Естественный отбор и феногеография
Естественный отбор и феногеография
Изучение естественного отбора — одна из важнейших задач в изучении микроэволюции. Без глубокого понимания действия этого единственного направленного эволюционного фактора ни о каком переходе к управляемой эволюции не может быть и речи. Точных же примеров, вскрывающих действие отбора, известно пока чрезвычайно мало — число их не превышает нескольких десятков.
Феногеография дает в руки исследователя удобное орудие для изучения отбора. Достаточно напомнить вошедший в школьные учебники пример с распространением меланистической формы березовой пяденицы в Англии — явление индустриального меланизма, вскрытое и проанализированное прежде всего на феногеографической основе. Другим классическим примером действия отбора, в котором феногеография сыграла не последнее место, следует считать случай с расшифровкой поддержания высокого уровня заболеваемости серповидно-клеточной анемией в некоторых районах Африки, — как раз в тех районах, где распространена малярия. Ген серповидно-клеточности смертелен в гомозиготном состоянии и, казалось бы, должен стать редким в популяции. Однако в гетерозиготном состоянии этот ген определяет устойчивость к заболеванию малярией — другому страшному бичу многих районов Африки. Отбор поддерживает определенную частоту смертоносного гена серповидно-клеточности в популяциях потому, что тем самым уменьшатся потери популяций от другой смертельной болезни. И серповидно-клеточность, и устойчивость к малярии можно рассматривать как своеобразные физиологические фены.
Приведу некоторые новые примеры выявления действия отбора с помощью феногеографии. Изучение наземной улитки Сереа vindobonensis в горных долинах Югославии, проведенное в конце 60-х годов английским генетиком Дж. Джонсоном, показало необычное распределение фенов бесполосных и полосатых раковин. Оказалось, что фен бесполосости встречается на хорошо прогреваемых склонах, тогда как улитки с темными раковинами (которые, как показали эксперименты, лучше выживают в холоде и способны лучше утилизировать солнечную энергию) занимают самые холодные склоны (рис. 17).
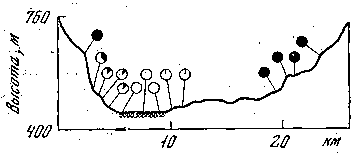
Рис. 17. Распределение бесполосных и полосатых раковин наземного моллюска в одной из долин Югославии
Черный цвет — встречаемость особей с темными раковинами, обитающих на самых холодных склонах
На другом виде этого же рода — улитке Сереа nemoralis были проведены феногеографические исследования, вскрывшие действие отбора. Когда концентрацию желтых раковин нанесли на карту Европы, оказалось, что существует прямая зависимость между частотой желтых форм и средней летней температурой. В экспериментах было выявлено, что желтые морфы по сравнению с более темными обладают пониженной выживаемостью в холодных условиях.
Из всех хорошо проанализированных работ по изучению связи географической изменчивости с действием отбора следует одно общее заключение: везде, где обнаруживается действие естественного отбора, существуют четкие клинальные изменения по типичным фенам — дискретным признакам.
Нутрия, обитающая на п-ове Флорида, обнаруживает четкую клинальную изменчивость по палевой и темной окраске: процент палевых особей в популяциях очень высок на севере района и очень мал на юге. Непосредственная причина такого изменения окраски пока неизвестна, но можно быть уверенным, что главный механизм клинальной изменчивости — направленное действие отбора, связанное с какими-то изменениями градиентов условий существования. Еще более крутую клину[21] описал М. Магомедмирзаев по изменению доли красных и коричневых семян в соцветиях солнцецвета иволистного на расстоянии 250 км по западному побережью Каспийского моря. Эколого-физиологический анализ показал связь коричневосеменной формы с большей ксерофильностью — приспособленностью к более сухим условиям обитания, что, видимо, и определило картину современного распространения этих двух фенов на данном участке ареала.
Два других примера касаются птиц Северной Атлантики. Большой поморник обитает практически по всем удобным местам побережий Северного, Норвежского, Гренландского и Баренцева морей. У него встречаются два цветовых фена — темная и палевая окраски. Процент палевых птиц очень мал на юге ареала и достигает 100 % на севере. Исследование показало, что палевая форма быстрее достигает половой зрелости: обладает повышенной агрессивностью. (Это важно для успешного питания, так как значительную часть своей добычи птицы отнимают у мелких морских чаек, что определило даже латинское название вида — parasiticus.) Однако в спаривании темные самцы имеют больше успеха. Наконец, темные самцы более жизнеспособны в условиях умеренного климата, а палевые в условиях арктических морей. Сложное сочетание всех этих преимуществ и определяет возникновение типичного сбалансированного полиморфизма — ни одна из форм не имеет решающего преимущества перед другой на большей части ареала (кроме Гренландии, где полностью отсутствует темная форма).
