Глава 10 Мир вирусов и его эволюция
Глава 10
Мир вирусов и его эволюция
Пер. Г. Януса
Вирусы были открыты как нечто совсем непримечательное, а именно необычная разновидность инфекционных агентов, а возможно, и особый род токсинов, вызывающих болезни растений, например табачную мозаику. Так как эти агенты проходили сквозь тонкие фильтры, задерживающие бактерии, было сделано верное предположение, что они отличаются от (типичных) бактерий. Вскоре после этого были открыты первые вирусы, поражающие животных. В их числе — вирус саркомы Рауса, первый известный вирус с канцерогенными свойствами, были открыты и удивительные патогены, которые, казалось, пожирали бактерии — их назвали бактериофагами, а в итоге они оказались бактериальными вирусами. В дальнейшем, в течение XX столетия, вирусологию ожидало блистательное развитие (Fields et al., 2001) — по двум причинам. Во-первых, вирусы важны для медицины и сельского хозяйства. Во-вторых, вирусы — простейшие генетические системы и потому стали излюбленными моделями, сначала для ранней молекулярной генетики (прежде всего благодаря работам знаменитой «фаговой группы» под руководством Макса Дельбрюка (Cairns, 1966), а затем для геномики. Однако к 1970-м годам генетика, а к концу 1990-х и геномика достаточно окрепли, чтобы продуктивно работать и с клеточными моделями[99]. В результате вирусология потеряла ведущую роль в фундаментальной биологии (появляясь, впрочем, в эпизодах).
Первая декада нового тысячелетия была отмечена настоящим возрождением вирусологических исследований, вызванным двумя группами открытий. Первым было обнаружение гигантских вирусов, таких как мимивирус. Их вирусные частицы и геномы достигают размеров клеток, размывая казавшуюся очевидной границу между вирусами и клетками (Raoult et al., 2004; Van Etten et al., 2010). Вторая, еще более замечательная серия открытий, была сделана в рамках метагеномики: к великому удивлению биологов, оказалось, что вирусы — наиболее распространенные биологические объекты на Земле (Edwards and Rohwer, 2005). Эти достижения стимулировали намного более широкий интерес к эволюции вирусов. Я рассматриваю эти результаты как открытие обширного древнего мира вирусов, который является неотъемлемой и основополагающей частью живого с момента его зарождения на Земле. Все это время мир вирусов интенсивно взаимодействовал с клеточными формами жизни, выработавшими колоссальное многообразие систем противовирусной защиты, но сохранил свою обособленность и, во многих отношениях, остается ключом ко всей истории жизни. В этой главе мы обсудим вирусный мир, его развитие, а также гонку вооружений между вирусами и клетками, которая пронизывает всю эволюцию[100]. С точки зрения автора, вирусы представляют собой одну из «империй жизни», другая же, разумеется, представлена клеточными организмами. В главе 11 обсуждается вклад вирусов в возникновение и эволюцию клеток.
Необыкновенное разнообразие и повсеместное распространение вирусов
Что такое вирус?
Определения в биологии даются трудно и никогда не являются достаточно полными. Тем не менее, прежде чем рассматривать в этой главе различные аспекты эволюции вирусов, их надо как-то определить. Действительно, нетрудно дать грубое общее определение. За последнее столетие знания о вирусах развились от туманных предположений их первооткрывателей, Дмитрия Ивановского и Мартина Бейеринка, до тончайших молекулярных деталей. Здесь мы определим вирусы следующим, очень общим образом: «облигатные внутриклеточные паразиты или симбионты, обладающие собственными геномами, кодирующими информацию, необходимую для вирусной репродукции (отсюда некоторая степень автономии от генетических систем хозяина), но не кодирующими всю систему трансляции и мембранного аппарата». Это определение подойдет ко всякому «по-настоящему» эгоистичному генетическому элементу. Ключевая фраза тут «кодирующие информацию, необходимую для вирусной репродукции, а следовательно, обладающие некоторой степенью автономии от генетических систем хозяина». Таким образом, обычные гены и опероны под это определение не подойдут, даже если обладают некоторыми эгоистическими чертами, потому что не кодируют никаких специальных приспособлений для самовоспроизведения. Таким образом, это определение охватывает все обширное множество биологических объектов, кодирующих «нечто», необходимое для самовоспроизведения, но не систему трансляции и мембраны.
