Видообразование вокруг нас
Видообразование вокруг нас
В современной биосфере полным-полно примеров видообразования, происходящего «здесь и сейчас». Все эти бесчисленные спорные виды, полувиды, супервиды, кольцевые виды, видовые комплексы и «букеты видов» (species flocks) — не что иное, как примеры незавершенного (или завершенного недавно) видообразования, в ряде случаев — очень быстрого.
Эволюция, как мы помним, процесс медленный. Но это в среднем. Темпы эволюции крайне неравномерны: вид может существовать, почти не меняясь, сотни тысяч и миллионы лет, а потом очень быстро — за считаные тысячелетия или даже века — сильно измениться или разделиться на несколько видов. Неудивительно, что те виды, которые находятся в фазе быстрых перемен именно в наши дни, привлекают особое внимание биологов.
Один из таких видов — трехиглая колюшка, небольшая рыбка, обычная в морях, реках и озерах Северного полушария. Стимулом для быстрой эволюции у колюшки стало освоение этой исходно морской рыбой пресных водоемов, что, в свою очередь, было связано с отступлением ледников около 10 тыс. лет назад и образованием на их месте множества больших и малых озер.
Эволюционные изменения у колюшки дальше всего зашли в семи озерах у тихоокеанского побережья Британской Колумбии (Юго-Западная Канада), где эти рыбки фактически разделились на два вида: придонный и пелагический (живущий в толще воды). Первый вид питается донными беспозвоночными, второй охотится на планктонных беспозвоночных в верхних слоях воды. Виды различаются не только диетой и образом жизни, но и размерами (придонный вид крупнее), формой тела, деталями строения, а также — что особенно важно — брачным поведением. Виды практически не скрещиваются между собой в природе, и даже если посадить в аквариум двух разнополых представителей придонного и пелагического видов, не оставив им возможности выбирать партнера, они, скорее всего, не станут скрещиваться (Rundle et al., 2000).
Зато пелагические колюшки из разных озер охотно скрещиваются друг с другом, так же как и придонные. И это несмотря на то, что разделение исходного вида на два произошло в семи озерах параллельно, т. е. независимо! (Здесь трудно не вспомнить аналогичные результаты, полученные в экспериментах по искусственному видообразованию.) В некоторых других американских озерах подразделение на придонный и пелагический виды наметилось, но не зашло так далеко. Во многих озерах подразделения вовсе нет: там живет исходная «генерализованная» форма колюшки, которая охотится как у дна, так и в толще воды.
Перед нами — типичный случай симпатрического видообразования. В данном случае движущим фактором видообразования было разделение исходной ниши и экологическая специализация.
О влиянии экологических факторов на видообразование мы знаем не так уж мало. Например, мы знаем, что видообразование во многом определяется и направляется набором доступных ниш. Что касается колюшек, то их разделению на два вида способствуют глубина озера и разнообразие условий в нем (например, одновременное наличие обширных отмелей и обрывистых берегов, под которыми сразу глубоко). Гораздо меньше ясности в вопросе о том, существует ли обратное влияние видообразования на свойства экосистемы. Теоретически такое влияние должно существовать. Например, модели предсказывают, что два специализированных хищника должны влиять на численность жертв иначе, чем один неспециализированный, «обобщенный» хищник. Однако проверить эти теории трудно. Колюшки — удобный объект для таких исследований, чем и воспользовались недавно биологи из США и Канады (Harmon et al., 2009).
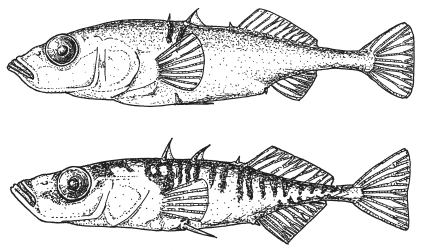
Придонная (вверху) и пелагическая (внизу) формы трехиглой колюшки.
