Динозавры осваивают воздух
Динозавры осваивают воздух
Современный мир изобилует летающими существами — насекомые, птицы, летучие мыши; есть и другие, которые хоть и не настоящие летуны, но уже и не совсем наземные жители — древесные лягушки, белки, шерстокрылы, ящерицы — «летучие драконы». Существует даже аэропланктон — мельчайшие животные и бактерии, приспособленные переноситься на большие расстояния воздушными массами. Человек тоже от них не отстает — поднимается в воздух при помощи технических средств. Таким образом, у многих групп животных прослеживается тенденция к освоению воздушного пространства.
Полет требует согласованных и множественных перестроек тела. Чем они сбалансированнее и глубже, тем искуснее получившийся летун. В этом разделе мы разберем, как полетели птицы, потому что их эволюционная история замечательно прояснилась в последние годы благодаря серии блестящих находок различных переходных форм.
Вскоре после публикации «Происхождения видов» был обнаружен первый скелет археоптерикса — знаменитой промежуточной формы между рептилиями и птицами. Он был захоронен в позднеюрских отложениях Германии, возраст которых, по последним данным, чуть менее 150 млн лет. У археоптерикса было развито оперение (типичная птичья черта), а по строению скелета он мало отличался от мелких динозавров-теропод. У него были когти на передних конечностях, зубы и длинный хвост. Характерных «птичьих» особенностей скелета было немного (крючковидные отростки на ребрах, вилочка). Археоптерикс — классический пример переходной формы. Но это ископаемое было лишь одиноким указателем на длинной дороге, ведущей к птицам. Ясно, что при постепенном становлении птичьих черт подобные промежуточные формы были гораздо разнообразнее. Но вот сохранились ли они? Удастся ли палеонтологам их обнаружить?
Удалось. Дальнейшие поиски в юрских и меловых слоях открыли богатую фауну примитивных переходных полудинозавров-полуптиц. Ниже показана филогенетическая схема, отражающая родственные связи большинства известных на сегодня переходных форм, сочетающих признаки птиц и динозавров.
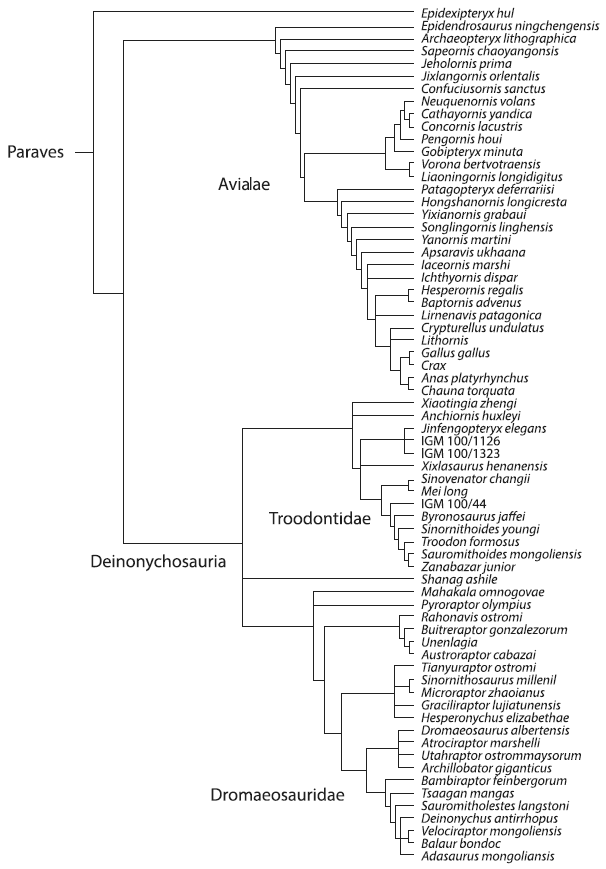
В силу правил современной кладистической систематики факт происхождения птиц от динозавров (а не от общих с динозаврами предков) требует рассматривать птиц как подгруппу динозавров, а для «собственно» динозавров теперь применяют громоздкий термин non-avian dinosaurs (нептичьи динозавры).