Я специально не указываю, что это «приспособление для самовоспроизведения» обязано быть белком, так что определенно подойдут и вироиды (растительные патогены с геномами размером всего около трехсот нуклеотидов, использующие для репликации транскрипционный аппарат хозяина). Также я не указываю, что вирусный геном обязательно должен кодировать капсид (то есть белковую оболочку вириона). Это могло бы показаться несколько контринтуитивным, поскольку исторически вирусы были известны в основном как частицы (вирионы), начиная с первой удачной кристаллизации вируса табачной мозаики Уэнделлом Стенли в 1934 году. Капсид настолько приковывает внимание, что Патрик Фортер и Дидье Рауль недавно определили вирусы как «капсид-кодирующие организмы», в противоположность клеточным формам жизни, определенным как «рибосомокодирущие организмы» (Raoult and Forterre, 2008). Это определение указывает правильное направление, если требуется разграничить клетки и вирусы как две основные формы жизни, но оно безосновательно узко и не очерчивает границы мира вирусов объективно.
Конечно, наличие капсида — очень важная и часто встречающаяся черта. Однако в этой главе мы рассмотрим четкие эволюционные взаимосвязи, сопровождаемые сходством геномной архитектуры и циклов репликации, существующие между традиционными, капсид-кодирующими вирусами и «голыми» эгоистичными элементами, такими как плазмиды и разнообразные мобильные элементы. В рамках данного определения (хотя, как и все определения, оно имеет границы применимости) все эти агенты принадлежат обширному миру вирусов.
Время от времени вспыхивают дискуссии на занимательную тему — являются ли вирусы «живыми». Последняя версия этого спора привлекла существенное внимание (Moreira and Lopez-Garcia, 2009). Сам по себе этот вопрос носит исключительно семантический характер и, следовательно, не является значимым. Приведенное здесь определение ясно отводит вирусам место во владениях биологии; как будет изложено далее в этой главе, сравнительная геномика выявила множественные связи между геномами вирусов и клеточных форм жизни. Неудачным последствием отрицания статуса «живых» за вирусами будет то, что тогда вирусы не будут иметь очевидного отношения к эволюции клеточных форм жизни. В этой и следующей главах мы увидим, что это противоречило бы действительности.
Разнообразие стратегий репликации-экспрессии среди вирусов
Все клеточные формы жизни обладают геномами, представленными дцДНК, которая транскрибируется в мРНК (транслирующиеся во многочисленные белки), а также в разнообразные некодирующие РНК. Единообразие генетического цикла клеточных форм жизни находится в разительном контрасте с вариабельностью циклов репликации-экспрессии у вирусов, некоторые из которых обладают РНК-геномами разной полярности, в то время как другие обладают геномами в виде оцДНК (Baltimore, 1971; см. рис. 10-1[101]). Некоторые вирусы и вирусоподобные элементы освоили переход от РНК к ДНК, сочетая действие кодируемой вирусом обратной транскриптазы (ОТ) и ДНК-зависимой РНК-полимеразы хозяина. Вирусы с плюс-цепью РНК отличаются использованием простейшего вообразимого генетического цикла, в то время как обратно транскрибирующиеся элементы связывают мир РНК и мир ДНК. Такая пластичность циклов вирусной репликации может иметь важные эволюционные следствия, о чем будет рассказано в следующей главе.
