосом. Из них 22 пары (их называют аутосомами) одинаковы у обоих полов. Но, помимо 44 аутосом, женщины имеют 2 одинаковые х-хромосомы, а мужчины — 1х и 1у хромосому. В половых клетках число хромосом уменьшается вдвое. Нетрудно сообразить, что яйцеклетки всегда будут иметь, помимо 22 аутосом, одну х-хромосому, а спермии — или х- или (y-хромосому. Спермий, несущий х-хромосому, сливаясь с яйцеклеткой, всегда дает зиготу женского пола (восстанавливается набор хх). Спермий с у-хромосомой, напротив, обусловит мужской пол (набор ху). Представим теперь полиплоид с набором ххуу. Такой набор приводит к возникновению гамет: 25 % хх, 50 % ху и 25 % уу. Зиготы же будут иметь набор 25 % хххх, 50 % ххху и 25 % ххуу. Половина потомства, таким образом, окажется аномальной в половом отношении, почти наверняка бесплодной. Возникновение таких организмов приведет к снижению плодовитости полиплоида и вытеснению его с жизненной арены.
Этого мало: ведь даже для того, чтобы описанный процесс произошел, нужно, чтобы оба родителя оказались одинаковыми полиплоидами. Вероятность такого события — величина чудовищно маленькая. Скрещивание же, например, тетраплоида с обычной, диплоидной, особью другого пола приводит к образованию триплоидов, генетически стерильных. Полиплоид, как и вообще любой организм с достаточно существенной хромосомной перестройкой, оказывается в положении библейского Каина. Если верить библии, у первых людей на Земле — Адама и Евы, было два сына — Каин и Авель. Каин убил Авеля, чем поставил под угрозу дальнейшее существование рода человеческого, но взял себе в жены «женщину из страны Нод», предки которой остались неизвестными, и род человеческий продолжился. Парадокс Каина неизбежно встает на пути всех теоретиков, которые, не удовлетворяясь дарвиновской теорией, ищут материал для эволюции в крупных, революционных перестройках генома. На ком женился Каин? Где найдет пару полиплоид?
Разумеется, хромосомный механизм определения пола может существенно отличаться в деталях от описанного нами. Половая хромосома может быть лишь одна (тогда самки имеют набор хх, а самцы хо); у птиц и ряда насекомых одинаковые половые хромосомы присущи самцам, а не самкам; наконец, половые хромосомы могут быть не самостоятельными, а прикрепляться к аутосомам. Однако во всех этих случаях объяснение редкости полиплоидии у животных, данное выдающимся американским генетиком Германом Меллером, остается в силе.
Наиболее убедительное его доказательство — это то, что у гермафродитных животных (например, дождевых червей) и у тех видов и рас двуполых животных, которые размножаются партеногенетически, без оплодотворения яйцеклетки, существуют полиплоидные ряды (n, 2n, Зn и т. д.), во всем аналогичные полиплоидным рядам растений.
Небольшой рачок артемия, обитающий в водах соленых озер, часто теряет способность к двуполому размножению, и у него отмечены партеногенетические полиплоидные расы. Такие же расы обнаружены у многих насекомых — жуков-долгоносиков, бабочек, прямокрылых, тлей.
Еще более интересны случаи так называемого гиногенеза — оригинальной модификации партеногенеза. Уже давно было подмечено, что в популяциях серебряного карася во многих наших прудах самцы встречаются крайне редко, а то и совсем отсутствуют. Оказалось, что икра карасей из таких однополых популяций развивается партеногенетически, без оплодотворения, но развитие икринки должно быть активировано спермием самца другого вида (вьюна, карпа). Спермий, дав толчок развитию, рассасывается в цитоплазме икринки. В потомстве, естественно, возникают одни самки. Однополая форма карася оказалась триплоидной, с тремя наборами хромосом (у двуполой формы 2n = 94, у однополой 141).
Караси-амазонки, исследованные советскими генетиками К. А. Головинской, Д. Д. Ромашовым и Н. Б. Черфас, не представляют собой исключения. Такой механизм известен у живородящей рыбки молинезии из мелких водоемов Центральной Америки. Гиногенетичными оказались многие насекомые, некоторые черви и амфибии. Часть описанных случаев отличается поразительной сложностью.