Началом становления птиц следует считать юрскую эпоху, когда многие хищные динозавры начали эволюционировать «в птичью сторону». Представителей той линии динозавров, в которой происходило это параллельное «оптичивание», объединяют под общим назанием Paraves. Сами современные птицы на схеме представлены четырьмя видами — курицей (Gallus) и гокко (Crax), уткой (Anas) и паламедеей (Chauna). Они занимают скромное место на одной из веточек среди множества ископаемых видов, формируя вместе с ними обширную группу Avialae, «сестринскую» по отношению к другой большой кладе оперенных динозавров — дейнонихозаврам. Дейнонихозавры подразделяются на троодонтид и дромеозаврид. Археоптерикс занял на схеме вполне логичное место одного из самых примитивных представителей всей разнообразной группы мезозойских летунов. В конце мелового периода, 65,5 млн лет назад, все они вымерли, кроме небольшой группы, которая пережила меловой кризис и дала начало современным птицам.
Самым примитивным представителем Paraves на сегодняшний день считается эпидексиптерикс. Он близок к общему предку первых летающих динозавров. Его нашли в 2008 году в юрских (от 168 до 152 млн лет назад) отложениях Внутренней Монголии (Китай). Это животное размером с голубя. По оценке исследователей, эпидексиптерикс весил примерно 160 г — меньше, чем другие птицеподобные динозавры. От своих родственников он отличался прежде всего коротким хвостом: у него всего 16 хвостовых позвонков, причем последние десять образуют нечто, напоминающее пигостиль примитивных птиц, хотя эти позвонки и не сливаются в единое целое. Длина хвоста составляет 70 % длины туловища, тогда как у ближайшего родственника — эпидендрозавра — хвост был втрое длиннее туловища.
Название «эпидексиптерикс» (от греч. epidexi — «показывать» и pteryx — «перо», «крыло») динозаврик получил неспроста. К хвостовым позвонкам прикреплялись четыре необычайно длинных пера с центральным стержнем и опахалом из неветвящихся бородок. Туловище, включая конечности, покрывали мелкие перья без стержня, похожие на пух других динозавров и примитивных птиц. Перьев, пригодных для полета, у эпидексиптерикса не было, и летать он наверняка не мог.
Эти длинные хвостовые перья служили исключительно «для красоты», т. е. для привлечения брачного партнера. Самцы многих современных птиц красуются перед самками длинными хвостовыми перьями, больше ни на что не годными и только мешающими летать. Наряд эпидексптерикса доказывает, что красота брачных нарядов была важна для «птичьей» ветви динозавров уже в самом начале их истории (Zhang et al., 2008).
Перья нужны для красоты, тепла и полета. По-видимому, сначала они выполняли две первые задачи и только позже пригодились для третьей. Да и полет начинался не сразу: маховые перья развивались постепенно. Увеличение несущей поверхности передних конечностей повышало стабильность бегущего или прыгающего динозаврика. Начальные этапы эволюции маховых перьев, возможно, были связаны со стабилизацией быстрого бега по неровной поверхности. При этом большие перья постепенно смещались на задний край конечности; строение самих перьев и мест их крепления постепенно совершенствовалось.
Оперенные примитивные предки птиц дали начало группе дромеозаврид. Эти животные распрощались с поверхностью земли, предпочитая деревья. Недавние исследования показали, что даже прославленный велоцираптор не столько бегал по равнинам, сколько лазил по деревьям, и его строение было приспособлено именно к этому образу жизни. Выяснилось, что велоцирапторы были покрыты перьями, что подразумевает теплокровность (Turner et al., 2007). Раньше их считали (эта версия увековечена фильмом «Парк юрского периода») скоростными равнинными стайными хищниками, наделенными страшным оружием — зубастыми челюстями и четырьмя гигантскими когтями (по одному на каждой из четырех конечностей). Но оказалось, что устройство и геометрия когтей на задних лапах велоцираптора сравнимы с когтями некоторых лазающих по деревьям птиц и млекопитающих. Это сравнение со всей определенностью переместило велоцираптора с земли на дерево. Велоцирапторы использовали колоссальные когти на ногах не только для того, чтобы убивать толстокожую жертву. При лазании по стволу они опирались на этот коготь. Он должен был быть крупным, чтобы удерживать на вертикальном стволе довольно массивное животное — эти динозавры весили около 15 кг.
