Стресс помогает справиться с вредными мутациями
Стресс помогает справиться с вредными мутациями
Как мы уже знаем, адаптация, развившаяся для повышения устойчивости к одному типу помех, может автоматически повысить устойчивость и к другим их типам. Типичный пример — белки-шапероны, помогающие другим белкам принять правильную трехмерную конфигурацию. Эта конфигурация может быть нарушена как из-за перепадов температуры, так и из-за мутаций, поэтому шапероны повышают устойчивость одновременно к обоим видам помех.
Из этого вытекает интересное следствие. Не исключено, что активация систем, предназначенных для противодействия неблагоприятным внешним условиям, может попутно уменьшать негативные эффекты вредных мутаций. Иными словами, реакция организма на средовой стресс может сглаживать проявления наследственных дефектов. Это предположение выглядит правдоподобным, что, впрочем, никоим образом не отменяет необходимость его экспериментальной проверки, детализации и выявления конкретных механизмов.
Именно с этой целью испанские биологи провели серию экспериментов на C. elegans (Casanueva et al., 2012). Первый эксперимент был поставлен, чтобы проверить, приводит ли активация генерализованного (общего, неспецифического) ответа на стресс к сглаживанию фенотипических эффектов вредных мутаций, т. е. к снижению их пенетрантности. Для этого были использованы генетически модифицированные черви (линия +HSF-1) с повышенной продукцией «фактора теплового шока 1» (heat shock factor 1, HSF-1). Этот белок является регулятором высокого уровня: он активирует целый ряд генов, участвующих в ответе на средовой стресс, в том числе гены шаперонов. Модифицированные черви постоянно находятся в состоянии «повышенной боевой готовности» и отличаются от обычных червей высокой устойчивостью к неблагоприятным воздействиям.
Исследователи скрестили червей +HSF-1 с носителями 11 разных вредных мутаций и сравнили пенетрантность этих мутаций у особей с обычным и искусственно повышенным уровнем HSF-1. Оказалось, что гиперактивность системы ответа на средовой стресс достоверно снижает пенетрантность восьми из 11 мутаций. Процент особей, у которых эти мутации проявились в фенотипе, оказался понижен у червей +HSF-1 по сравнению с контролем на 18–88 %.
Более того, оказалось, что эффекты мутаций, пенетрантность которых снизилась у червей +HSF-1, зависят также и от температуры. Если подвергнуть молодую личинку обычного (немодифицированного) червя тепловому шоку (нагреванию до 35 °C на два часа), пенетрантность мутаций снижается примерно так же, как у червей +HSF-1. Если же пенетратность мутации не зависит от уровня HSF-1, то и тепловой шок на нее не влияет. Скорее всего, те три мутации, которые оказались нечувствительны к уровню HSF-1, просто «не лечатся» шаперонами (например, они могут быть связаны не с нарушением структуры какого-нибудь белка, а с полным его отсутствием).
Таким образом, эксперимент показал, что реакция на стресс, вызванная путем генетических модификаций или средового воздействия (теплового шока), действительно снижает пенетрантность вредных мутаций. Проявится или нет мутация в фенотипе, зависит, во-первых, от генетики (например, от генетически обусловленного базового уровня HSF-1), во-вторых, от жизненной истории особи, в том числе от стрессов, перенесенных в детстве. Главное тут — соблюсти меру: слишком сильный стресс может повредить систему «забуферивания» мутаций при помощи шаперонов и тем самым усугубить фенотипические эффекты этих мутаций (Queitsch et al., 2002), однако умеренный стресс, перенесенный в определенном возрасте, напротив, может их сгладить.
Как мы уже говорили, неполная пенетрантность наблюдается и в изогенных популяциях, даже если условия, в которых развиваются особи, одинаковы. В ходе дальнейших экспериментов авторы, работая с изогенными линиями C. elegans, попытались выяснить, чем вызывается эта странная вариабельность у животных с одинаковыми генами и «жизненным опытом». Черви C. elegans способны к самооплодотворению (см. главу 3), поэтому вывести изогенную линию нетрудно. Более того, такие линии нередко возникают и в природе.