Один из видов американской саламандры — амбистома латерале (близкий вид — тигровая хорошо известен любителям живой природы под названием аксолотля) в одних и тех же водоемах может быть представлен диплоидной и триплоидной расой. Триплоидные амазонки, более крупные и сильные, чем обычная диплоидная форма, могли бы ее вытеснить и вымереть вслед за ней сами, так как их икринки стимулируются к развитию диплоидными самцами. Однако баланс между формами поддерживается тем, что у триплоидов плодовитость значительно меньше, да и развитие икринки затягивается на более длительный срок.
В последнее время снова возник спор о роли полиплоидизации в эволюции животных, особенно тех, у которых пол не однозначно определяется хромосомным набором, а зависит от соотношения в организме женских и мужских половых гормонов. Особенно стремятся доказать ее широкую распространенность в природе сторонники так называемого «мгновенного видообразования», когда в течение одного поколения возникает новый вид, не смешивающийся с материнской формой. Но на пути подобных построений неизбежно встает парадокс Каина. Растения обходят его, так как способны к самоопылению и вегетативному размножению. Но как быть с животными?
И все же есть факты, свидетельствующие о возможном удвоении генома у двуполых животных. Рыжий таракан-пруссак имеет 24 хромосомы, а крупный черный — 48. У карпа 104 хромосомы, а у большинства других карповых — 52. Лососевые рыбы, как это установлено достаточно твердо, — тетраплоиды. Однако многие подобные случаи оказались проявлением так называемой ложной полиплоидии, обусловленной расщеплением и слиянием хромосом и рядом других факторов. В опыте полиплоидия животных возможна. Непрямым путем — через партеногенез и межвидовую гибридизацию Б. Л. Астаурову и его сотрудникам удалось сконструировать аллотетраплоид — гибрид домашнего и дикого тутового шелкопряда. Я умышленно говорю — сконструировать, потому что, когда читаешь описание этого замечательного опыта, именно это слово приходит на ум. Аллотетраплоид Астаурова, как и рафанобрассика Карпеченко — форма, генетически изолированная от обоих родительских видов. Для того чтобы ее получить, потребовалось:
1) научиться получать партеногенетическое потомство с диплоидным набором хромосом (здесь индуктором развития яйцеклетки выступает не сперма другого вида, а нагрев до 48 °C в течение 18 минут);
2) получить тетраплоидных самок, размножающихся партеногенетически (число хромосом 4n = 112);
3) скрещиванием тетраплоидных самок с обычными диплоидными самцами получить триплоидные формы шелкопряда;
4) из триплоидных форм получить гексаплоидные, с шестью хромосомными наборами;
5) гексаплоидных самок домашнего шелкопряда скрестить с диплоидными самцами дикого. Потомство будет иметь соответственно — три набора хромосом от домашнего и один — от дикого шелкопряда, то есть будет аллотетраплоидным;
6) путем отбора из поколения в поколение повысить плодовитость гибридов.
Значение подобных опытов трудно переоценить, ибо они закладывают фундамент новой отрасли селекции и генетики — получению плодовитых гибридов между разными видами животных. Однако вряд ли виды часто возникали в природе подобным путем, так как арсенал средств природы несравненно беднее, чем у современного экспериментатора. У природы другой козырь — время, в течение которого самые радикальные перестройки генома могут стать в принципе не невозможными…
Теперь остановимся коротко на менее решительных перестройках генома — частичном удвоении числа хромосом, соединении двух хромосом в одну или, наоборот, разделении их. Такие случаи встречаются гораздо чаще, чем авто- и аллополиплоидия.
Долгое время среди генетиков бытовало убеждение, что число хромосом — абсолютно стабильный видовой показатель. Однако оно оказалось не совсем верным. Описаны случаи, когда число хромосом изменяется или путем распада одной хромосомы на две, или путем слияния двух негомологичных (не образующих пары при мейозе) в одну. Возможна и полисомия — удвоение не всего хромосомного набора, а лишь части хромосом. Такие межхромосомные перестройки весьма важны для эволюционного процесса. Увеличение числа хромосом приводит к резкому увеличению комбинаторной изменчивости. Наоборот, слияние их снижает изменчивость, следствием чего является замедление образования форм, стабилизация вида.