Скорее всего, у велоцирапторов был «птичий» механизм фиксации когтей. Когда птица сидит на ветке, ее когти обхватывают ветку и фиксируются в этом положении без затрат энергии. Это позволяет птице спать сидя на дереве и при этом не падать. Опорой для древесного хищника при посадке на стволе также служил жесткий хвост, составленный сросшимися позвонками и окостеневшими сухожилиями. Перья, древесный образ жизни, да еще и в стае — ну чем не птичка!
Анхиорнис (Anchiornis huxleyi) принадлежит к другой части филогенетического дерева — к базальным (т. е. примитивным, ранним) троодонтидам. Анхиорнис жил раньше археоптерикса — 155–151 млн лет назад. Когда его нашли и описали (а это произошло в 2009 году), он сразу занял почетное место среди самых удивительных и неожиданных животных. Это летающее создание величиной с ворону имело контурные маховые перья и на передних, и на задних конечностях. Превосходные отпечатки безошибочно передают строение скелета и перьев, так что в интерпретациях нет сомнений — на ногах у анхиорниса были крупные перья, пригодные для полета.
Это не первый найденный четырехкрылый динозавр — такие были известны и раньше, например микрораптор, живший 125 млн лет назад. Поначалу многие эксперты трактовали четырехкрылость как некий курьез, экстравагантный, но незначительный поворот эволюции Paraves.
В 2005 году был описан еще один динозавр с крупными перьями на ногах — педопенна (что переводится как «пероног»), жившая одновременно с археоптериксом или, возможно, чуть раньше. Она известна только по остаткам оперенных задних конечностей. Педопенну интерпретировали как базального представителя Paraves (на схеме она не показана).
Судя по распределению четырехкрылых форм по эволюционному дереву, четырехкрылость была свойственна первым представителям всех трех ветвей Paraves: троодонтидам, дромеозавридам и Avialae. Из этого следует, что современные птицы произошли от четырехкрылых предков. Между прочим, у примитивных Avialae, включая археоптерикса, действительно были на ногах контурные перья, хотя и не такие крупные.
В отличие от тех современных птиц, у которых имеется более или менее развитое оперение на ногах, у микрораптора, педопенны и анхиорниса контурные перья задних конечностей располагались упорядоченно и образовывали большие плоские поверхности, которые, несомненно, влияли на аэродинамические свойства животного. Как именно использовались эти дополнительные плоскости — для планирования, маневрирования или, допустим, торможения, — точно не известно. Едва ли четырехкрылые динозавры могли активно махать своими «задними крыльями» — скелет их задних конечностей не приспособлен для таких движений, — но они могли разворачивать ноги таким образом, чтобы перья помогали планировать. Четырехкрылый полет в любом случае не слишком эффективен.
Анхиорнис, судя по скелету, был не очень хорошим летуном (микрораптор летал получше). Задние конечности анхиорниса, если забыть о перьях, больше похожи на ноги профессионального бегуна. Но крупные перья на ногах едва ли помогали ему быстро бегать — скорее, наоборот, должны были мешать. У современных быстро бегающих птиц прослеживается тенденция к утрате оперения на ногах. Может быть, анхиорнис еще не успел «решить», что для него важнее — летать или бегать, и строение его задних ног является результатом компромисса между разнонаправленными векторами отбора. В ходе дальнейшей эволюции ранних птиц крылья и ноги более четко поделили функции: первые для полета, вторые для бега.
Анхиорнис по многим чертам скелета похож на троодонтид, к которым его и отнесли, но некоторые признаки (например, очень длинные передние конечности) сближают его с дромеозавридами и Avialae. В результате формальные диагнозы (списки отличительных признаков) трех групп Paraves становятся более расплывчатыми. Это типичная ситуация, неприятная для систематиков, но абсолютно неизбежная при изучении базальных представителей крупных таксонов (об этом мы говорили при обсуждении кембрийских типов). Понятно, что чем ближе мы подбираемся к общему предку всех Paraves, тем менее четкими становятся различия между тремя эволюционными линиями, входящими в состав этой группы. «Переходные формы» размывают эти различия. Дарвин был прав, рассуждая о стирании границ между таксономическими группировками.