Ранее уже было показано, что в изогенных линиях C. elegans наблюдается случайная (не связанная ни с генетическими, ни со средовыми факторами) вариабельность по уровню экспрессии некоторых генов — участников реакции на стресс, причем от этого уровня зависит, во-первых, устойчивость червей к перегреву, во-вторых — продолжительность их жизни: особи с высоким уровнем шаперонов живут дольше. Речь идет, напомним, о генетически идентичных особях (Rea et al., 2005). Поэтому авторы вполне логично предположили, что случайные различия в уровне экспрессии шаперонов и других компонентов реакции на стресс могут влиять также и на фенотипический эффект вредных мутаций.
Для проверки этого предположения генетически идентичных червей с той или иной вредной мутацией подвергали кратковременному перегреву на стадии личинки. Затем их делили на две группы в зависимости от силы реакции на стресс, которую определяли по уровню экспрессии участника этой реакции — гена hsp-16.2. Оказалось, что у червей с сильной реакцией на стресс вредные мутации проявляются в фенотипе реже, чем у особей, слабо реагирующих на перегрев. Сама же сила реакции на стресс в свою очередь зависит от случайных различий в уровне экспрессии генов теплового шока. Такие различия есть у генетически идентичных личинок еще до того, как их подвергнут перегреву. У кого эти гены изначально работают активнее, тот и выдает более сильную реакцию на перегрев.
Таким образом, пенетрантность мутаций зависит не только от уровня реакции на стресс, обусловленного генетикой (как у червей +HSF-1) или средой, но и от случайных колебаний этого уровня, не зависящих ни от генов, ни от среды. У генетически идентичных червей имеется случайная изменчивость по силе реакции на стресс, причем те особи, которые реагируют на стресс активнее, получают сразу три важных преимущества. Во-первых, они легче переносят перегрев, во-вторых — дольше живут, в-третьих — они лучше защищены от вредных мутаций.
Возникает естественный вопрос: если сильная реакция на стресс так полезна, почему этот признак остается нестабильным, почему он варьирует даже у генетически идентичных особей? Казалось бы, отбор давно должен был закрепить в генофонде C. elegans какие-нибудь мутации, обеспечивающие стабильно высокий уровень активности «стрессовых» генов.
Исследователи предположили, что раз этого не произошло, значит, тут есть какой-то подвох. Возможно, высокая активность этих генов не дается даром и, кроме пользы, приносит также и вред. Дальнейшие эксперименты подтвердили это предположение. Оказалось, что черви с более интенсивной реакцией на тепловой шок отличаются пониженной плодовитостью. За долгую жизнь и устойчивость к перегреву и вредным мутациям червям приходится платить низкой эффективностью размножения.
Черви C. elegans в природе живут в гниющих растительных остатках и питаются бактериями. Условия в местах их обитания переменчивы и непредсказуемы. Пока условия остаются благоприятными, особи с пониженным уровнем экспрессии «стрессовых» генов оказываются в выигрыше благодаря высокой скорости размножения. При ухудшении условий преимущество оказывается на стороне более жизнестойких индивидов с высоким уровнем шаперонов. Наилучшая стратегия в такой обстановке, по-видимому, состоит в том, чтобы не класть все яйца в одну корзину. Черви минимизируют риск вымирания, позволяя данному признаку широко варьировать даже у генетически идентичных особей (а природные микропопуляции C. elegans часто имеют низкое генетическое разнообразие из-за малого числа основателей и самооплодотворения). Как бы ни сложились обстоятельства, какая-то часть генетически однообразной, но фенотипически вариабельной популяции с большой вероятностью выживет и будет успешно размножаться.