В последние годы широко распространено мнение, что первичные, примитивные виды имеют геном, состоящий из многих мелких хромосом, а эволюционный процесс идет путем слияния их в крупные. Возможно, во многих случаях это так — и все-таки мне кажется, что здесь уместно вспомнить гетевское — «теория мертва, лишь древо жизни вечно зеленеет». Эволюцию нельзя загонять в узкие рамки одной тенденции. Есть прямые свидетельства того, что изменения генома шли в разных направлениях, многократно комбинируясь. Примером могут служить олени. До наших дней сохранились мелкие примитивные тропические оленьки — мунтжаки; в той же группе есть и значительно эволюционно подвинутые виды. Оказывается, у большинства видов оленей 50–70 хромосом, у примитивного оленя Давида, как у лани и благородного оленя, 2n = 62, у одного из видов мунтжаков (мунтжак Ривса) — 46, а у другого — только 6, как у комара, наименьшее известное для млекопитающих хромосомное число! Комментарии, как говорится, излишни.
Хромосомные мутации
Менее заметны при цитологическом анализе, но довольно часто встречаются такие хромосомные перестройки, как инверсии и транслокации. В случае инверсии участок хромосомы перевертывается на 180°; такая перестройка затрагивает только одну хромосому. При транслокации происходит обмен участками между двумя негомологичными хромосомами (отличие от кроссинговера!).
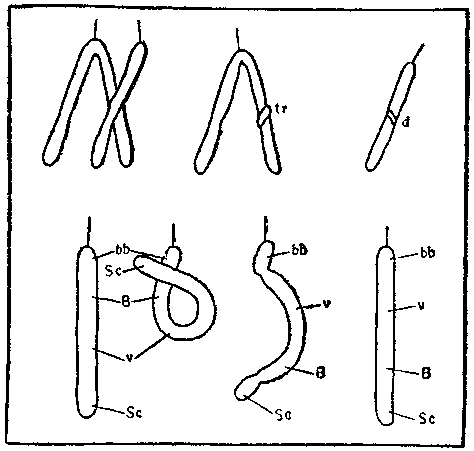
Единая схема возникновения мутации (по А. С. Серебровскому, 1929). Верхний ряд — образование транслокаций (tr) и нехваток (d); нижний ряд — образование инверсии; Sc, v, В, bb — символы разных генов.
Замечательный советский генетик Александр Сергеевич Серебровский еще в 1929 году расшифровал механизм возникновения инверсий и транслокаций — разрыв хромосомы с последующим воссоединением в другом месте. Если хромосома разорвется сразу в двух точках, оторванный участок может, перевернувшись на 180°, воссоединиться с остальным телом хромосомы. Разорванные концы хромосом, как оказалось, «слипаются» с высокой вероятностью (молекулярный механизм этого явления мы начинаем угадывать только сейчас).
Но если разрыв произошел в одном месте, «поворота кругом» не происходит. Конец хромосомы, именуемый теломером, воссоединяться даже со свежим разрывом не может, в данном случае возможно лишь восстановление прежней структуры. Если же она не восстанавливается, оторванный кусок бесследно рассасывается, и генетическая информация, заключенная в нем, пропадает. Такие аварии называют концевыми нехватками или дефишенси. Судьба организма, геном которого потерпел дефишенси, складывается по-разному. Если потерянный кусок мал и не содержит жизненно важных генов, организм, особенно полиплоидный, это легко перенесет. Потеря большого куска хромосомы может быть смертельной. Например, потеря значительной части одной хромосомы в геноме человека приводит обычно к смертельной генетической болезни, называемой «синдромом мяукания» (cri de chat — по звукам, издаваемым больными младенцами).
В случае двойного разрыва хромосомы оторванный кусок также может рассосаться в плазме (воссоединяется только концевой кусок с теломером). Такие нехватки называют делециями. Описано много случаев делеций у самых различных организмов. Крупные делеции, как правило, в гомозиготном состоянии летальны, мелкие — вызывают самые разнообразные изменения признаков фенотипа.
Казалось бы, инверсии гораздо безобиднее для организма, ибо генетическая информация при них не теряется, а лишь меняется ее расположение (делецию можно сравнить со случаем, когда рассеянный киномеханик забывает показать одну из частей фильма, а инверсию — когда он их перепутывает). Однако из собственного опыта вы знаете, что зрители кричат «сапожники» в обоих случаях. Так и с инверсиями. Они часто в гомозиготном состоянии летальны, то есть, будучи унаследованными от отца и матери, приводят к смерти организма. Такие инверсии обнаруживаются только у гетерозигот, где хотя бы одна хромосома из пары не инвертирована.
