Как найти фены?
Как найти фены?
В генетике для анализа того или иного признака достаточно скрестить пару особей и по характеру распределения признаков в потомстве (на протяжении ряда поколений) точно определить особенности любого признака или свойства. Для фенетического анализа необходимо значительное число особей. Без них исследователь не сможет составить достаточно полное представление о существующих в природе дискретных вариациях, среди которых и приходится искать фены.
Опыт, накопленный при исследовании фенов в разных группах животных и растений, позволяет предложить следующие пути выделения фенов на материале природных популяций.
1. Сопоставление наблюдаемой изменчивости по отдельным комплексам признаков или свойствам (окраске, рисунку, форме отдельных частей тела, вариантам строения отдельных органов и систем и т. п.).
2. Выделение признаков, подверженных возрастным и половым изменениям. Для первичного анализа они сложны, хотя среди них тоже можно найти фены, но это — более специальная задача. Обычно такие признаки не включаются в фенетическое исследование на животных, хотя, как упоминалось выше, могут быть с большим успехом использованы у растений.
3. Выделение в общей изменчивости признаков и свойств дискретных характеристик, их анализ на дальнейшую делимость (дробимость, подразделяемость).
Предположим, что изучаются такие особенности окраски, как рисунок и цвет. Дискретность в изменчивости нужно искать именно в цветовой гамме, а не в микроскопически определяемых характеристиках пигмента или микроструктуре поверхности органа, вызывающего цветовые эффекты (как у чешуекрылых). Равным образом было бы неправильно свести рисунок окраски (полосы, пятна на отдельных участках тела и т. п.) к изучению, например, гистологии кожного покрова. И в том и в другом случае исследователь потеряет качество изучаемой группы признаков и перейдет к изучению совсем другой группы признаков и свойств (среди которых тоже можно искать качественные альтернативные дискретные признаки).
4. Анализ данных по генетике филогенетически близких генетически изучаемых форм. Чтобы проанализировать окраску рыжей полевки, нужно познакомиться с работами по генетике окраски других грызунов (домовая мышь, крысы и др.). Зная закон гомологических рядов в наследственной изменчивости, можно считать, что общие закономерности окраски млекопитающих будут в чем-то существенном общими. За цвет покровов у млекопитающих несут ответственность шесть основных, сходных у разных видов млекопитающих генов, а за рисунок — еще несколько таких же генов. Конечно, у каждого вида несколько основных генов могут дополняться другими, их действие может в разной степени модифицироваться, но сам факт наличия гомологичных генетических основ позволяет уверенно вести фенетический анализ окраски практически всех млекопитающих.
5. Анализ косвенных данных о характере наследования отдельных признаков у данного вида (выражение признаков у родителей и потомков и т. п.).
6. Проверка правильности выделения фенов на природном материале. Обычно правильно выделенные фены на достаточно разнообразном материале из природных популяций обязательно покажут или определенные тенденции в распространении их в пределах ареала вида, или изменение во времени (на палеонтологическом материале).
Теперь рассмотрим описанный порядок выделения фенов на конкретных примерах.
Пример I. Цель исследования — анализ внутривидовой изменчивости окраски (цвета) покрова прыткой ящерицы для выяснения особенностей процесса микроэволюции на уровне отдельной популяции, групп популяций и всего вида в целом. Таким образом, представляли интерес признаки от самого мелкого до самого крупного масштаба.
1. Изучение вариантов окраски по цвету покровов из нескольких природных популяций показало присутствие широкого спектра серо-зеленых и желто-коричневых тонов в окраске спины, бело-голубовато-зеленых в окраске брюха, бело-желтых в окраске горла.
2. Сопоставление окраски молодых и взрослых животных показало, что существуют значительные возрастные изменения окраски — окраска спины молодых однородно коричнево-серая. Кроме того, окраска брюха и горла также изменяется с возрастом. Поэтому для дальнейшего анализа использовались лишь половозрелые особи, легко выделяемые по размерам тела.
Исследование окраски самцов и самок показало сложную картину: в одних популяциях половой диморфизм и окраске спины выражался резко (самцы зеленые, самки коричневые), в других — и среди самцов и среди самок встречались коричневые и зеленые особи. Сложным оказался и признак окраски горла: как правило, самцы имели более ярко окрашенное горло (ярко-желтое или даже зеленое), но нередко встречались и самки с желтым горлом. Окраска брюха у самцов более зеленая, чем у самок, но варьировала и у того и у другого пола весьма значительно.
