Случайность и закономерность
Случайность и закономерность
Одно из центральных утверждений синтетической теории эволюции (СТЭ) состоит в том, что все эволюционные изменения основаны на отборе случайных, ненаправленных мутаций. Однако эволюция совсем не похожа на случайный, хаотический процесс. В ней явно есть направленность. Например, очевидно, что господствующие на Земле формы жизни постепенно становятся все более сложными. Как же так? Может ли направленный и как будто вполне осмысленный процесс идти на основе случайных, хаотических наследственных изменений — мутаций? В более общей форме этот вопрос можно сформулировать так: каково соотношение случайности и закономерности в эволюции? Этот вопрос был и остается одним из центральных в теоретической биологии и одновременно одним из самых спорных. Отчасти мы уже ответили на него в главе 4, где говорилось, в частности, о том, что естественный отбор — то есть избирательное запоминание системой возникающих изменений — неизбежно придает эволюции упорядоченный, направленный характер.
Особенно ярко проявляется закономерный характер эволюции в явлении параллелизма. Так называют те случаи, когда у разных организмов независимо возникают похожие признаки. Мы уже вскользь упоминали об этом явлении, когда говорили о приспособлении бактерий к антибиотикам (глава 4). Пришло время поговорить о параллелизмах подробнее.
Некоторые примеры общеизвестны. Так, представители разных классов позвоночных животных, перешедшие к активному плаванию в толще воды, независимо друг от друга приобрели обтекаемую рыбообразную форму тела и плавники (рыбы, ихтиозавры, дельфины). Еще более интересны примеры возникновения почти идентичных наборов «жизненных форм» на разобщенных континентах. Например, Южная Америка долгое время была изолирована от других материков, но еще до отделения туда проникли примитивные плацентарные млекопитающие кондиляртры — «древние копытные». Эволюция кондиляртр протекала далее независимо в Южной и Северной Америке, причем последняя время от времени соединялась с Евразией сухопутным мостом — Берингией. В результате на «Большой земле» (в Северной Америке и Евразии) кондиляртры дали начало отрядам парно- и непарнокопытных, куда относятся всем известные лошади, носороги, антилопы, олени, гиппопотамы и т. д. В Южной Америке расцвели другие отряды копытных, произошедшие от тех же кондиляртр — нотоунгуляты[68], литоптерны, астрапотерии и пиротерии. Среди этих «южных копытных» появились формы, поразительно схожие с настоящими лошадьми, носорогами, верблюдами, зайцами и даже слонами. В Южной Америке процветали также и сумчатые, быстро вымершие на «Большой земле». Среди южноамериканских сумчатых тоже появились жизненные формы, удивительно похожие на некоторых североамериканских и азиатских зверей. Один из ярких примеров — сумчатый саблезубый тигр тилакосмилус, аналог настоящих саблезубых тигров, которые относились к семейству кошачьих и охотились на крупных толстокожих копытных на просторах Азии, Африки и Северной Америки. Тилакосмилус, хоть и был сумчатым, вероятно, занимал ту же экологическую нишу и охотился на крупных толстокожих «южных копытных»[69].
Впрочем, не стоит преувеличивать это сходство. В Южной Америке появились и такие звери, аналогов которым среди обитателей «Большой земли» найти не удается. Это броненосцы, муравьеды, ленивцы, к которым относятся и удивительные гигантские наземные ленивцы мегатерии. Все эти звери не похожи ни на кого, только на самих себя. В целом южноамериканские звери были менее совершенными, чем их аналоги с «Большой земли». Например, сумчатые хищники вроде тилакосмилуса все-таки не могли сравниться в ловкости и сообразительности с «настоящими», несумчатыми, львами и тиграми. Поэтому с южноамериканскими хищными млекопитающими успешно конкурировали гигантские хищные нелетающие птицы фороракосы. Кроме того, там появились самые крупные летающие птицы всех времен. Некоторые из них, такие как аргентавис, достигали в размахе крыльев семи с половиной метров при весе до 80 кг и, возможно, могли проглотить целиком зверя размером с зайца. Совсем недавно в Патагонии был найден самый большой в мире птичий череп. Его длина — 71,6 см.