Другим примером служит распространение фена «очковости» у тонкоклювой кайры (Uria aagle). Этот фен выражается в появлении темного кольца вокруг глаз, соединенного через клюв тонкой темной полоской. В самой южной колонии вида — на побережье Португалии — «очковые» кайры не встречаются, но к северу число таких птиц увеличивается и достигает максимума в субарктических районах. Сравнительно мало известно о направлениях отбора, поддерживающих такую клинальную изменчивость. Вскрытие этих конкретных направлений отбора — дело физиологов, экологов, этологов. Феногеография в данном случае выступает в виде компаса, стрелка которого показывает на те ситуации в природе, в которых ясно прослеживается влияние отбора.
Последним из серии примеров, показывающих возможности феногеографии для выявления в природных популяциях процессов естественного отбора, может послужить замечательный пример клинальной изменчивости моли (Amates glareosa) на Шетлендских островах, расположенных у северного побережья Англии.
Изучение этой ночной бабочки было проведено английским генетиком Г. Кеттлевелом, который в 50-х годах успешно исследовал индустриальный меланизм насекомых, в частности знаменитую березовую пяденицу. У моли встречаются две формы — серая и темная, меланистическая. На архипелаге, протянувшемся на 136 км, на севере существуют только меланистические популяции, на юге — почти полностью свободные от меланистов. Нанесенные на график изменения частоты меланистов показали резкий перерыв клинальной изменчивости в одном из районов главного острова — там, где проходит небольшая долина. Здесь на протяжении 13 км концентрация меланистической формы изменяется на 35 % (что соответствует в среднем 2,7 % на каждый километр), по всему остальному архипелагу она меняется в среднем в пять раз медленнее. Так феногеография указала на самое интересное место для дальнейших исследований. Такие исследования были организованы и проведены в окрестностях долины с вересковыми пустошами и редкими полями на известняковых склонах.
На этом можно и заключить описание этого случая, поскольку дальше для расшифровки природной ситуации потребовались не феногеографические, а обстоятельные эколого-генетические исследования. На обеих сторонах долины шириной всего в 1–3 км выпускали меченых бабочек и определяли, как они разлетаются от мест выпуска. Оказалось, что отдельные особи часто улетают на расстояние свыше полутора километров. Всего здесь было помечено и выпущено 1682 бабочки, из них поймано — 65. Несмотря на то что постоянные ветры во время эксперимента дули так, что должны были бы переносить бабочек с одной стороны долины на другую, была найдена только одна бабочка, перелетевшая на противоположную сторону долины.
Эксперимент показал, что долина является реальной преградой в передвижении бабочек. До сих пор непонятно, в чем тут дело. Непонятно и адаптивное значение потемнения бабочек на севере архипелага. Есть несколько предположений, в частности, о преимуществе покровительственной окраски темных форм в условиях более длительного северного светового дня. Об этом говорит и тот факт, что в желудках мелких чаек на севере архипелага было найдено 20,7 % светлых форм, тогда как в местной природной популяции светлых форм было всего 2,7 %. К этому надо добавить, что обнаружилась разница в продолжительности существования в природе особей местного происхождения и привезенных из других мест и выпущенных в данном месте. Так, например, по данным одного эксперимента длительность жизни местных бабочек составила 3,4 дня, а «привозные» — как темные, так и светлые формы — жили почти на день меньше. Известная английская пословица «East or West, home is best» оправдывается здесь как нельзя лучше!
Исследования этой бабочки показали, что клинальная изменчивость обеспечивается не равномерным потоком мигрантов при наличии градиента природных условий, а каждая клина представляет собой множество мелких, частично изолированных популяционных островков с изменяющимися в зависимости от микроусловий микронаправлениями естественного отбора. На больших пространствах микронаправления могут складываться в заметные макрофеногеографические изменения.