Даже сравнительно безобидные инверсии ограничивают комбинаторную изменчивость. Как говорят генетики, они «запирают кроссинговер». Это и понятно: ведь кроссинговер — обмен гомологичными частями хромосом. Если геном не гомозиготен по инверсии, хромосомы обменяются негомологичными частями, и у половых клеток будет двойной набор одних генов и нехватка других, а потому они окажутся нежизнеспособными.
Эволюционная роль инверсий, как и прочих хромосомных перестроек, именно в ограничении рекомбинаций. В результате удачная комбинация генов оказывается изолированной и размноженной. Исследуя наборы хромосом разных видов, цитогенетики могут порой установить, каким путем возникла генетическая изоляция, нескрещиваемость, — путем слияний хромосом или же в результате одной или нескольких инверсий.
Второй тип хромосомных перестроек — транслокации, при которых негомологичные хромосомы обмениваются частями, имеет меньшее значение для эволюции, на что обратил внимание еще Серебровский. Причина ясна: даже если транслокация в гомозиготном состоянии вполне жизнеспособна, гетерозиготы образуют нежизнеспособные гаметы с нехватками генного материала. Размножаемость особей, гетерозиготных по этой мутации, резко падает (до 50 %), и естественный отбор отметает такие формы. А так как вероятность встречи двух животных с одинаковой транслокацией ничтожна, то и случаи «использования» подобных мутаций в эволюционном процессе можно буквально сосчитать по пальцам. Лишь у растений с их способностью к самоопылению и вегетативному размножению транслокация может закрепиться в популяции и дать начало форме, не скрещивающейся с родительской. Такие формы описаны, например, у энотеры, пиона, дурмана. Их гибриды с другими формами стерильны. Некоторые из этих форм-транслокантов энотеры и сбили с толку Де Фриза, решившего, что он наблюдает воочию процесс видообразования. Оказалось, что у энотеры часто все хромосомы транслоцируются одна на другую, образуя при мейозе замкнутое кольцо. В результате возникает множество самоопыляемых рас, нерасщепляющихся и изолированных друг от друга. Формально это, конечно, виды… если признать достаточным критерием вида генетическую обособленность.
Удивительна сложность транслокаций у многих растений. Один из видов энотеры — энотера биеннис имеет два разных хромосомных кольца, получивших специальные названия — рубенс и альбиканс. При мейозе кольца расходятся, но жизнеспособна лишь пыльца с набором рубенс, а женские половые клетки — лишь с набором альбиканс. Оплодотворение восстанавливает двойной набор; естественно, 50 % мужских и женских гамет пропадает зря. Из поколения в поколение гетерозиготность восстанавливается такой сравнительно дорогой ценой; возможность возникновения гомозигот рубенс-рубенс и альбиканс-альбиканс пресечена с самого начала.
У животных дело иное — способны к размножению лишь некоторые транслоканты, у которых половая формула самки хх, а самца — хо. Если аутосома присоединится, как говорят, транслоцируется на половую хромосому, то становится ее частью и возникает нео-у-хромосома. Транслокации между аутосомами у животных, как правило, заканчиваются для потомства фатально. Лишь немногие виды, например, скорпионов смогли приобрести комплексы транслоцированных хромосом, аналогичные тем, что мы наблюдаем у пионов и энотеры.
Это и натолкнуло Серебровского на оригинальную идею нового метода борьбы с вредными насекомыми, который он назвал генетическим.
Генетический метод по-гениальному прост. Нужно вывести в культуре линию насекомых, подлежащих уничтожению, причем линию гомозиготную по транслокации. Если мы выпустим в природу массу самцов транслоцированной линии, они будут скрещиваться с дикими самками; дальнейшее размножение гетерозиготного потомства будет подавлено, так как не менее 40–50 % эмбрионов отмирают уже на ранних стадиях. Серебровский четко указал условия, повышающие эффективность метода:
1) самцов-транслокантов надо выпускать в самом начале периода размножения, когда численность популяции вредителя невелика;
2) наибольший эффект дадут линии, гомозиготные не по одной, а по нескольким транслокациям. Их легче получить для видов, геном которых состоит из многих хромосом. Если же геномы вредителей малохромосомны (как у мух и комаров), можно выпускать в природу сразу несколько транслоцированных линий.
