9.1. Головной мозг
9.1. Головной мозг
В анатомии мозга позвоночных обычно выделяют пять отделов, а у млекопитающих – шесть.
Продолговатый мозг (myelencephalon) является продолжением спинного мозга и, в общем виде, сохраняет его структуру, особенно у низших позвоночных. У высших позвоночных в продолговатом мозге четко выделяются отдельные ядра, в которых локализуются центры рефлекторных актов: дыхания, глотания, сосания, сердечной деятельности, равновесия.
Мозжечок (cerebellum) – это мозговой центр координации движений. Он не имеет самостоятельных двигательных систем, но образует связи со всеми системами мозга, связанными с движением. В мозжечок поступает информация о положении тела и состоянии мышц из среднего и переднего мозга, ретикулярной формации, разнообразных сенсорных систем. Эволюционно мозжечок возникает в результате интенсивного развития вестибулярных ядер.
У млекопитающих кора мозжечка состоит из трех слоев. Нейроны второго слоя – клетки Пуркинье, имеют весьма специфичное строение, образуя густую сеть из десятков тысяч отростков дендритов. Каждая клетка Пуркинье образует более 200 000 синаптических контактов. Кору приобрел в ходе эволюции и мозжечок птиц. Мозжечок птиц и млекопитающих достигает больших размеров из-за необходимости обработки огромного массива сенсорной информации, обусловленной сложностью координации их движений.
Мост (pons) как особый отдел выделяется только у млекопитающих. Он представлен в основном проводящими путями, которые связывают передний, средний, продолговатый и спинной мозг.
Средний мозг (mesencephalon) сохраняет вид трубки с полостью – сильвиевым водопроводом. Выше его располагается область, эволюционно связанная с развитием зрения, – тектум. У рыб и амфибий тектум является важнейшим мозговым центром, где обрабатываются сенсорные сигналы из всех источников. У рептилий тектум становится настоящим ассоциативным центром мозга. Огромно его значение и у птиц.
У млекопитающих тектум теряет свое значение и принимает форму четверохолмия. В нем располагаются подкорковые центры зрения и слуха, осуществляющие рефлекторные ориентировочные реакции. Большая часть сенсорных сигналов у млекопитающих направляется в полушария переднего мозга. Самое крупное ядро среднего мозга – красное ядро, в котором переключаются эфферентные нейроны из мозжечка и контролируется тонус мышц.
Промежуточный мозг (diencephalon) претерпевает в ходе эволюции позвоночных значительные преобразования. Он формировался как отдел, в котором сосредоточены многочисленные центры нейрогормональной регуляции. До сих пор не решен вопрос о гомологии его отделов у разных классов.
В промежуточном мозге выделяют три отдела: эпиталамус, таламус, гипоталамус.
Эпиталамус состоит из группы ядер и особой нейроэндокринной железы – эпифиза, роль которой долгое время оставалась непонятной. Р. Декарт рассматривал эпифиз как «хранилище души», и этот взгляд нашел широкую поддержку. В настоящее время эпифизу приписывают контроль циркадных ритмов, времени полового созревания, хотя сам механизм такой регуляции во многом непонятен. В эволюции позвоночных эпифиз развивается на основе теменного глаза, содержащего светочувствительные элементы.
Таламус претерпевает наибольшие структурные преобразования в эволюционном ряду позвоночных. Его роль возрастает по мере повышения уровня организации, при этом синхронно уменьшается роль тектума. У млекопитающих таламус включает более 150 ядер и становится «центральной переключательной станцией» мозга, где собирается вся сенсорная информация, направляемая затем в полушария переднего мозга.