Эксперименты проводились в цилиндрических аквариумах диаметром 2 м и глубиной 1 м. Каждый аквариум представлял собой маленькую экосистему. Сначала в него наливали чистую воду и насыпали на дно песок. В воду добавляли немного азотных и фосфорных удобрений, чтобы сделать возможным развитие фитопланктона. Затем в каждый аквариум добавляли по 2,5 л донного материала из двух озер, в одном из которых колюшка подразделилась на два вида, а в другом нет. Этот ил со дна озер содержал в себе яйца и споры всевозможных мелких озерных обитателей. Через неделю, когда в аквариуме уже вовсю кипела жизнь, туда выпускали колюшек.
В первые десять аквариумов посадили колюшек исходного вида, т. е. «генералистов», не подразделенных на пелагическую и бентосную формы (опыт 1). В две другие группы по десять аквариумов посадили придонных и пелагических колюшек (опыты 2 и 3). Наконец, в последние десять резервуаров выпустили представителей придонного и пелагического видов вместе (опыт 4). Рыбок мерили не штуками, а граммами: в каждый аквариум сажали от 5 до 6 г колюшек. Поскольку три формы колюшек различаются по размеру, это соответствовало разному количеству рыб (от трех массивных придонных рыб до шести худосочных пелагических). Затем в течение десяти недель ученые следили за состоянием «экосистем».
Сравнивая результаты опытов 1 и 4, исследователи намеревались оценить влияние диверсификации колюшек на экосистему. Опыты 2 и 3 нужны были для того, чтобы, сравнивая их с опытом 4, можно было выяснить, какие аспекты этого влияния объясняются разнообразием (числом видов) колюшек, а какие — их поведением и экологической специализацией.
Оказалось, что и разнообразие колюшек, и их специализация заметно влияют на свойства экосистемы. Эффекты оказались сложными, многообразными и не всегда соответствующими тому, что ожидалось на основе теоретических моделей. Например, можно было предположить, что пелагический вид — охотник на планктонных рачков и другие мелкие организмы — будет способствовать увеличению продуктивности экосистемы (потому что производителям органики — одноклеточным водорослям — станет легче жить, если колюшки съедят тех, кто ими питается). Но это не подтвердилось: фитопланктон лучше развивался не там, где имелись пелагические колюшки, а там, где жили колюшки исходного вида. Дело в том, что зоопланктон в аквариумах был разнообразен и далеко не все его представители являются строгими вегетарианцами. Многие планктонные рачки охотятся на других рачков, тем самым способствуя росту биомассы фитопланктона. Поэтому итоговая продуктивность системы зависит не столько от эффективности выедания рачков рыбами, сколько от того, каких именно рачков они будут поедать. А в этом отношении три вида колюшек заметно отличаются друг от друга.
Кроме различий в составе фито- и зоопланктона были выявлены различия в прозрачности воды. Этот показатель связан с количеством фитопланктона и с составом растворенной органики, причем эти связи сложны и неоднозначны. Самая прозрачная вода была в опыте 1, самая мутная — в опытах 2 и 4. По расчетам авторов, диверсификация колюшек (или ее отсутствие) может объяснить до 10 % тех различий в прозрачности воды, которые наблюдаются в реальных озерах, населенных колюшками. Это существенная величина, особенно если учесть, что прозрачность воды сильно влияет на развитие фотосинтезирующих организмов в озерах, а следовательно и на экосистему в целом.
Таким образом, видообразование, происходящее в последние 10 тыс. лет у колюшек в канадских озерах, существенно влияет на озерные экосистемы. Это влияние должно сказываться и на эволюции других озерных жителей. В частности, диверсификация колюшек может способствовать видообразованию у других организмов, т. е. порождать «цепные реакции видообразования». Для проверки этого предположения необходимы дополнительные исследования.
—————
Направленность эволюции колюшек зависит от наличия убежищ
Итак, около 10 тыс. лет назад трехиглая колюшка, исходно морская рыбка, стала осваивать пресные водоемы, образовавшиеся на месте растаявших ледников. Освоение пресных вод сопровождалось морфологическими изменениями, которые происходили независимо в разных популяциях. Одним из таких изменений является облегчение костяного панциря — уменьшение числа пластин, защищающих тело с боков. У морских колюшек пластин больше, чем у пресноводных. В основе этого эволюционного изменения лежат закрепленные отбором мутации в гене Ectodysplasin, который регулирует развитие эпителиальных структур. У людей мутации этого гена являются причиной наследственного заболевания — ангидротической эктодермальной дисплазии.