Когда разнообразие вирусных стратегий репликации-экспрессии стало очевидным, возникло искушение «сыграть в Менделеева» и создать исчерпывающую таблицу возможных циклов репликации-экспрессии, заполнить ее наблюдаемыми разновидностями, а затем попытаться предсказать, какие клетки таблицы заполнятся благодаря будущим открытиям, а какие окажутся «запрещенными» по каким-либо фундаментальным причинам. Насколько мне известно, первую такую попытку предпринял мой учитель вирусологии, Вадим Израилевич Агол (Agol, 1974, первоначальная публикация вышла в мало кому теперь известном русском научном журнале, в котором я ее и прочел). Именно эта статья, благодаря ее захватывающей ясности и ценнейшей попытке (по крайней мере, такой воспринимал ее я в то время, между первым и вторым курсом университета) воспользоваться глубокими, пусть даже и простыми, соображениями симметрии в биологии, прежде всего и побудила меня изучать вирусы. Я никогда не сожалел об этом решении; спустя годы я выработал собственную версию классификации стратегий геномов (Koonin, 1991)[102]. Помимо незыблемой центральной догмы, кажется, есть один фундаментальный запрет: одноцепочечная ДНК никогда не транслируется, поэтому РНК всегда вовлечена в цикл самовоспроизводства любого генетического элемента. В отличие от случая с белками в рамках центральной догмы, здесь, по-видимому, нет прямого химического обоснования для такого запрета (и на самом деле, экспериментально трансляция оцДНК была продемонстрирована Hulen and Legault-Demare, 1975; McCarthy and Holland, 1965). Однако единственная известная нам система трансляции, очевидно, эволюционировала в направлении синтеза белков лишь на матрице РНК (подробнее см. гл. 12). Помимо этого исключения, все возможные на базе молекул РНК и ДНК циклы репликации-экспрессии, по-видимому, реализованы в мире вирусов, хотя некоторые экзотические формы генома, такие как гибрид РНК-ДНК, встречаются редко (см. рис. 10-1).
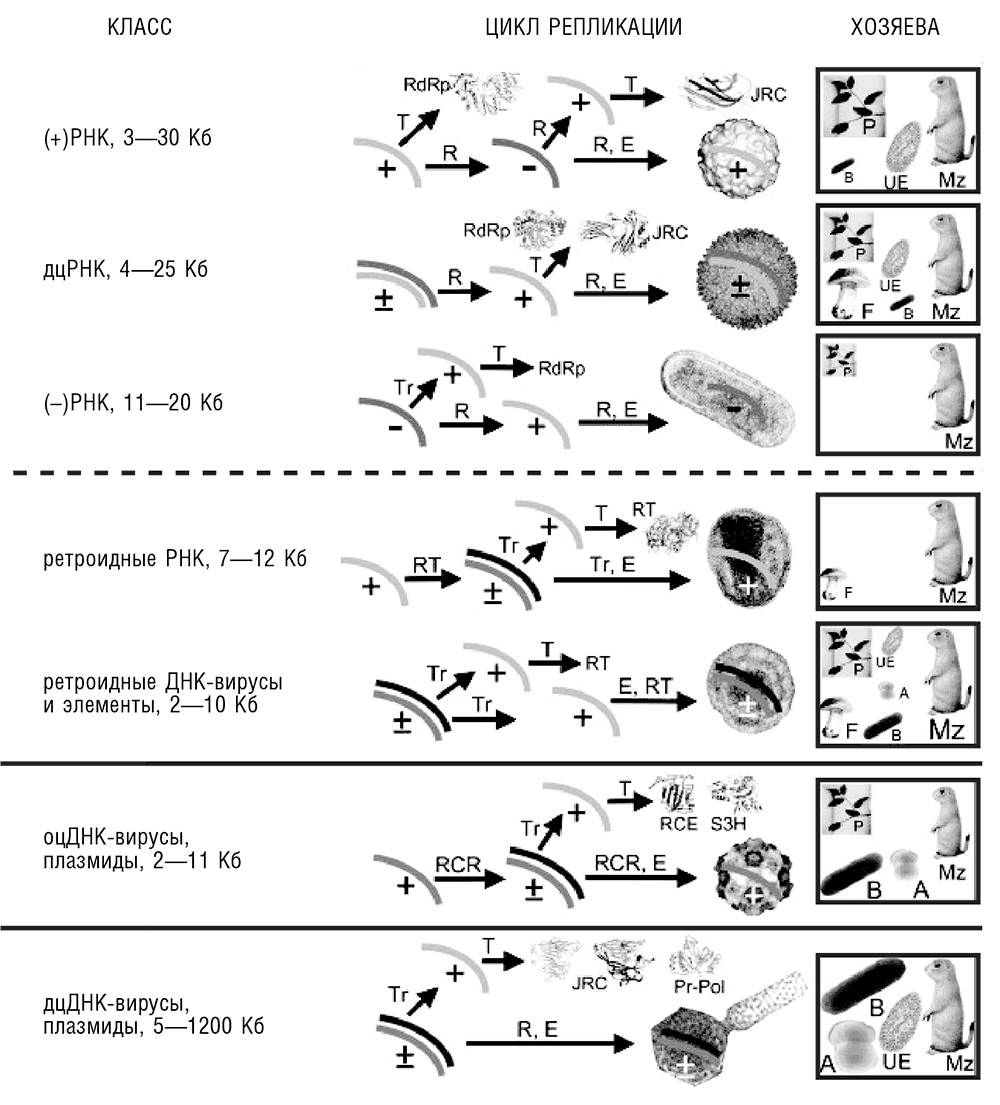
Рис. 10-1. Разнообразие стратегий репликации-экспрессии у вирусов и вирусоподобных элементов. Для каждого класса вирусов и схожих элементов указаны приблизительные размеры генома (Кб, в тысячах оснований). «+» обозначает плюс-цепь (однонаправленную с мРНК), «–» обозначает минус-цепь. Tr — транскрипция, T — трансляция, R — репликация, E — капсидирование вируса, A — археи, B — бактерии, F — грибы, Mz — животные, P — растения, UE — одноклеточные эукариоты. Для каждого