Как найти границы популяций?
Как найти границы популяций?
Популяции внутри любого вида отличаются друг от друга по частоте проявления разных аллелей, что внешне должно выражаться в различной концентрации разных фенов. Поэтому, если при исследовании видового населения в природе мы обнаруживаем резкий перепад в частоте каких-либо фенов, можно делать обоснованное предположение о существовании здесь популяционной границы.
Рассмотрим несколько характерных примеров.
На рис. 12 приведен пример первой целенаправленной феногеографической работы: доказана концентрация, фенов в популяциях кур в одном из районов Дагестана, изученная А. С. Серебровским. Здесь в период исследования куры вокруг аулов жили в полудиком состоянии, гнездились в кустарниках, поэтому их можно было рассматривать как модели настоящих природных популяций. Поскольку генетика основных признаков кур в то время уже была известна, А. С. Серебровский использовал результаты генетических исследований и перенес их на природные наблюдения, выделяя фены.
Исследование велось в окрестностях р. Аварское Койсу. Дагестан — страна горных ущелий, иногда в сотни метров глубиной. Аварское Койсу течет в одном из таких ущелий. Ширина ущелья несколько сот метров, но перелететь его куры не могут. Это — серьезный изоляционный барьер. Изоляция между другими изученными группами кур менее значительна. Группировки кур, обитающих на одной стороне реки, сходны по набору аллелей, но резко отличаются по этим показателям от кур, живущих на другом берегу.
Ширина популяционных границ зависит (хотя и не прямо) от степени подвижности организмов. Как правило, у более подвижных животных она больше, чем у малоподвижных, например у моллюсков. В ряде изученных случаев частота фенов полосатости и окраски раковины виноградной улитки в Англии и Франции резко менялась на протяжении всего 20–30 м на сплошном участке, заселенном этим видом.
На рис. 13 показана концентрация фена «краснохвостости» в населении обыкновенных белок в районе Верхней Волги. Как показали исследования И. С. Томашевского, на протяжении нескольких десятков километров частота этого фена резко изменяется, маркируя тем самым реальную природную границу между популяциями. Возможно, что это граница не одной популяции, а целой группы, так как именно здесь проходит граница между подвидами белок.
Множество примеров выделения популяционных границ ныне дает изучение биохимических фенотипов — частоты встречаемости разных изозимов. Стал классическим пример хорошо улавливаемых природных границ между группами популяций домовой мыши в Ютландии (Дания). Данные по двум разным фенам (эстераза 1 и 2) совпадают, хотя гибридная зона в каждом случае (по отношению к той и другой эстеразе) имеет специфическую конфигурацию и широту. Подобная ситуация очень типична. Так, например, гибридная зона между двумя популяциями ящериц (Cnemidoforus tigris), принадлежащих к разным подвидам, в юго-западной части штата Нью-Мехико (США), определяемая по морфологическим фенам чешуйчатости и окраски, составляет около 2 км, а определяемая по биохимическим фенам превышает 40 км.
Интересный пример определения популяционной границы — случай с обыкновенной двухточечной божьей коровкой. Она встречается в двух основных формах — черной и красной. По частоте этих форм различаются разные популяции и группы популяций внутри вида. По наблюдениям С. С. Сергиевского, в 1976–1977 гг. одна из резких границ, между популяциями, определяемая по изменению распространения этих форм, проходит по центральным районам Ленинграда — на протяжении нескольких сот метров.
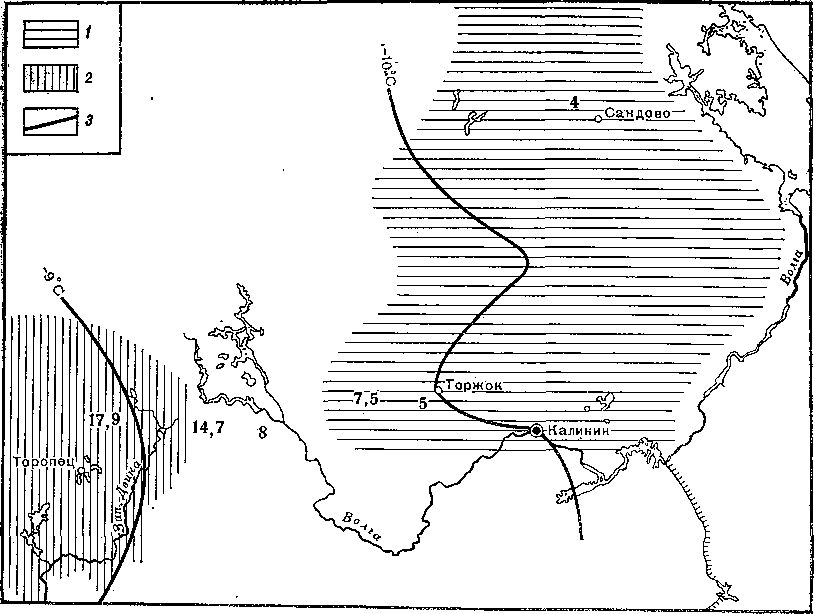
Рис. 13. Концентрация фена «краснохвостости» в популяциях обыкновенной белки в верховьях р. Волги 1 — западная популяция, 2 — северная популяция, 3 — изотерма января
Последними из этой серии примеров выделения фенетическими методами популяционных границ в природе приведем два случая, связанные с характером песни у птиц. Сейчас в связи с широким распространением портативных и эффективных способов записи песен число работ в этой области очень велико. Параболические рефлекторы позволяют с высокой точностью записывать голоса отдельных птиц на расстоянии до сотен метров. К югу от г. Аккра на побережье Гвинейского залива резкая граница между группами блестящих нектарниц с резко различными песнями проходила на площади шириной всего в 50 м и оставалась стабильной на протяжении нескольких лет.