В ходе приспособления к жизни в пресных водоемах у многих популяций трехиглой колюшки уменьшилось число костных пластинок на боках.
Есть три основные гипотезы о причинах уменьшения числа пластин у колюшек при переходе к жизни в пресных водах. Согласно первой, все дело в большей «дороговизне» костной ткани в пресных водоемах по сравнению с морем. В озерах и реках, где живут колюшки, меньше кальция, чем в морской воде, поэтому для того, чтобы вырастить костяной панцирь, рыбкам приходится тратить больше калорий.
Вторая гипотеза винит во всем хищных пресноводных насекомых, таких как гладыши и личинки стрекоз. Показано, что шипы и броня колюшек защищают их от хищных рыб, повышая шансы схваченной рыбки вырваться из зубов хищника. Но против насекомых эти средства защиты бессильны: шестиногим хищникам даже удобнее хватать мальков за их колючки и костяные пластинки (со взрослой рыбкой насекомые не могут справиться). Экспериментально установлено, что хищные насекомые действительно могут избирательно выедать мальков с длинными колючками и тяжелой броней (Marchinko, 2009).
Третья гипотеза предполагает, что причина редукции брони — в разной доступности укрытий. Морские колюшки плавают в толще воды, где спрятаться абсолютно негде. Остается полагаться на броню. В пресных водоемах колюшки стали держаться ближе ко дну, где укрытий достаточно. Возможно, в этой ситуации для колюшек решающее значение приобрела маневренность и способность быстро юркнуть в укрытие. Эксперименты показали, что облегченная броня действительно улучшает маневренность и способность к «быстрому старту». Однако нужно еще доказать, что при наличии укрытий колюшки с облегченным панцирем действительно получают селективное преимущество, т. е. реже поедаются хищниками.
Недавно это удалось сделать биологам из Хельсинкского университета (Leinonen, 2011). Они сажали группы колюшек с полным и неполным набором костных пластин в большие аквариумы со щуками, причем в половине аквариумов были укрытия, где колюшки могли спрятаться от хищника. Спустя три месяца подсчитывалось число выживших рыбок.
Результаты подтвердили гипотезу о роли укрытий в эволюции костной брони. В аквариумах без укрытий число пластин положительно коррелировало с выживаемостью: чем меньше у рыбки пластин, тем больше были ее шансы погибнуть в зубах щуки. В аквариумах с укрытиями все было наоборот: здесь вероятность выживания росла с уменьшением числа пластин (что объясняется ростом маневренности). Впрочем, это не исключает и две другие возможные причины редукции панциря (недостаток кальция и хищные насекомые). Ведь у этих гипотез тоже есть экспериментальные подтверждения. Вполне возможно, что все три фактора действуют вместе, что объясняет очень быструю эволюцию этого признака в некоторых водоемах.
Исследование подтвердило два важных эмпирических обобщения, которые иногда называют даже «аксиомами» эволюционной биологии: 1) полезность признака зависит от контекста: один и тот же признак может быть полезным для одних популяций и вредным для других, живущих в иных условиях; 2) эволюция — это постоянный поиск компромиссов: например, колюшки не могут быть одновременно юркими и бронированными, поэтому приходится выбирать что-то одно в зависимости от обстоятельств.
Завершая рассказ о колюшках, нельзя не упомянуть о примере обратимой эволюции, который эти рыбки продемонстрировали в озере Вашингтон на северо-западе США. Лет 50 назад у тамошних колюшек было мало костных пластин. К настоящему времени число пластин значительно выросло: колюшки фактически вернулись к своему прежнему, «морскому» состоянию. Предполагаемая причина в том, что вода в озере за это время очистилась, а в прозрачной воде колюшкам труднее спрятаться от хищной форели. Поэтому тяжелая броня снова стала для них более выгодной, чем маневренность.