Это исследование интересно тем, что обращает внимание на разновидность изменчивости, которую биологи обычно игнорируют в своих теоретических построениях: на случайную изменчивость, определяемую не генами и не средой, а неизбежными стохастическими флуктуациями на молекулярном уровне, в том числе на уровне экспрессии генов. По-видимому, роль такой изменчивости в жизни и эволюции организмов важнее, чем принято считать.
Молекулярные механизмы реакции на тепловой шок сходны у большинства животных. Вариабельность по силе реакции на стресс характерна для многих организмов. Поэтому можно предположить, что некоторые виды стрессовых воздействий способны сглаживать эффекты вредных мутаций не только у C. elegans, но и других животных. Не исключено, что некоторые из полученных выводов могут быть приложимы и к наследственным болезням человека. Но это пока только предположение.
—————
Сложность организма и сложность программы развития
Соотношение между кажущейся сложностью организма и реальной сложностью генно-регуляторных сетей, составляющих его «программу развития», далеко не однозначно. Видимая сложность порой порождается весьма простой программой.
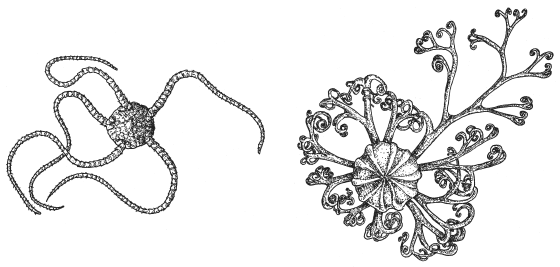
Например, чтобы перейти от внешне простой типичной офиуры (на рисунке слева) к причудливому горгоноцефалюсу с фрактально ветвящимися руками (справа), достаточно, чтобы каждая рука в какой-то момент своего развития раздвоилась, так же как и каждая ветвь руки. Для этого нужно добавить в программу развития следующую инструкцию для растущих концов рук: «При выполнении таких-то условий разветвись надвое». В результате одной-единственной команды автоматически получается сложная, завораживающая фрактальная структура.
Для чего же на самом деле необходимо усложнение генетической программы развития? Герберт Спенсер (1820–1903), философ и социолог, с легкой руки которого дарвиновское «наследование с изменением» (descent with modification) стали называть эволюцией, дал прекрасное определение тому, что мы сейчас назвали бы «прогрессивным развитием» или «развитием по пути усложнения». Спенсеровское определение таково: прогрессивное развитие — это переход от неопределенной несвязной однородности к определенной связной разнородности.
Так вот, усложнение программы развития необходимо для изменений, подпадающих под это определение. Этот вывод трудно обосновать экспериментально — мы пока слишком многого не знаем о програмах развития реальных организмов, — но он быстро становится очевиден пользователю, поигравшему немного с программой EvoDevo.
Усложнение генетической программы развития необходимо, во-первых, для клеточной дифференцировки. Если мы хотим, чтобы из одного исходного типа эмбриональных клеток получились клетки нескольких разных типов (а это и есть переход от однородности к разнородности), нам придется для каждого типа клеток написать свою «подпрограмму».
Во-вторых — это менее очевидно, хотя не менее важно, — усложнение программы необходимо для повышения упорядоченности фенотипа, для подавления «внутреннего хаоса» и роста устойчивости к внешним помехам. Повышение упорядоченности — это, по Спенсеру, переход от неопределенного состояния к определенному. Все эти «избыточные» регуляторные контуры и отрицательные обратные связи, которые приходится добавлять в программу, чтобы развитие стало стабильным, — это самое настоящее, подлинное усложнение, хотя на фенотипическом уровне оно может проявиться всего-навсего в снижении стохастической изменчивости, в более редком появлении всевозможных уродств. При этом каждый дополнительный регуляторный контур, развившийся «для стабильности», может в новых условиях сработать каким-то неожиданным, но интересным образом и стать источником эволюционных новшеств.