Не ограничившись теорией, Серебровский приступил к выведению линий-транслокантов комнатной мухи и амбарного долгоносика. Нам сейчас, пожалуй, трудно оценить всю смелость подобного шага — во все времена вредителей полагалось уничтожать, а не разводить, чтобы потом выпустить в природу. Однако ни смелости, ни находчивости Серебровскому не нужно было занимать. Ведь не кто иной, как он вывез в 1927 году из Германии новую породу кроликов способом, который не пришел бы в голову самому искушенному контрабандисту. В то время высоко (до 200 марок за самку с приплодом) ценилась порода кроликов реке с «плюшевым» мехом. Желая сэкономить государству валютные средства, Серебровский привез из Германии пару метисов, имевших самый беспородный вид и ничего не стоивших. Но эти метисы были гетерозиготами — носителями рецессивной мутации реке. Уже в первом поколении выщепилось 2 рекса, в третьем эта мутация была уже размножена и пошла в производство.
К сожалению, разработка генетического метода с 1948 года была прервана; к тому же в эти годы шло повальное увлечение ДДТ и прочими инсектицидами. Химические методы борьбы казались самыми простыми и дешевыми; однако они уничтожали все живое чересчур прямолинейно, хочется сказать — по-скалозубовски («Уж коли зло пресечь…»). После того, как было установлено, что на одного вредителя, уничтоженного инсектицидами, приходится десять полезных насекомых, а ДДТ обнаружили не только в молоке кормящих матерей, но и в яйцах антарктических пингвинов, о генетическом методе вспомнили. Многих энтомологов-практиков пленила его абсолютная безопасность и четкая направленность действия. В самом деле: он касается только объекта, против которого разработан. За рубежом быстро перевели статью Серебровского, и она стала настольной для целой армии исследователей. Быстрому распространению метода помогли и новые источники мутагенного излучения — ядерные реакторы и поставляемые ими изотопы. Выращенных в лаборатории самцов-вредителей облучали дозой, вызывающей множественные хромосомные мутации в ядрах спермиев, а затем рассеивали с самолетов. Результаты были великолепными.
Таким способом в 1954 году уничтожили муху, откладывающую яички в раны домашнего скота, сначала на острове Кюрасао в Карибском море, а потом и в ряде североамериканских штатов. С тех пор список побед генетического метода многократно умножился, и сам он значительно усовершенствован. Стерильных самцов, например, обрабатывают половыми запаховыми веществами (феромонами), делающими их неотразимыми для диких самок. Другой путь — выведение в лаборатории рас, имеющих в геноме доминантные гены с летальным эффектом. Для хлопкового долгоносика получена, к примеру, раса с геном, блокирующим период зимнего покоя (диапаузу). В лаборатории она, разумеется, может разводиться бесконечно. Но гибриды ее с дикой расой в природе осенью вымерзают.
Вся история генетического метода — наглядное доказательство того, что наиболее отвлеченные, самые теоретические изыскания лучше служат практике, чем деляческий практицизм. Ведь все началось с того, что Серебровский в 1929 году задумался: почему инверсии в эволюции закрепляются неизмеримо чаще транслокаций? Одни выгоды от генетического метода борьбы с вредителями стократно окупают все расходы по изучению той же эволюции. К тому же мы попутно перестаем отравлять природу и самих себя инсектицидами, а впереди открываются еще более заманчивые перспективы направленной перестройки природы.
Точковые мутации — насущный хлеб эволюции
А теперь поговорим о точковых, или генных, мутациях, изменяющих каким-то образом структуру одного гена. Они не видны в световой микроскоп (подавляющая часть их неразличима пока даже в электронном микроскопе). Однако они, пожалуй, играют важнейшую роль в эволюции, поставляя для нее львиную долю материала. Все рассмотренные нами до них крупные, революционные перестройки генома на деле оказываются «реакционными». Удвоение хромосомного набора, транслокация, довольно крупная инверсия — все они ограничивают скрещиваемость, ограждая изменившийся организм от других членов популяции «китайской стеной», практически во многих случаях непреодолимой (парадокс Каина!). Даже безобидная инверсия резко ограничивает рекомбинацию, запирая кроссинговер. Точковые мутации не препятствуют рекомбинации, поэтому они легко перераспределяются, тасуясь, как карты.