класса вирусов (элементов) показаны типичные структуры их белков-маркеров, а также ультрамикроскопические изображения характерных представителей. RdRp — РНК-зависимая РНК-полимераза; JRC — белок капсида с укладкой типа рулета; RT — обратная транскриптаза; RCR — (инициирующая) эндонуклеаза репликации способом катящегося кольца. Справа показан диапазон возможных хозяев. Размер соответствующего изображения и сокращения приблизительно соответствует представленности данного класса вирусов в этом таксоне (по Koonin et al., 2006, с изменениями).
Диапазон сложности геномов, их функционального содержания и разнообразие геномной архитектуры вирусов
Диапазон размера геномов вирусов с различными геномными стратегиями весьма велик: размеры геномов самого крупного известного вируса — мимивируса — и самых мелких вирусов (таких как цирковирусы) отличаются на три порядка. Если мы включим сюда вироиды, которые не кодируют белки, но являются полноценными эгоистичными генетическими элементами и даже патогенами, размах колебаний расширяется до четырех порядков (см. рис. 10-1). Если иметь в виду, что в геномах вирусов обычно белок-кодирующие гены расположены вплотную, то колебания количества генов укладываются приблизительно в тот же диапазон. Размер генома сильно зависит от природы генома и цикла репликации-экспрессии. Видимо, лишь у вирусов, содержащих дцДНК, геном может достичь больших (по вирусным меркам) размеров, более 35 Кб, и (на данный момент) вплоть до 1,1 Мб (Van Etten et al., 2010). Все классы РНК-вирусов, все элементы, способные к обратной транскрипции, и все вирусы, содержащие оцДНК, обладают небольшими геномами, никогда не превышающими 35 Кб, — и то к этому пределу приближается лишь одна группа довольно редких РНК-вирусов животных (коронавирусы и родственные им формы, составляющие порядок Nidovirales (Gorbalenya et al., 2006). Причина этого очевидна: бульшая химическая стабильность и регулярная структура дцДНК благоприятна для функций хранения информации и репликации, а далее эволюция систем репарации дцДНК еще больше усилила функциональное разделение между дцДНК и другими формами нуклеиновых кислот. В дополнение к разнообразию стратегий репликации-экспрессии и широкому диапазону размеров, вирусные геномы принимают все возможные молекулярные конфигурации, включая как линейные, так и циркулярные молекулы ДНК или РНК и как единичные, так и разделенные на множественные сегменты геномы (хромосомы, см. рис. 10-1).