Другое популяционное исследование песни касается дроздов-белобровиков в окрестностях г. Осло. На территории в 85 км обитало семь разных групп дроздов с характерными диалектами песни, устойчиво сохраняющимися на протяжении всего периода изучения в течение двух-трех поколений. Выборочное исследование территории в 250 км2 показало, что группы в 10–12 диалектов составляют «супердиалект» — большую группу родственных диалектов, по всей вероятности являющуюся настоящей популяцией.
В природе при определении популяционных границ можно встретить две принципиально разные ситуации. Границы между популяциями могут быть чрезвычайно резкими, легко определяемыми или нечеткими, размытыми, причем популяции связаны целой гаммой постепенных переходов по концентрации отдельных фенов. В последнем случае феногеографические методы помогают выделить только популяционные центры. При этом подходе можно сравнивать произвольно взятые выборки из разных районов и определять, относятся ли они к одной общей популяции (по частоте фенов). Приведу несколько характерных примеров, связанных с изучением окраски.
На спине взрослых самцов гренландских тюленей всегда присутствует яркий черно-белый рисунок, издалека напоминающий крылья (отсюда поморское название этих животных — крыланы). Рисунок всегда различается в деталях. Наши исследования показали, что по частоте проявлений основных типов рисунка (рис. 14) группа гренландских тюленей (Pagophilus groenlandicus), обитающих в водах о-ва Ньюфаундленд, отличается от тех, которые размножаются в водах Гренландского моря (район о-ва Ян-Майен) и в Белом море. Такие статистически достоверные различия в частоте разных типов окраски послужили одним из аргументов в пользу генетической самостоятельности изученных группировок, их различной популяционной принадлежности (что впоследствии хорошо подтвердилось мечением). Другой пример. По частоте белого пятна на груди среди просмотренных 1183 кошек на улицах Парижа выяснено, что здесь живут в диком состоянии по крайней мере три популяции кошек (генетическая обусловленность этого признака окраски давно известна). Подобного рода примеров немало.
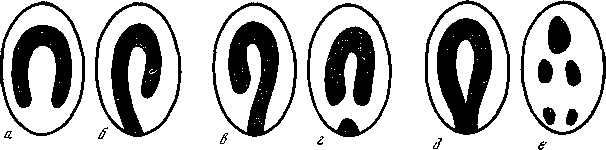
Рис. 14. Различные типы окраски (а — е) самцов гренландского тюленя, по концентрации которых обнаружены резкие различия между популяциями
Нет другого существа, так хорошо изученного по форме тела, как человек. В этнической антропологии есть буквально сотни, если не тысячи примеров различий отдельных популяций людей но разным, генетически простым признакам (фенам).
В табл. 4 приведены три таких примера, касающиеся фенов, изучение которых в мире животных пока практически невозможно: способности ощущать вкус фенилтио-мочевины (одни люди не ощущают его вообще, другие считают горьким как хинин); цветовой слепоты, при которой люди (дальтоники) не различают красный и зеленый цвета; частоты рождения дизиготных близнецов (по Дж. Харрисону и др., 1968).
Два следующих примера из этой серии относятся к изучению поведения. Индивидуальная изменчивость пения птиц, о которой уже говорилось выше, была известна натуралистам и любителям очень давно. Давно известны и географические различия в пении птиц. Различаются курские и киевские соловьи, тюрингенские и франкфуртские овсянки, многие другие примеры широко известны еще с XVIII–XIX вв.
Таблица 4.
Фонетическая характеристика (частота признака в %) некоторых популяций человека.