Механизм генных мутаций до развития молекулярной биологии оставался неизвестным. Говорили, что при них ген изменяет структуру, что в общем-то было тавтологией (напомним, что мутация и означает изменение). Однако незнание механизма отнюдь не мешало постижению этого фактора изменчивости.
Удалось установить, что влияние точковой мутации на развитие фенотипа может быть разным — от едва заметного до смертельного, летального. Организм, гомозиготный по летальному гену, погибает на какой-нибудь из стадий развития. Известно также, что разные гены (локусы гомологичных хромосом) различаются по изменчивости. Одни из них мутируют чаще, другие реже. Возникновение многих мутантов дрозофил и других форм животных и растений наблюдали многократно. Возможны и обратные мутации, когда мутантный ген возвращается к прежнему состоянию, а фенотип — к дикому типу. Это явление называют реверсией. Если реверсии не наблюдается, то скорее всего произошла потеря гена — та же делеция, но в масштабе, недоступном световому микроскопу.
Иногда реверсия бывает ложной. Она имитируется в фенотипе прямой мутацией другого гена (так называемые супрессорные мутации). Любопытно, что порознь обе такие мутации могут быть летальными или же резко снижать жизнеспособность гомозиготного по ним организма. Собравшись в геноме вместе, они не проявляются в фенотипе — восстанавливается дикий тип. Минус на минус дает плюс. Это напоминает двух героев народной сказки, один из которых лишился ног, а другой зрения. Объединив усилия, они разыскали источник живой воды, исцелили свои увечья и восторжествовали над врагами. Эта сказочная ситуация часто реализуется в природе.
Еще более любопытен другой факт. До сих пор мы говорили о генах, представленных не более чем двумя формами — аллелями (например, желтый и зеленый цвет горошин в опытах Менделя). Аллель — слово мужского рода, хотя часто сбиваются и употребляют его в женском. Если вы окажетесь в таком затруднении, вспомните сходное имя (пастушок Лель из «Снегурочки» Островского). Так вот, один и тот же ген может иметь не два, а больше — до нескольких десятков и даже сотен аллельных состояний, изменяясь по-разному. Это явление называется множественным аллелизмом. Все вы, вероятно, видели кроликов различной окраски — черных, белых с черными ушами и лапами и полностью белых с красными глазами (альбиносов). Это все — аллели одного гена.
Другой, более серьезный пример — мутантные аллели гена, отвечающего у человека за одну из реакций с гипоксантином и гуанином. Практически все аллели приводят в гомозиготном состоянии к накоплению в организме мочевой кислоты, но уровень развития болезни разный. Один аллель вызывает лишь подагру, другой — почечнокаменную болезнь. В полной форме еще в детском возрасте возникает тяжелейшее нервное заболевание с параличами и характерными припадками, во время которых больной кусает сам себя (синдром Леша-Нихана). Более известный пример, также из генетики человека, — это так называемые группы крови. Они обусловлены множественными аллелями, из которых шире всего распространены три (так называемая система АВ0, читается А-Б-ноль).
Как мы уже упоминали, стабильность гена хотя и не абсолютна (как думал Вейсман), но весьма высока. У человека в среднем частота мутирования отдельных генов 1:100 000; впрочем, разные авторы называют разные цифры. Но число генов в ядрах клеток настолько велико, что не менее 10 % яйцеклеток и спермиев несет какую-либо вновь возникшую мутацию. Каждый из нас — носитель доброй тысячи мутаций. Почему же изменчивость в природе все же ограничена? Ведь все виды достаточно стабильны и сохраняют, как часто говорят, «дикий тип». Все дело в гетерозиготности: большинство мутантных аллелей рецессивно (вспомните эволюцию доминантности!). Не проявляясь в фенотипе, мутировавший ген может переходить сотни поколений от предков к потомкам, и, лишь встретившись с таким же геном в гомологичной хромосоме, будет обнаружен. Эти процессы имеют важнейшее значение для эволюции, и о них мы будем еще много говорить.