Гипоталамус у всех классов позвоночных служит интегративным центром вегетативной нервной системы, эндокринной системы, эмоций, регулятором гомеостаза. Имея многочисленные ядра, гипоталамус связан с другими отделами мозга. Несмотря на крошечные размеры (гипоталамус человека весит менее 4 г), он участвует в регуляции всех сфер жизнедеятельности организма. Физиологи рассматривают гипоталамус как сложный «коммутатор», соединяющий нервную и эндокринную системы (Карш Ф., 1987). Наличие в гипоталамусе нейросекреторных клеток является общей чертой для всех классов позвоночных.
Передний мозг (telencephalon) у позвоночных состоит из двух полушарий. Эволюция переднего мозга производит наибольшее впечатление в сравнительно-анатомических исследованиях. Возникнув исключительно в связи с обонятельной рецепцией, у представителей высокоорганизованных классов он становится основным ассоциативным центром, от которого и зависят когнитивные способности (Ромер А., Парсонс Т., 1992). В процессе эволюции это сопровождается прогрессирующей концентрацией тел нейронов (серого вещества), в результате чего формируются кора и базальные ядра.
Кора полушарий впервые возникает у рептилий и состоит из трех отделов. Древняя кора (палеокортекс) включает в себя обонятельные доли. Старая кора (архикортекс) является центральной структурой лимбической системы, регулирующей эмоционально-мотивационное поведение. Новая кора (неокортекс) активно функционирует только у млекопитающих, хотя возникает в ходе эволюции уже у высших рептилий.
Появление неокортекса, возможно, было обусловлено необходимостью анализа сенсорной информации вомероназального органа. Вомероназальный орган возникает в связи с выходом позвоночных на сушу для хеморецепции в воздушной среде. После образования единой обонятельной сенсорной системы он, вероятно, реорганизуется в систему половой хеморецепции (Савельев С. В., 2005). В условиях первобытного леса ее роль оказалась весьма велика. Уже зачаточный неокортекс рептилий оказался связан почти со всеми важными центрами мозга.
У птиц доминирование сенсорных систем зрения, слуха, вестибулярного аппарата привело к редукции обонятельной и вкусовой систем, что, в свою очередь, привело к редукции коры. Наличие фрагментов архи-, палео– и неокортекса у предков птиц подтверждает такую версию. Высшую интегративную функцию в регуляции деятельности мозга у птиц приобретают производные базальных ядер – стриатум и гиперстриатум.
У млекопитающих, наоборот, неокортекс монопольно забирает высшие интегративные функции и составляет основную часть коры переднего мозга. Эволюция мозга млекопитающих – это, по сути дела, эволюция неокортекса. Особенно впечатляющих размеров неокортекс достигает у человека (рис. 9.1). Вставочные нейроны, образующие гигантское число связей, составляют 99 % всех его нейронов (Хамори Й., 1985). Именно с неокортексом связаны когнитивные достижения человека, поэтому этот раздел мозга всегда вызывал особый интерес нейрофизиологов. Базальные ядра у млекопитающих не играют ведущей роли – так, полосатое тело (аналог стриатума и гиперстриатума птиц) является центром непроизвольных стереотипных движений.
Под корой полушарий переднего мозга находится «белое вещество», состоящее из миелинизированных отростков нейронов, формирующих разнообразные пучки. Проекционные пучки связывают кору с другими отделами мозга. Ассоциативные пучки связывают различные участки одного полушария. Комиссуральные пучки связывают участки коры левого и правого полушарий. Самой крупной комиссурой является мозолистое тело, которое у человека включает в себя более 200 млн волокон.