—————
Самый большой птичий череп всех времен. Фороракосы — крупные нелетающие хищные птицы, появившиеся в Южной Америке около 62 млн лет назад, то есть почти сразу после вымирания динозавров. Многие палеонтологи полагают, что между этими двумя событиями существует тесная связь. Исчезновение динозавров освободило нишу двуногого бегающего хищника, которая активно эксплуатировалась разнообразными хищными динозаврами в течение всей истории этой группы рептилий. После вымирания динозавров эта ниша была немедленно освоена их ближайшими родственниками — птицами. Семейство фороракосов, последние представители которого вымерли около 2 млн лет назад, включает самых крупных птиц всех времен. Их рост мог достигать трех метров и более. Однако многие детали строения крупных фороракосов остаются неизвестными из-за фрагментарности имеющихся находок.
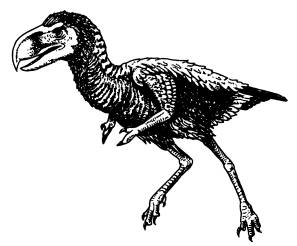
Titanis walleri — родственник фороракоса, живший 2 млн лет назад в Техасе и Флориде. На передних конечностях у него имелась пара пальцев, образующих подобие клешни.
В 2006 году палеонтологи нашли в Аргентине неплохо сохранившийся череп фороракоса длиной 716 мм. Это самый большой птичий череп из всех известных науке. Возраст находки — 10–15 млн лет (средний миоцен). Вместе с черепом найдены кости ног, которые оказались неожиданно длинными и тонкими. Это противоречит прежним представлениям о том, что крупнейшие представители фороракосов были тяжеловесны и медлительны. По всей видимости, эти чудовищные птицы на самом деле были очень быстрыми бегунами.
(Источник: Luis M. Chiappe, Sara Bertelli. Skull morphology of giant terror birds // Nature. 2006. V. 443.)
—————
Несмотря на удивительное разнообразие и кажущуюся высокую приспособленность южноамериканских зверей, многие из них не выдержали конкуренции с североамериканскими переселенцами: после воссоединения Южной и Северной Америк, а это произошло в плиоцене, многие северные виды отправились осваивать южные территории. Большинство южноамериканских хищников и копытных были вытеснены пришельцами и вымерли.
В этом проявилось одно из «правил» эволюции, которому пока нет однозначного объяснения. На больших территориях быстрее идут эволюционный прогресс и рост конкурентоспособности, тогда как на малых территориях, в том числе на изолированных островах, быстрее происходит появление всевозможных странных, необычных, причудливых форм. Это обстоятельство биологи подметили уже давно, но лишь недавно сумели его подтвердить строгими цифрами.
—————
Эволюция на островах идет быстрее. Еще со времен Дарвина, исследовавшего галапагосских вьюрков, удаленные острова считаются своеобразными «эволюционными лабораториями», в которых виды претерпевают быстрые и порой весьма изощренные эволюционные изменения. В частности, известно, что крупные животные, попав в изолированные и обедненные островные экосистемы, часто мельчают, а мелкие, наоборот, достигают гигантских размеров, причем в ряде случаев на это требуется совсем немного времени. Однако до сих пор представления об ускоренной эволюции на островах не имели строгого количественного обоснования, а отдельные факты и вовсе им противоречили: например, ископаемому сицилийскому слону Elephas falconeri, рост которого был менее метра, а масса (100 кг) составляла лишь 1% от массы его континентального предка, потребовалось очень много времени (около 300 тысяч лет), чтобы так измельчать.
Чтобы точно измерить темпы эволюционных изменений на островах и сравнить их с аналогичными данными по материковым популяциям, необходимо было собрать весьма значительный объем палеонтологических данных, причем весь материал должен был быть точно датирован. К настоящему времени палеонтология уже накопила достаточно данных для такого анализа, что и позволило Вирджинии Миллиен из Университета Макгилла (Канада) получить строгие оценки темпов эволюции на островах и на «Большой земле».
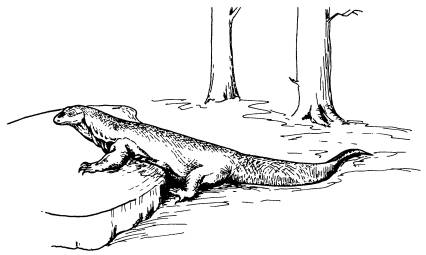
Гигантский варан, обитающий на островах Комодо, Ринджа, Падар и Флорес (Индонезия) — один из общеизвестных случаев «островного гигантизма».