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
ВТМ помогает изучать мутации
ВТМ помогает изучать мутации Вирус табачной мозаики, который уже не раз помогал нам разгадать многие загадки жизни, предлагается и в качестве модели для изучения мутаций. ВТМ содержит рибонуклеиновую кислоту, которая, как мы увидим далее, выполняет ту самую генетическую
Дезинфекция спасает матерей и помогает хирургам
Дезинфекция спасает матерей и помогает хирургам Еще до того, как Пастер и Кох доказали, что бактерии являются возбудителями многих инфекционных болезней, главный врач родильного дома в Вене Игнац Земмельвейс объявил войну неизвестным врагам жизни. В этом родильном доме
Ответ на стресс
Ответ на стресс В стабильной среде разница между мужским и женским полом сводится лишь к чисто анатомическим отличиям репродуктивных органов. Иначе говоря, половой диморфизм сведен к минимуму. Однако стоит среде начать меняться, и это опосредованно вызывает расхождение
4.5. Стресс
4.5. Стресс Понятие стресса ввел в 1927 г. американский физиолог У. Кеннон (1871–1945) для обозначения физиологических реакций, возникающих в организме человека и животных под действием несущих угрозу факторов. Наибольший вклад в исследования этого явления внес известный
СТРЕСС
СТРЕСС Наш организм постоянно подвергается неблагоприятным воздействиям, которые могут иметь физический характер. Например, сильное охлаждение или перегрев тела, потеря крови и различные травмы. Неблагоприятными воздействиями на организм могут быть лишения
Как самец лягушки ринодермы помогает самке выполнять родительские обязанности?
Как самец лягушки ринодермы помогает самке выполнять родительские обязанности? Ринодерма (Rhinodermatinae) – небольшая яркоокрашенная лягушка, обитающая в горах Южной Америки. Когда самка откладывает в воду немногочисленные крупные яйца, самец захватывает их ртом и
Как императорский пингвин-самец помогает самке в выведении птенцов?
Как императорский пингвин-самец помогает самке в выведении птенцов? Императорские пингвины, самые крупные из пингвинов, размножаются на льду без каких-либо гнезд. Отложив на лед или снег яйцо, самка передает его самцу, а сама уходит кормиться у кромки льда. Держа на лапах
Брат помогает брату
Брат помогает брату Почему у совы все птенцы разного возраста?Если вы заглянете в гнездо к птице (только не хищной), то найдете там птенцов, которые мало отличаются друг от друга и величиной и, значит, возрастом. Иначе и быть не может: ведь почти все птицы начинают
Неконтролируемый стресс в семье
Неконтролируемый стресс в семье Эффективное применение неконтролируемости как средства манипуляции поведением другого человека описано У. Шекспиром в «Укрощении строптивой» – пьесе, которую принято считать «шедевром комедийного искусства». Петруччо, сам и через
Стресс и чувство тревоги
Стресс и чувство тревоги «Стресс» — это современное слово, которое широко используется — и часто неправильно — во многих популярных журналах и книгах. Тысячи пособий по практической психологии обещают научить, как избежать стресса или справиться с ним. Но, согласно
Почему помогает психотерапия
Почему помогает психотерапия Итак, психические расстройства могут быть нежелательным последствием слишком интенсивного сообщения эпигенома мозга с внешней средой. Вместе с другими факторами эпигенетические переключатели меняют биохимический баланс нервной системы.
Как эпигеном помогает в учебе
Как эпигеном помогает в учебе Научных фантастов уже давно занимает идея целенаправленного стирания человеческих воспоминаний. Эту идею блестяще пародирует фильм «Люди в черном». Супергерой агент Джей в исполнении Уилла Смита мужественно сражается со всевозможными
Длительный стресс старит
Длительный стресс старит Действительно важно в теломерах и теломеразе то, что они чувствительны к сигналам извне. Положительное воздействие окружающей среды и других клеток может как стабилизировать защитные колпачки на концах хромосом, так и увеличить количество