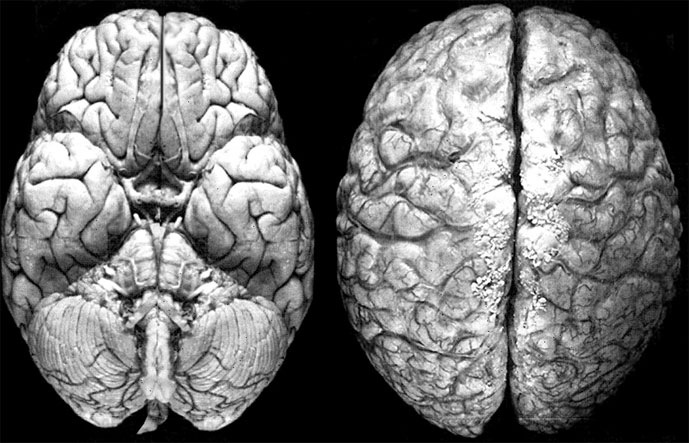
Рис. 9.1. Мозг человека

Рис. 9.2. Дельфин
Сложная система проводников, соединяющая разные части коры, наводит на мысль, что кора представляет собой единое образование. Вместе с тем, давно показано, что определенные участки коры связаны с выполнением определенных функций. Эта проблема остается одной из сложнейших в нейрофизиологии.
Среди современных млекопитающих выделяют несколько вариантов строения переднего мозга, который часто характеризуется бороздами и извилинами. Так, китообразные (отр. Cetacea) имеют своеобразную схему распределения сенсорных зон в неокортексе, высокий индекс цефализации (отношение массы мозга к массе тела), но микроструктура коры демонстрирует примитивный уровень организации. При этом дельфины (рис. 9.2), в свою очередь, имеют весьма специфичную структуру мозга и сложную систему ассоциативных связей внутри коры, обусловливающую разнообразные формы поведения.
Поскольку мозг – это весьма энергоемкая структура, существуют энергетические ограничения его эволюции. Для создания «интеллектуального» мозга необходим организм с достаточно большой массой тела: тогда относительные расходы на содержание мозга уменьшатся (Савельев С. В., 2005). Но у крупных организмов обычно большая продолжительность жизни и малое число потомков, поэтому их эволюция должна протекать весьма медленно. Исключительно высокая скорость эволюции мозга млекопитающих, и особенно человека, остается одной из загадок эволюционной физиологии. Вероятно, причиной этому послужили изменения в регуляторных генах. С ними связано резкое увеличение размеров неокортекса у приматов и еще более резкое – у человека.
Эволюционная генетика наглядно показала, что усложнение живых систем, в первую очередь, определяется совершенствованием процессов регуляции экспрессии генов, а не их количеством. Более того, в эволюции человека был использован вариант в противовес общей эволюционной тенденции. Недавние сенсационные находки в Индонезии нового вида карликовых людей, живших 20–90 тыс. лет назад, демонстрируют путь миниатюризации мозга и усложнения его организации. Этот вид, который получил название Homo floresiensis, ведет независимое происхождение от архантропов. Мы наблюдаем резкое уменьшение размеров тела и мозга с одновременным возрастанием интеллекта. Рост Homo floresiensis не превышал 1 м, а объем мозга был как у шимпанзе – 380 см3, но это не мешало им создавать орудия, даже более прогрессивные, чем у соседних Homo sapiens (Мосевицкий М. И., 2008).
Однако самый совершенный мозг не способен самостоятельно воспринимать сигналы внешнего мира (Хамори Й., 1985). Эту роль выполняет особый отдел – сенсорные системы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
МОЗГ В КОРОБКЕ
МОЗГ В КОРОБКЕ Молодой энергичный швед Эрик Стеншё не спешил строить свои гипотезы (успеется!) и не жаждал проверять чужие (и так работы невпроворот!). Он просто работал с панцирными рыбами. Материала у него хватало. Скандинавские геологи интенсивно изучали Арктику.
Головной мозг человека
Головной мозг человека Головной мозг, передний отдел центральной нервной системы позвоночных животных и человека, помещающийся в полости черепа. Г.м. – материальный субстрат высшей нервной деятельности и главный регулятор всех жизненных функций организма.У
Влюбленный мозг
Влюбленный мозг «В структуру человеческой личности вплетено очень много легковоспламеняющегося материала, и хотя эта часть может до поры до времени дремать… но если к ней поднести факел, спрятанное внутри тебя тут же вспыхнет обжигающим пламенем», — так писал Джордж
Вакцинация и ее влияние на головной мозг
Вакцинация и ее влияние на головной мозг В книге Харриса Култера «Vaccination, Social Violence and Criminality» (см. «Дополнение») рассматривается теория о том, что вакцинация может быть причиной изменения психологии и поведения человека. По моему мнению, следует обратить внимание на
У каких животных самый большой головной мозг?