Популяция Ощущение вкуса фенилтио-мочевины Популяция Цветовая слепота Популяция Число дизиготных двоен на 1000 родов Хинди 33,7 Бельгийцы 8,6 Негры (Нигерия) 39,9 Датчане 32,7 Хинди 8,1 Англичане 31,5 Норвежцы 8,0 Негры (Заир) 18,7 Испанцы 25,6 Норвежцы 8,0 Малайцы 16,0 Шотландцы 7,8 Греки 10,9 Японцы 7,1 Немцы 7,5 Англичане 8,9 Лопари 6,4 Китайцы 6,2 Шведы 8,6 Китайцы 2,0 Мексиканцы 2,3 Итальянцы 8,6 Индейцы(Бразилия) 1,2 Индейцы(США) 2,0 Французы 7,1 Испанцы 5,9 Негры(Заир) 1,7 Японцы 2,7В средней полосе нашей страны для таких исследований удобен зяблик с его сравнительно простой песней. В одной из работ известный зоолог А. Н. Промптов сравнивал песни зябликов из двух мест Московской области и из Западного Урала. На основании анализа нескольких тысяч песен он установил, что под Звенигородом (70 км от Москвы) чаще встречаются особи с двух- и четырехколенными песнями, тогда как певцы с трехколенными песнями чаще живут вблизи Москвы. На Западном Урале четырехколенные певцы вообще отсутствуют. Зато на Урале значительно чаще, чем в Московской области, во втором колене песни встречаются свистовые варианты; под Москвой в этом колене преобладают трескучие звуки. Наконец, на Урале обнаружены одноколенные варианты песни, никогда не встречавшиеся в Подмосковье. В таком исследовании привлекает чрезвычайная простота: нет необходимости в сложных звукозаписывающих и расшифровывающих установках, достаточно записной книжки и карандаша. Сейчас практически для всех птиц Европы и Северной Америки получены данные по географической изменчивости элементов песни. Существенное преимущество описанных приемов в том, что не надо убивать животных для получения данных по фенофонду и феногеографии.
Итак, все эти примеры показывают два главных пути выделения популяций в природе: либо посредством поиска резких границ, маркируемых заметными перепадами распространенности (концентрации) фенов, либо путем сопоставления фенетических характеристик отдельных групп особей (чтобы проверить, не относятся ли они к различным генетическим группировкам). Естественно, что для детального исследования вполне возможно и сочетание этих двух подходов: сначала в общей форме, чтобы выделить популяции по сравнению групп особей, а затем попытаться найти более резкую природную границу между ними.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
8.3. «Самоорганизация» на уровне популяций
8.3. «Самоорганизация» на уровне популяций Уделяя особое внимание морфофизиологическим структурам отдельного организма, изучая громадное, почти непостижимое разнообразие форм, очень легко оказаться в плену организмоцентризма, т. е., грубо говоря, считать, что структура
ГРАНИЦЫ ИНТЕЛЛЕКТА
ГРАНИЦЫ ИНТЕЛЛЕКТА Человекообразные обезьяны, без сомнения, больше других животных приближаются к человеку по строению тела, но, рассматривая жизнь муравьев, их общественную организацию, их обширные сообщества, тщательно устроенные жилища и дороги, их владение
Какая европейская река меняет свое название после пересечения государственной границы?
Какая европейская река меняет свое название после пересечения государственной границы? Берущая начало в России, на Валдайской возвышенности, река Западная Двина несет свои воды через две государственные границы – сначала российско-белорусскую, а затем
Генетика популяций
Генетика популяций Генетика популяций постулирует, что единица эволюционного процесса должна представлять неделимое единство и быть способной изменяться в ряду поколений. Ни вид, ни особь не удовлетворяют этим критериям. Элементарной единицей эволюционного процесса
3.1.1. Температурные границы существования видов
3.1.1. Температурные границы существования видов В среднем активная жизнедеятельность организмов требует довольно узкого диапазона температур, ограниченного критическими порогами замерзания воды и тепловой денатурации белков, примерно в пределах от 0 до +50 °C.Границы
8.2.2. Классификация популяций
8.2.2. Классификация популяций Экологи руководствуются различными принципами для выделения и классификации популяций как территориальных группировок в пределах вида.Н. П. Наумов на примере млекопитающих рассматривает вид как иерархическую систему популяций различных
8.5. Динамика популяций
8.5. Динамика популяций 8.5.1. Биотический потенциал Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины
8.5.4. Стратегии выживания популяций
8.5.4. Стратегии выживания популяций Различия в биотическом потенциале видов зависят от их размеров, систематической принадлежности и других причин, но при прочих равных условиях связаны зависимостью со смертностью в популяциях. Эта закономерность, подмеченная еще Ч.
8.5.8. Гомеостаз популяций
8.5.8. Гомеостаз популяций Поддержание определенной плотности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа
8.6.1. Модификация и регуляция популяций
8.6.1. Модификация и регуляция популяций Современная теория рассматривает динамику численности популяций как авторегулируемый процесс. Любой популяции организмов в конкретных условиях свойствен определенный средний уровень численности, вокруг которого происходят
Я верю, что где-то есть на свете «моя половина», но как мне её найти?
Я верю, что где-то есть на свете «моя половина», но как мне её найти? Не верьте. Концепция «двух половин», предполагающая наличие где-то одного, единственно подходящего данному человеку (и более никому другому) партнёра – поэтичный миф, и не более того. В брачном поиске люди
5.1. Границы биосферы
5.1. Границы биосферы Биосфера является одним из трех (гидросфера, атмосфера и литосфера) компонентов климатической системы. Ее можно уподобить тонкой пленке, покрывающей поверхность нашей планеты. Плотность органического вещества равна 1 г/см2. Для сравнения, средняя
Как найти фены?
Как найти фены? В генетике для анализа того или иного признака достаточно скрестить пару особей и по характеру распределения признаков в потомстве (на протяжении ряда поколений) точно определить особенности любого признака или свойства. Для фенетического анализа