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Видообразование и конвергенция
Видообразование и конвергенция Теперь нам бы хотелось обсудить значение ламарковской гипотезы об обратной связи сомы и зародышевой линии для теории видообразования и конвергенции. Помимо катастрофического исчезновения видов, например, из-за падения метеоритов или
Затерянные миры вокруг нас
Затерянные миры вокруг нас Открытие затерянного мираВ 1937 году лётчик Джим Энджел, потерпев аварию, приземлился на одном из плоскогорий в глубине девственного леса Южной Америки. Ауян-Тепюи, Гора Дьявола, так называлось это гиблое плато. Он посадил самолёт над болотом, у
Часть вторая. Микробы вокруг нас
Часть вторая. Микробы вокруг нас Меня посетили благородные дамы, которым очень хотелось посмотреть на маленьких змеек в капле уксуса. Однако некоторым из них эти змейки показались настолько отталкивающими, что они поклялись никогда больше не брать уксуса в рот. Но что бы
Видообразование на разных островах идет параллельными путями
Видообразование на разных островах идет параллельными путями Уединенные острова и озера представляют собой уникальные эволюционные лаборатории, где небольшие популяции животных и растений могут сильно меняться за весьма малое по геологическим масштабам время. Этому
Видообразование — личное дело каждого
Видообразование — личное дело каждого Симпатрическое видообразование, по идее, должно начинаться с того, что разные особи одного и того же вида начинают приспосабливаться к разным экологическим нишам в пределах общего ареала. Приспособление должно быть сопряжено с
Эволюция цветного зрения и видообразование
Эволюция цветного зрения и видообразование Зрение у цихлид играет важную роль при выборе партнера: самцы каждого вида имеют свой особый наряд, а самки превосходно разбираются в нюансах их окраски. Зрение может быть специально настроено на спектральные характеристики
Жизнь вокруг очага[38]
Жизнь вокруг очага[38] Гешер Бенот Яаков — знаменитая ашельская стоянка на территории Израиля в северной части рифта Мертвого моря. Люди («гейдельбергские в широком смысле») жили там 750–790 тыс. лет назад. Это была болотистая местность на берегу древнего озера. Уже многие
АЛЛОПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
АЛЛОПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ Способ возникновения новых видов зависит от географического расположения. Большинство видов появляется в результате аллопатрического видообразования (алло — другой, патрис — родина), то есть в результате географического разделения
СИМПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
СИМПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ Симпатрическое видообразование происходит внутри одной популяции (сим — вместе, патрис — родина). Оно менее распространено, чем аллопатрическое. Этим термином обозначается процесс образования разных видов внутри постоянно
5.3. Видообразование
5.3. Видообразование Процесс образования новых видов, происходящий на основе преобразований популяций, называется видообразованием. Понятно, что невозможно изучать видообразование, не имея определения вида и критериев его выделения.Вид – это фундаментальное понятие в
Глава 1. ЗАТЕРЯННЫЕ МИРЫ — ВОКРУГ НАС!
Глава 1. ЗАТЕРЯННЫЕ МИРЫ — ВОКРУГ НАС! Весной 1948 года в пустыне штата Нью-Мексико упал и разбился таинственный летательный аппарат. Из-под его обломков были извлечены обгоревшие останки нескольких маленьких человекообразных существ, чей рост не превышал 90 сантиметров.
Чудеса вокруг магнита
Чудеса вокруг магнита До сих пор не припомню, зачем я поставил черную подковку магнита на стеклянную лабораторную посудину, где содержались крохотные насекомые — мелиттобии. Может быть, детская привычка? Мальчишкой я был убежден, что любое великое открытие сделать очень
11. Видообразование как результат эволюции
11. Видообразование как результат эволюции Вспомните!Что такое вид?Какие виды древних растений и животных вам известны?Какую роль играет изоляция в процессе эволюции?Видообразование – это процесс возникновения новых видов. В настоящее время на земном шаре обитает
Вода вокруг нас
Вода вокруг нас Громадна водная поверхность Земли. Свое название — океан — она получила по имени одного из титанов древнегреческой мифологии.«Вода — краса всей природы», — говорил писатель С. Т. Аксаков. Вода окружает нас везде и всюду, и без воды жизнь невозможна. В
Глава первая Мир вокруг нас
Глава первая Мир вокруг нас Чудеса далекие и чудеса близкиеВы хотите путешествовать?Конечно, хотите!Кто в детстве и юности не мечтает о далеких, полных приключений и опасностей путешествиях?Ведь и вы, читатель, наверное, мечтаете о том же.Что же, давайте мечтать вместе. А