Миллиен обработала данные по 86 островным и 84 континентальным популяциям млекопитающих, относящихся к 88 видам и 14 отрядам. Для каждой популяции была подсчитана скорость эволюционных изменений одного или нескольких размерных признаков (от 1 до 57 признаков для каждой популяции, в среднем 5,06 признака на популяцию) в течение определенного интервала времени (от 20 до 12 000 000 лет, средняя величина интервала — 462 тысячи лет). Использовались только линейные признаки (длина черепа, отдельных костей, зубов и т. д.).
Выяснилось, что морфологическая эволюция на островах действительно идет быстрее, однако этот эффект заметен лишь на временных интервалах менее 45 тысяч лет. При увеличении временного интервала выше этого порога различия между островными и континентальными популяциями становятся статистически незначимыми.
Выявленные различия нельзя объяснить разным таксономическим составом островной и континентальной биоты (то есть тем, что на островах некоторые группы млекопитающих встречаются чаще, чем на континенте, и наоборот). Оказалось, что темпы эволюции не являются «филогенетически консервативными». Например, виды, эволюционировавшие на континенте медленнее других, вовсе не обязательно будут продолжать вести себя сходным образом и на изолированном острове.
По-видимому, млекопитающие, попавшие на удаленные острова, очень быстро приспосабливаются к новым условиям, что порой сопровождается разительными изменениями размеров и пропорций тела. Из-за высокой скорости этих изменений в большинстве случаев палеонтологам не удается обнаружить переходных форм между островными эндемиками и их континентальными предками. После относительно короткого периода быстрой адаптации темп эволюции островных животных, как правило, замедляется (поэтому не удалось выявить различий в темпах эволюции на временных интервалах свыше 45 тысяч лет).
Фрагментация естественных местообитаний (в том числе и в результате деятельности человека) может ставить животных в условия, аналогичные попаданию на изолированные острова. В этом случае можно ожидать резкого ускорения эволюционных изменений по «островному» сценарию. Реальность таких явлений подтверждается фактами. Например, было показано резкое ускорение эволюционных изменений размеров тела за последние 200 лет у 25 видов млекопитающих в Дании. И это явление связывают с фрагментацией местообитаний.
Полученные результаты показывают, что многие виды континентальных млекопитающих потенциально способны к очень быстрым эволюционным преобразованиям. Темп эволюции может возрасти в три и более раза всего за пару десятилетий. Получается, что в условиях растущей антропогенной нагрузки на природные экосистемы многие привычные нам животные могут начать изменяться самым неожиданным и быстрым образом.
(Источник: Milleien V. 2006. Morphological Evolution Is Accelerated among Island Mammals.// PLoS Biol 4(10): e321.)
—————
Но вернемся к теме параллелизмов. В последние десятилетия существенно изменились представления о том, каким образом происходили в ходе эволюции крупнейшие прогрессивные изменения — ароморфозы[70]. Оказалось, что практически во всех хорошо изученных случаях переход на более высокий уровень организации происходит не в какой-то одной эволюционной линии, а в нескольких, развивающихся параллельно. При этом отдельные признаки, из которых складывается ароморфоз, иногда появляются в разных линиях почти одновременно, а иногда — в разное время и даже в разном порядке. Прогрессивные признаки постепенно накапливаются, пока наконец в одной или немногих линиях они не соберутся все вместе, и тогда палеонтологи «объявляют» о рождении новой группы.
В этом ярко проявляется закономерный характер эволюции. В какой-то момент словно начинает носиться в воздухе новая «идея» — например, идея млекопитающих. И многие разные группы не сговариваясь начинают развиваться в одном и том же направлении, хотя и немножко разными путями. Чтобы подчеркнуть это обстоятельство, многие биологи в последние годы стали называть происхождение млекопитающих «процессом маммализации зверообразных рептилий», происхождение птиц — «орнитизацией», амфибий — «тетраподизацией», членистоногих — «артроподизацией», покрытосеменных растений — «ангиоспермизацией» и т. д.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Неустранимая случайность
Неустранимая случайность Итак, онтогенез — это процесс самоорганизации, в ходе которого согласованные действия множества одинаково запрограммированных клеток, следующих сравнительно простому набору правил поведения, приводят к самосборке сложных многоклеточных