Кроме того, мозаика птичьих и динозавровых черт, столь широко представленная у Paraves, еще раз напоминает о параллельной эволюции. Начало эволюции каждой крупной группы, как правило, представлено несколькими линиями, в каждой из которых своим чередом, со своими особенностями и со своей скоростью шло приспособление к новой среде или новому образу жизни. В случае «оптичивания» динозавров это линии дромеозаврид, троодонтид и Avialae.
—————
Птицы, динозавры и их гены
Признать, что птицы являются динозаврами — а именно этого требует современная систематика, — психологически довольно трудно. К тому же есть и «нединозавровые» гипотезы происхождения птиц. Правда, к сегодняшнему дню у них почти не осталось сторонников (об одной из них мы рассказали в книге «Рождение сложности»). Мог бы помочь генетический анализ, но ДНК динозавров не сохранилась (по крайней мере, пока не выделена и не прочитана). Но все же подобраться к генам динозавров ученые смогли (Organ et al., 2009). Они косвенным образом оценили размер геномов различных динозавров. Для этого они воспользовались тем обстоятельством, что в ископаемых костях, если они достаточно хорошо сохранились, на срезах бывают видны маленькие полости, в которых при жизни животного помещались клетки костной ткани — остеоциты. Известно, что размер генома во многих группах живых существ положительно коррелирует с размером клеток. Срезы костей 26 видов современных четвероногих, взятых для исследования, обнаружили линейную зависимость между размером генома и средним объемом остеоцита. Найденная зависимость позволила ученым с приемлемой точностью оценить размеры геномов 31 вида динозавров и ископаемых птиц.
У современных птиц размер генома необычно мал — от 0,97 до 2,16 млрд пар нуклеотидов, в среднем 1,45. Для сравнения, у жабы 6,00, крокодила 3,21, коровы 3,7, кошки 2,9, мыши 3,3, человека 3,5. А что же динозавры? Геномы птицетазовых динозавров (вопреки своему названию не родственных птичьей ветви), имели средний размер около 2,5 млрд пар нуклеотидов, что сопоставимо с современными рептилиями. Геномы теропод (хищных двуногих динозавров), в том числе самых древних, живших задолго до появления птиц, были значительно меньше — в среднем 1,78 млрд п. н. Из девяти исследованных видов теропод только у одного (овираптора) размер генома оказался за пределами диапазона, характерного для современных птиц.
Таким образом, общий предок всех динозавров, скорее всего, имел типичный для наземных позвоночных большой геном. Это состояние сохранилось у птицетазовых динозавров, а также у рептилий, доживших до наших дней. У ящеротазовых динозавров (к которым относятся тероподы) еще на заре их истории (в триасе) произошло радикальное сокращение генома. Птицы, таким образом, унаследовали маленький геном от своих предков — динозавров-теропод, а не приобрели его позже как адаптацию к полету.
Тем не менее между размером генома и полетом все-таки есть связь. Об этом свидетельствуют два обстоятельства. Во-первых, у нелетающих птиц (таких как страусы) геном больше, чем у летающих. Во-вторых, у летучих мышей геном меньше, чем у других млекопитающих.
Еще одно генетическое исследование, пролившее свет на раннюю эволюцию птиц, было выполнено в 2011 году.
С точки зрения эмбриологии и сравнительной анатомии три пальца крыла у птиц соответствуют пальцам II, III и IV исходной пятипалой конечности. Это противоречит палеонтологическим данным: ископаемые находки демонстрируют постепенную утрату пальцев IV и V в эволюционном ряду, ведущем от первых триасовых динозавров к археоптериксу и другим вымершим ящерохвостым птицам. Три оставшихся у них пальца — это пальцы I, II и III. Значит, если современные (веерохвостые) птицы являются потомками ящерохвостых, то пальцы их крыльев должны быть пальцами I–II–III, а не II–III–IV. Получается, пальцы — всего лишь пальцы! — не соглашаются происходить от динозавров. И этого достаточно, чтобы заронить сомнения в справедливости динозавровой теории. Но стараниями генетиков этот конфликт между эмбриологией и палеонтологией был разрешен (Wang et al., 2011).