У каких животных самый большой головной мозг? Самый большой головной мозг у финвала (сельдяного кита), его масса составляет 6–7 килограммов. Для сравнения: масса головного мозга индийского слона 4–5 килограммов, человека – 1,4 килограмма, лошади – 500 граммов, гориллы – 430
10.2. Мозг и психика
10.2. Мозг и психика Как связаны живая материальная система и психика? Все ли живое обладает психикой? Является ли наличие нервной системы обязательным условием проявления психического? Если да, то можно ли описать психические явления физиологическими терминами? Можно ли
ЕСТЕСТВОЗНАНИЕ И МОЗГ [ 19 ]
ЕСТЕСТВОЗНАНИЕ И МОЗГ[ 19 ] Можно с полным правом сказать, что неудержимый со времен Галилея ход естествознания впервые заметно приостанавливается перед высшим отделом мозга, или, общее говоря, перед органом сложнейших отношений животных к внешнему миру. И казалось, что
Психика и мозг
Психика и мозг Прежде чем мы начнем разбираться в том, как повреждения мозга могут сказываться на нашем восприятии окружающего мира, нужно немного подробнее рассмотреть связь между нашей психикой и мозгом. Эта связь должна быть тесной. Как мы узнали из пролога, всякий
Наш неадекватный мозг
Наш неадекватный мозг До открытия слепоты к изменениям любимым фокусом психологов были зрительные иллюзии (обманы зрения). Они тоже позволяют без труда демонстрировать, что мы видим не всегда то, что есть на самом деле. Большинство подобных иллюзий известны психологам
Наш креативный мозг
Наш креативный мозг Путаница чувствЯ знаю нескольких людей, которые выглядят совершенно нормальными. Но они видят мир, непохожий на тот, что вижу я. Будучи синестеткой, я живу в ином мире, нежели окружающие, – в мире, где больше цветов, форм и ощущений. В моей вселенной
Наш мозг справляется и без нас
Наш мозг справляется и без нас В эксперименте Либета мы как будто отстаем от того, что делает наш собственный мозг. Но в итоге мы все же нагоняем его. В других экспериментах наш мозг управляет нашими действиями так, что мы об этом даже не знаем. Так происходит, например, при
Эпилог: Я и мой мозг
Эпилог: Я и мой мозг Мы встроены во внутренний мир других людей точно так же, как мы встроены в окружающий материальный мир. Все, что мы делаем и думаем в настоящий момент, во многом определяется людьми, с которыми мы взаимодействуем. Но мы воспринимаем самих себя иначе. Мы
Что делает мозг?
Что делает мозг? На минуту приостановите чтение и составьте список действий, контролируемых вашим мозгом в данный момент. Лучше запишите их на листе бумаги, так как запоминание длинного перечня — не из тех процедур, которые наш мозг выполняет с легкостью. Когда вы
Что такое мозг?
Что такое мозг? Итак, мозг заботится о том, чтобы мы чувствовали и двигались, осуществляет внутреннюю регуляцию, обеспечивает продолжение рода и адаптацию. Если вы когда-нибудь изучали биологию, то должны помнить, что эти свойства характерны для всех животных. Даже
Мозг в действии
Мозг в действии Исследования активности мозга методами ПЭТ, ОФЭКТ и ФМРТ дают нам достаточно детальную картину специфических функций отдельных участков мозга. Мы можем узнать, какие отделы мозга связаны с теми или иными из пяти видов ощущений, какие участки