В целом феногеография дает широкие возможности для изучения естественного отбора — этого во многом еще таинственного ключевого фактора эволюционного процесса. Главная роль, которую играет при этом феногеография, — роль «указателя» на ситуации, которые перспективны для изучения с целью выявления направления и давления отбора. Возможно, что в ряде случаев феногеография и непосредственно поможет определить направления естественного отбора. Это можно сделать посредством выявления, например, корреляции в распространении какого-либо фена или группы фенов с распространениями каких-либо факторов среды. Эти факторы среды могут быть абиотическими (температура, влажность и т. п.), но могут быть и биотическими — связанными с другими организмами. Например, была обнаружена тесная связь между распространенностью фена семи боковых крупных чешуй у колюшки и наличием в водоеме хищных рыб. Некоторые фены у прыткой ящерицы в своем распространении совпадают с ареалом рябины в Евразии. Задача дальнейших экологических исследований — выяснить существо этой связи, но само ее существование вскрывается именно феногеографическим подходом.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Естественный отбор
Естественный отбор Итак, Дарвин показал, что основным средством создания всех пород домашних животных является искусственный отбор. Даже в то далёкое время, когда люди вели этот отбор, не ставя перед собой определённой цели, бессознательно, они достигали поразительных
Естественный отбор
Естественный отбор Я не усматриваю предела деятельности этой силы, медленно и прекрасно приспособляющей каждую форму к самым сложным жизненным отношениям. Ч. Дарвин Осы, бабочки и дарвинизмВ предыдущих главах мы неоднократно говорили о естественном отборе. Это и
Естественный отбор в природе и в лаборатории
Естественный отбор в природе и в лаборатории Действие отбора изучают не только в лабораторных экспериментах, но и в ходе многолетних наблюдений в природе. Первый подход позволяет контролировать условия среды, выделяя из бесчисленного множества реальных жизненных
ЕСТЕСТВЕННЫЙ ОТБОР: НЕ ВСЕ В ЭВОЛЮЦИИ ДВИЖЕТСЯ ВПЕРЕД
ЕСТЕСТВЕННЫЙ ОТБОР: НЕ ВСЕ В ЭВОЛЮЦИИ ДВИЖЕТСЯ ВПЕРЕД Шок Дарвина Леонардо да Винчи (1452–1519) говорил: «В природе нет ошибок, но знай: ошибка есть в тебе». Слишком совершенным казалось этому гению все богатство и разнообразие форм в природе, чтобы хоть немного усомниться в
ЕСТЕСТВЕННЫЙ ОТБОР
ЕСТЕСТВЕННЫЙ ОТБОР Естественным отбором считается процесс, который Дарвин назвал «борьба за существование», при котором наиболее приспособленные организмы выживают, а наименее приспособленные погибают. Согласно положениям дарвинизма, естественный отбор в популяции с
ЕСТЕСТВЕННЫЙ ОТБОР ПОД ВЛИЯНИЕМ ХИЩНИКОВ
ЕСТЕСТВЕННЫЙ ОТБОР ПОД ВЛИЯНИЕМ ХИЩНИКОВ Чарлз Дарвин осознавал важность размера популяции при определении выживаемости индивидов, соревнующихся между собой за ограниченное число ресурсов удовлетворения основных потребностей, в особенности за пищу. В этом процессе
ЧТО МОЖЕТ ГРУППОВОЙ ЕСТЕСТВЕННЫЙ ОТБОР
ЧТО МОЖЕТ ГРУППОВОЙ ЕСТЕСТВЕННЫЙ ОТБОР Групповой брак приводит к близкородственному скрещиванию и через несколько поколений делает всех членов группы близкими по набору генов. В такой ситуации не столь важно, мое или твое потомство выжило, я или ты погиб преждевременно.
Естественный отбор
Естественный отбор Естественный отбор – важнейший фактор эволюции. Дарвинизм (а именно на базе дарвинизма построена СТЭ), как уже отмечалось выше, называют теорией естественного отбора.Кратким и удачным определением отбора может служить сформулированное И. Лернером:
5.3. Естественный отбор и развитие этики сексуальных контактов
5.3. Естественный отбор и развитие этики сексуальных контактов Сила и продолжительность половой любви бывают такими, что невозможность обладания представляется обеим сторонам великим, если даже не величайшим несчастием; они идут на огромный риск, даже ставят на карту
7. ВОЙНЫ И ЕСТЕСТВЕННЫЙ ОТБОР
7. ВОЙНЫ И ЕСТЕСТВЕННЫЙ ОТБОР Высочайшее счастье для человека и его величайшая радость — разбить и уничтожить врага, стереть его с земли, забрать все, что он имел, заставить плакать его жен, скакать на его лучших и любимых лошадях и обладать его красивыми
Что может групповой естественный отбор
Что может групповой естественный отбор Групповой брак приводит к близкородственному скрещиванию и делает через несколько поколений всех членов группы близкими по набору генов. В такой ситуации не столь важно, чье — мое или твое — потомство выжило, я или ты погиб
9. Естественный отбор – главная движущая сила эволюции
9. Естественный отбор – главная движущая сила эволюции Вспомните!Какие виды отбора вы знаете?Назовите известные вам формы естественного отбора.Естественный отбор – это преимущественное выживание и размножение наиболее приспособленных особей каждого вида и гибель
Естественный отбор — быть сильнее своего животного начала
Естественный отбор — быть сильнее своего животного начала Нам же особенно важно, что это комендант своей силой заставляет тело следовать инстинктам. (Не пропустите этот момент!) То есть это комендант (его сила) определяет животное начало в теле. И с точки зрения физики
Глава 12 Естественный отбор: кто выживет?
Глава 12 Естественный отбор: кто выживет? Есть горстка ученых, пытающихся выявить признаки, общие для всех когда-либо существовавших человеческих культур и цивилизаций. Эти ученые роются в трудах этнографов и в статьях антропологов в поисках различий между племенами и
Естественный отбор — единственный направленный фактор эволюции
Естественный отбор — единственный направленный фактор эволюции Несомненно, важнейшим эволюционным фактором является естественный отбор. При определении естественного отбора Ч. Дарвин использовал понятие «выживание наиболее приспособленного». При этом имелось,