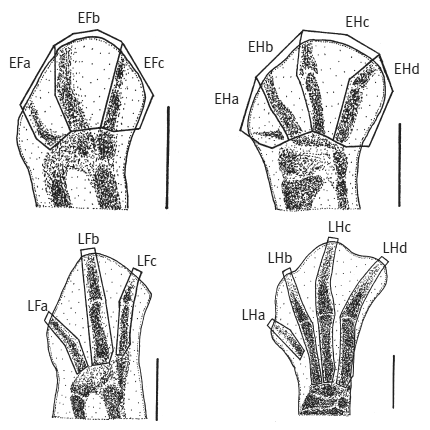
Зачатки крыльев (слева) и ног (справа) цыпленка на ранней (вверху, E) и более поздней (внизу, L) стадиях развития. Длина масштабной линейки — 1мм. Пальцы нумеруются слева направо. Видно, что по своему положению в зачатках конечностей три пальца крыла (Fa, Fb, Fc) соответствуют пальцам ноги II, III и IV (Hb, Hc, Hd). Однако набор генов, работающих в зачатке первого пальца крыла (Fa), соответствует первому пальцу ноги (Ha), а не второму (Hb). Из Wang et al., 2011.
Они измерили активность генов в зачатках пальцев передних и задних конечностей куриного эмбриона. Всего в зачатках пальцев удалось зарегистрировать активность 14 692 генов. Анализ проводился по отдельности для каждого пальца на двух стадиях развития зачатков (ранней и поздней). Выяснилось, что по характеру экспрессии генов первые (самые внутренние) пальцы крыльев и ног четко отличаются от остальных (внешних) пальцев, но при этом они похожи друг на друга. Первые пальцы отличаются от других по уровню экспрессии 556 генов. Это весомый аргумент в пользу того, что первый палец крыла — это на самом деле палец I, а не II, как следует из данных эмбриологии.
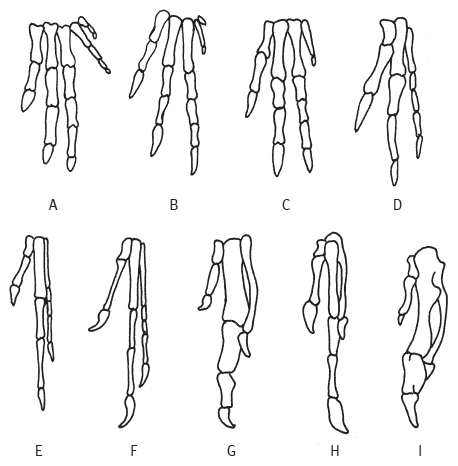
Редукция пальцев IV и V в эволюционном ряду от ранних динозавров к птицам. Пальцы нумеруются слева направо. (A — C) триасовые динозавры: (A) Heterodontosaurus (птицетазовый динозавр, не родственный птицам), (B) Herrerasaurus (ранний представитель теропод), (C) Coelophysis (более продвинутый представитель теропод). (D — E) юрские тероподы: (D) Allosaurus, (E) Ornitholestes. (F — G) ящерохвостые птицы: (F) Archaeopteryx, (G) меловой Sinornis. (H — I) современные (веерохвостые) птицы: (H) птенец гоацина (Opisthocomus), (I) курица (Gallus). Из Vargas A. O., 2005. Beyond Selection // Revista Chilena de Historia Natural. V. 78. P. 739–752.
Очевидно, именно работой этих 556 генов определяется «идентичность» первых пальцев с молекулярно-генетической точки зрения. Что касается второго и третьего пальцев крыла, то их программы развития, похоже, сформировались заново из фрагментов программ развития пальцев II, III и IV. Поэтому нельзя определенно сказать, каким пальцам ноги они гомологичны с молекулярно-генетической точки зрения. Но если учесть весь комплекс имеющихся данных (молекулярно-генетическую гомологию первых пальцев, взаимное расположение пальцев крыла, палеонтологические данные), то все-таки получается, что три пальца птичьего крыла происходят от пальцев I–II–III, а не II–III–IV. А это значит, что конфликт между данными эмбриологии и палеонтологии снят.
Полученные результаты подтвердили высказанную ранее гипотезу, согласно которой в эволюции динозавров-теропод имело место смещение областей работы программ развития пальцев I–II–III на одну эмбриональную позицию в дистальном направлении (от туловища), так что палец I стал развиваться там, где положено развиваться пальцу II (Wagner, Gauthier, 1999).