Закон гомологических рядов, или параллельная изменчивость
У читателя может сложиться впечатление, что если «запасы изменчивости», накопленные в генофондах и непрерывно пополняемые мутационным процессом, столь велики, то в принципе возможно получение организмов с любым набором признаков.
Однако это далеко не так. Цветоводы с незапамятных времен выводили розы с цветами самой невероятной окраски. Тем не менее, никто не видел синюю розу. Нормальный цвет глаз плодовой мушки — дрозофилы — красновато-коричневый, но генетики вывели мутантные линии с глазами белыми, киноварными, ярко-красными и бурыми. А вот дрозофилу с голубыми или зелеными глазами никто не смог получить — и вряд ли это вообще возможно. Есть признаки для того или иного вида как бы запретные.
И наоборот, уже давно было подмечено, что если какой-либо признак встречается у одной из форм вида, весьма вероятно, что он обнаружится у вида близкородственного. Фигурально выражаясь, ген нельзя уподобить бильярдному шару, который случайно может обратиться к вам любой точкой своей поверхности. Скорее, это монета, которая падает или орлом, или решкой, а в случае множественного аллелизма — игральная кость, выпадающая лишь какой-либо из своих шести граней.
Такие случаи знал уже Дарвин: «…одинаковые признаки время от времени проявляются у некоторых разновидностей или рас, ведущих начало от одного и того же вида и, реже, в потомстве отдаленных видов». В подтверждение этого он ссылался на французского ботаника Ш. Нодэна, обнаружившего параллельную изменчивость у тыквенных, и английского энтомолога Б. Уолша. Впоследствии много подобных фактов обнаружили ботаники (М. Дюваль-Жув и уже известный нам Де Фриз) и палеонтологи (Э. Коп и Г. Осборн). Однако не хватало исследователя с синтетическим складом ума, способного охватить умственным взором (как писал М. В. Ломоносов — «умными очами») все это громадное скопище разрозненных фактов и построить общую теорию.
А теперь перенесемся в 1920 год. Казалось бы, до науки ли тут — но в Саратове собирается III Всероссийский селекционный съезд. После доклада молодого, но уже пользующегося заслуженной известностью генетика Н. И. Вавилова аудитория разражается громом рукоплесканий. И на вопрос — что происходит? — посторонний свидетель слышит ответ — «это биологи приветствуют своего Менделеева».
Сравнение весьма лестное, образное, но обязывающее. Что же сделал Вавилов? Можно ли его труд сравнить с периодической системой Д. И. Менделеева? Попробуем проследить ход мыслей Вавилова, путь, по которому он прошел, когда делал свое открытие.
В начале XX века систематика растений (да и животных) чудовищно осложнилась по сравнению с той, что дал отец систематики Карл Линней в XVIII веке. Линней знал одну пшеницу, одну рожь, один рис. Но линнеевские виды — линнеоны — оказались отнюдь не однородными единицами. Их удалось разделить на сотни и тысячи наследственных форм, причем процесс дробления представлялся явно бесконечным. Дикие, неокультуренные человеком виды не являлись исключением. А. Жордан, например, обыкновенную невзрачную травку — крупку разбил более чем на 200 видов! Такие мини-виды стали в отличие от линнеонов называть жорданонами. Систематики уже захлебывались в океане видов, которые описывали сами.
Итак, исходная точка Вавилова — полиморфизм крупных видов — линнеонов. Оттолкнувшись от нее, он подмечает параллелизм изменчивости линнеонов. Пшеницы, например, распадаются на три группы видов — с 42, 28 и 14 хромосомами. В каждой из этих групп имеются сходные формы, или возникшие параллельно, или унаследованные от общего предка: остистые и безостые, бело-, красно- и черноколосые, озимые и яровые. Такие же ряды Вавилов находит у ячменей и ржи, овсов и проса, хлопчатника и пырея, огурцов, дынь и тыкв.
Этого мало. Параллельная изменчивость обнаружилась и у представителей различных родов. Так, если у пшениц имеются формы с опушенными колосьями, то у ржи они также должны иметься. И действительно, Вавилов находит их сначала на Памире, а потом в Армении. Сходство гомологичных[8] форм из разных родов настолько велико, что их порой принимали за межродовые гибриды. Например, С. И. Коржинский за 23 года до доклада Вавилова описал гибриды арбузов и дынь — гибриды ложные, никогда не существовавшие. Вавилов нашел сорта арбузов, похожие не только на дыни, но и на тыквы и кабачки.