3. Очень широкий спектр цветовых вариаций включал первоначально восемь цветовых оттенков для окраски спины (темно-зеленый, ярко-зеленый, светло-зеленый, желтый, светло-коричневый, красноватый, коричневый, темно-коричневый) и по два-три оттенка окраски брюха и горла. Для проверки правильности отнесения той или иной особи к определенной группе окраски были составлены специальные цветовые таблицы. Однако практика применения таблиц показала субъективность восприятия разными наблюдателями выделенной первоначально цветовой гаммы. Безусловно совпадающим у всех исследователей оказалось лишь выделение зеленого и коричневого цветов в окраске спины (впоследствии в качестве очень редких вариантов были найдены «красные» — кирпичного цвета — и черные особи). В качестве дискретных признаков были выделены зеленые и коричневые тона в окраске спины, и пришлось полностью отказаться от точного учета окраски брюха и горла.
4. Имеющиеся в литературе данные по наследованию окраски у прыткой ящерицы касались менделевского характера наследования кирпично-красной окраски спины. Данные по генетике окраски ящериц другого рода (американских Sceloporus graciosus) показали высокую наследственную обусловленность песчано-красной окраски спины.
5. Косвенные данные по наследованию окраски в природе и эксперименте практически отсутствовали из-за значительного возрастного полиморфизма в окраске и несравнимости окраски взрослых и неполовозрелых особей. Известны наблюдения единичных черноокрашенных и молодых и взрослых особей в одних и тех же местообитаниях (это позволило предполагать их близкое родство).
6. Анализ частоты встречаемости зеленого и коричневого цвета в окраске спины у самцов и самок показал в целом определенную закономерность: на севере и самцы и самки были окрашены в коричневый цвет, на юге практически все самцы имели зеленую окраску, тогда как среди самок встречались и коричневые и зеленые особи. Иными словами, и для самцов и для самок было установлено закономерное изменение частоты встречаемости зеленой и коричневой окраски с юга на север, причем характер этой закономерности у самцов и самок отличался. Таким образом, фены окраски показали различие в масштабе крупных частей ареала вида. При анализе же внутри отдельных популяций эти признаки не дали каких-либо определенных результатов — их распределение было хаотично.
Пример II. Этот пример касается анализа изменчивости швов черепа гренландского тюленя. Была поставлена задача найти доказательства популяционной самостоятельности трех группировок этого вида в Северной Атлантике. Имевшийся материал включал черепа животных разного возраста (от 2 до 36 лет), добытых во всех трех предполагаемых популяциях.
1. Сопоставление всей наблюдаемой изменчивости показало, что швы бывают полностью заросшими, без каких-либо следов соединения костей, бывают хорошо заметными и разной формы (рис. 6). Кроме того, встречались черепа, у которых на правой половине швы были хорошо развиты, а на левой отсутствовали.
2. Сравнение черепов разновозрастных особей показало, что заращение швов не имеет прямой связи с возрастом: у самых старых животных были найдены хорошо выраженные швы, тогда как у молодых в некоторых случаях швы зарастали полностью. Половой диморфизм в развитии швов не обнаружен. Все это позволило оставить для дальнейшего анализа все первоначально выделенные признаки присутствия или отсутствия швов хотя бы на одной стороне и их формы.

Рис. 6. Разная форма швов в черепе млекопитающих
3. Присутствие швов оказалось дискретным признаком: не было найдено животных с их частичным присутствием.
4. Анализ данных по генетике показал, что некоторые линии домовых мышей различались по степени зарастания швов к моменту достижения половой зрелости (у всех мелких млекопитающих с возрастом швы зарастают). Из антропологии известны случаи наследственного сохранения несросшимися швов черепа в течение всей жизни. В литературе имеются данные о нахождении незаросших швов черепа у старых особей самых разных видов млекопитающих, в том числе у многих хищных — отряда, филогенетически близкого к ластоногим.
5. Данные по сравнению характера швов черепа у родителей и потомков из природных популяций отсутствовали.