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
1. Динозавры и сородичи
1. Динозавры и сородичи
Неведомые динозавры
Неведомые динозавры Таинственная гибель динозавров порождает множество гипотез (последняя из них объясняет вымирание ящеров падением астероида, но и это опять-таки гипотеза, а не доказанный факт). Почти каждый год приносит новые открытия, в свете которых динозавры
Эректусы осваивают мир
Эректусы осваивают мир В течение второго миллионолетия до н. э. эректусы постепенно расселялись по просторам Евразии, неся с собой древнейшую технологию обработки камня — олдувайскую. Технические усовершенствования если и появлялись кое-где за пределами Африки, то
ДИНОЗАВРЫ
ДИНОЗАВРЫ Во время триасового периода (245–202 миллиона лет назад) пресмыкающиеся архозавры (правящие ящеры) эволюционировали в четыре основные группы: два отряда динозавров, птерозавров и крокодилов. Две группы динозавров (ящеротазовые и птицетазовые) были в не более
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ Птицетазовые динозавры по-научному называются Ornithischia. Форма таза у них такова, что кости ног направлены вниз, параллельно друг другу. Все они были растительноядными и в эпоху распространения этих динозавров — в юрский и меловый периоды (202—65
ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ Ящеротазовые динозавры (Saurischia), появившиеся ранее другой группы, имели строение таза, схожее с другими пресмыкающимися. Две кости ног расходились у них в разных направлениях. Некоторые из них были растительноядными, другие — хищными. Они часто
Воздух, которым мы дышим
Воздух, которым мы дышим Чистый свежий воздух питает легкие и очищает душу — подобно тому как полноценное питание обеспечивает жизненной энергией тело (не случайно слова «душа» и «дыхание» происходят во всех языках от одного корня). Однако в настоящее время чистый
Воздух!
Воздух! Отпечатки дисков, полос и ветвей на 600-миллионнолетних камнях кажутся малопримечательной группой окаменелостей, но это впечатление обманчиво. Это следы, оставленные настоящими революционерами: организмами совершенно нового типа, прежде невиданными. Это первые
Почему вымерли динозавры?
Почему вымерли динозавры? Согласно теории, имеющей в настоящее время наибольшее количество сторонников, 65 миллионов лет назад на Землю упал астероид диаметром около 10 километров. Установлено даже место его падения – полуостров Юкатан в Мексике. Энергия, выделившаяся
Что весит больше – пища, потребляемая человеком за сутки, или воздух, который он вдыхает за этот же период времени?
Что весит больше – пища, потребляемая человеком за сутки, или воздух, который он вдыхает за этот же период времени? Вес воздуха, вдыхаемого и выдыхаемого человеком в течение суток, подсчитать несложно. При каждом вдохе человек вводит в свои легкие около полулитра воздуха.
4.2.1. Воздух как экологический фактор для наземных организмов
4.2.1. Воздух как экологический фактор для наземных организмов Низкая плотность воздуха определяет его малую подъемную силу и незначительную спорность. Обитатели воздушной среды должны обладать собственной опорной системой, поддерживающей тело: растения –
ВОЗДУХ И КРЫЛЬЯ
ВОЗДУХ И КРЫЛЬЯ ПРИКРЫТОЕ сверху сплошным, без единой щелочки, панцирем, живет в земле своей невидимой жизнью гнездо.Чаще всего где-то глубоко под сводами купола, отделенная от него целыми лабиринтами лазеек и переходов, связывающих этажи, лежит небольшая, плоская, почти
Динозавры, или дивоящеры
Динозавры, или дивоящеры Потомки архозавров особенно размножились во второй половине мезозойской эры. Они отличались исключительным разнообразием. Некоторые остались хищниками с короткими передними ногами. Все они обладали толстым и очень сильным хвостом, который был
Как развивались динозавры
Как развивались динозавры Экспедиции, работавшие на раскопках в пустыне Гоби, несколько раз находили гнезда с яйцами. Высказывались предположения, что это яйца крупных черепах. Но новейшие исследования показали, что это яйца динозавров. Яйца динозавров были обнаружены в
Прыжок в воздух
Прыжок в воздух Следующую остановку я решил сделать десять миллионов лет спустя — в разгаре эоцена, второй эпохи третичного периода. Но прежде чем рассказать об удивительных приключениях, выпавших там на мою долю, я должен сделать небольшое отступление. Дело в том, что