Я вспомнил этот пример Вавилова, когда мы как-то достали в туземной деревне на острове Молекула (архипелаг Новые Гебриды) загадочные плоды, формой и размером семечек похожие на огурцы, но по вкусу — типичные арбузы.
Насколько же далеко идет гомология изменчивости? Оказывается, ее можно проследить между весьма отдаленными семействами и даже классами. Однако эти примеры, естественно, касаются лишь самых общих черт строения. Так, альбинизм (возникновение бесхлорофильных форм), гигантизм и карликовость, превращение корней в корнеплоды, сходные форма плодов и окраска цветов возникают в семействах, связанных друг с другом чрезвычайно отдаленным родством. Свекла, например, родственница лебеды и дерева пустынь — саксаула (семейство маревых), но она образует корнеплоды, удивительно похожие на корнеплоды моркови (семейство зонтичных) или редьки (семейство крестоцветных).
Вавилов отметил также, что процессы параллельной изменчивости наблюдались не только у высших растений, но и у водорослей и грибов. Гомологические ряды изменчивости установлены у таких животных, как инфузория и паразитические черви, моллюски и морские пауки — пантоподы, земноводные, черепахи, ящерицы, птицы и млекопитающие. Например, известный генетик Дж. Холдэйн изучил параллелизм окраски шерсти у грызунов и хищных. Оказалось, что у мышей, крыс, кроликов и морских свинок, по-видимому, есть идентичные гены окраски, хотя кроликов сейчас вообще выделяют из грызунов в самостоятельный отряд зайцеобразных, а морская свинка — родственница, как ни странно, не крысе, а дикобразу.
Дальнейшая судьба закона гомологических рядов, или гомологической изменчивости, оказалась весьма оригинальной.
Генетики и селекционеры в большинстве восторженно приветствовали идеи Вавилова и охотно применяли их в своей работе. Подобно тому, как химики искали новые элементы по карте — периодической системе Менделеева, селекционеры искали и находили нужные им формы, исходя из закона гомологии. Бобовое растение люпин, часто произрастающее в подмосковных палисадниках, кроме красивых цветов, дает большой выход зеленой массы и отлично обогащает почву азотом. Казалось бы, это прекрасная кормовая культура. Однако люпины не поедаются скотом из-за горького вкуса. Руководствуясь трудами Вавилова, селекционеры из ГДР довольно быстро нашли негорькие формы люпинов, которые и стали родоначальниками новых сортов.
Отношение же к закону Вавилова теоретиков порой было скептическим. Речь идет не о тех, кто отказывал Вавилову в праве на приоритет, вспоминая всех его предшественников. (Отдавать предшественникам должное, разумеется, необходимо, но здесь не следует перегибать палки, в противном случае творцом закона гомологических рядов следует признать первого человека, заметившего, что кроме белых и черных собак существуют белые и черные кошки). Гораздо опаснее были те, кто признавал параллелизм в изменчивости, но делал из этого антидарвиновские выводы.
Ход рассуждений был при этом следующим. Согласно Дарвину, изменчивость неопределенна. Однако сами факты гомологической изменчивости якобы свидетельствуют, что Дарвин ошибался: изменчивость направленна, она имеет цель. В результате сложилось парадоксальное положение — в конце концов трудно поверить, что творец закона гомологических рядов был ограниченнее (если не сказать — глупее) легиона последующих комментаторов и, оставаясь на стороне дарвинизма, не догадывался, что он одним из самых своих значительных открытий опровергает Дарвина.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Как изменяется температура с погружением в глубь Земли?
Как изменяется температура с погружением в глубь Земли? Земной шар покрыт каменной оболочкой – это земная кора. При углублении на каждые 33 метра внутрь земной коры температура повышается в среднем на 1 градус. Этот темп повышения температуры с глубиной (геотермический
Изменяется ли длина носа взрослого человека с возрастом?
Изменяется ли длина носа взрослого человека с возрастом? Измерив длину носа у 2500 человек, швейцарские медики пришли к выводу, что с возрастом она увеличивается. У 97-летнего мужчины нос в среднем на 0,8 сантиметра длиннее, чем у 30-летнего. В 30 лет средняя длина носа у