6. Анализ частоты встречаемости разного типа швов в трех популяциях гренландского тюленя показал существование различий между популяцией о-ва Ньюфаундленд, с одной стороны, и Гренландского и Белого морей — с другой. Эти различия выявлены при сравнении встречаемости незаросших швов, а не форм швов. Фен присутствия швов оказался маркером, позволяющим подтвердить самостоятельность одной из изученных популяций.
Кратко охарактеризованный путь выделения фенов на зоологическом материале применим и для работы с растениями. При выделении фенов важно обратить внимание на их достаточную многочисленность.
Для решения каких-то сравнительно простых вопросов популяционного исследования иногда бывает достаточно одного фена. Например, в тех случаях, когда фен в одной популяции присутствует в высокой концентрации, а в другой — отсутствует или присутствует в ничтожных концентрациях. В этих случаях уже можно сделать обоснованное предположение о существовании границ между исследованными совокупностями особей.
Однако опыт работы Н. И. Вавилова с разными видами растений, наш опыт работы с ящерицами, анализ наиболее успешных случаев использования фенетических методов на других видах показывает, что для надежной и полной характеристики всего диапазона пространственной изменчивости (от выделения отдельных семей до выделения подвидов) надо учитывать многие десятки (а то и несколько сотен) признаков. Тогда есть полная уверенность, что любая задача популяционного анализа будет успешно решена.
Не надо бояться потерять много времени на выделение фенов. Дело в том, что все сказанное относится именно к исследованиям какой-либо группы, ранее не изучавшейся с целью выделения признаков, которые могут быть приняты как фены. В группах, где проведены такие исследования, ситуация складывается скорее обратная. Если по какому-либо виду уже собран фенетический материал (другими исследователями, в других районах и в другое время), то каждая последующая работа оказывается способной привести ко все более интересным и глубоким выводам. Возникает полная аналогия с выкладыванием картины из мозаики: после добавления одного-двух элементов прежде казавшиеся рассыпанными в беспорядке частицы складываются в общую осмысленную картину, проявляются сложные узоры и рисунки.
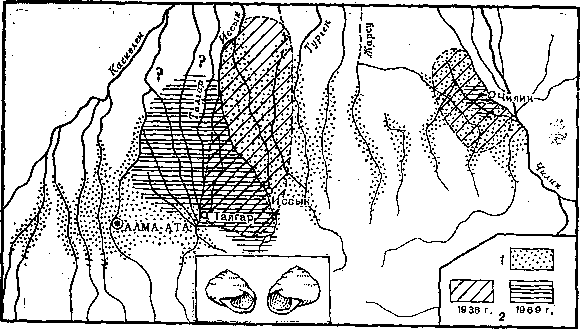
Рис. 7. Распространение право- и левозакрученных форм наземного моллюска и изменение ареала левозакрученных форм в Заилийском Алатау на протяжении 30 лет (с 1936 по 1969 г.)
1 — правозакрученные моллюски, 2 — левозакрученные
Приведем только один пример, касающийся исследований фена — левозакрученности раковин у наземных легочных моллюсков (Fruticicola lantzi) в предгорьях Заилийского Алатау. Левозакручениость — типичный фен, имеющий у всех изученных других видов моллюсков моногенную основу. Распространенность этого фена в популяциях Средней Азии первоначально изучена в конце 30-х годов, повторно — в конце 60-годов. Оказалось, что в одной группе популяций территория, занятая особями с этим феном, во много раз сократилась, а в другой — сдвинулась на несколько десятков километров (рис. 7). Каждое новое исследование этого интересного природного эксперимента будет все более простым и в то же время информативным, позволяя выяснять динамику генетического состава популяций.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Я верю, что где-то есть на свете «моя половина», но как мне её найти?
Я верю, что где-то есть на свете «моя половина», но как мне её найти? Не верьте. Концепция «двух половин», предполагающая наличие где-то одного, единственно подходящего данному человеку (и более никому другому) партнёра – поэтичный миф, и не более того. В брачном поиске люди
Как найти границы популяций?
Как найти границы популяций? Популяции внутри любого вида отличаются друг от друга по частоте проявления разных аллелей, что внешне должно выражаться в различной концентрации разных фенов. Поэтому, если при исследовании видового населения в природе мы обнаруживаем