Функциональный набор вирусных генов разительно различается в зависимости от цикла экспрессии и, что еще важнее, от размера генома и генетической сложности. Малые вирусные геномы кодируют практически исключительно белки, непосредственно участвующие в репликации генома, а также субъединицы вириона. Часто — и для всех известных РНК-вирусов и элементов, способных к обратной транскрипции, это именно так, — вирус кодирует полимеразу, участвующую в репликации его собственного генома. Это легко объяснить: клеточные хозяева вируса обычно не кодируют РНК-зависимой РНК-полимеразы или обратной транскриптазы, которая могла бы осуществить репликацию или обратную транскрипцию длинных молекул РНК. РНК-зависимые РНК-полимеразы и обратные транскриптазы, которые кодируются в геномах клеточных форм жизни и осуществляют «обычные» клеточные функции, такие как теломераза и РНК-зависимая РНК-полимераза, участвующая в РНК-интерференции эукариот, способны синтезировать лишь короткие олигонуклеотиды (см. гл. 7 и обсуждение ниже в этой главе). У ДНК-вирусов, напротив, есть возможность использовать аппарат репликации (и транскрипции) хозяина, и они широко пользуются этой возможностью. Так, многие вирусы этого типа, в частности большинство известных вирусов, поражающих архей, так же как и многочисленные умеренные бактериофаги (такие как фаг лямбда, классический модельный организм), не кодируют даже репликативной полимеразы и никаких других белков, непосредственно участвующих в репликации. В таких случаях последовательности ДНК, ответственные за распознавание и привлечение репликативного аппарата хозяина, оказываются основными факторами, определяющими возможность автономной репродукции вируса (см. определение выше), хотя вирусные белки выполняют другие важные функции в вирусной репродукции, такие как подавление или репрограммирование экспрессии генов и метаболизма хозяина. Вирусы с самыми большими геномами, напротив, вдобавок к белкам, составляющим аппарат репликации вирусного генома, кодируют целую коллекцию разнообразных белков, участвующих в процессах репарации, мембранного транспорта, ряде метаболических путей, а в некоторых случаях и трансляции. Обычно (и во всех случаях, когда затрагивается трансляция) вирус кодирует не систему или путь целиком, а только ферменты для одного-двух шагов, которые дополняют или видоизменяют соответствующие функциональные системы клетки хозяина.
Вирусы, содержащие различные формы нуклеиновых кислот, распределены по таксонам хозяев не равномерным и не случайным образом. В частности, необычайное разнообразие содержащих дцДНК бактериофагов и вирусов архей находится в разительном контрасте с отсутствием дцДНК-вирусов у растений. РНК-вирусы, напротив, чрезвычайно распространены и разнообразны у растений и животных, но среди бактерий на данный момент представлены лишь двумя небольшими семействами и до сих пор не обнаружены у архей[103] (см. рис. 10-1). В некоторых случаях биологическая подоплека характерного спектра хозяев вируса совершенно ясна. Например, в растениях крупные вирусы столкнулись бы с серьезными проблемами с межклеточным распространением, так как плазмодесмы (каналы, соединяющие окруженные клеточной стенкой клетки растений) непроницаемы для больших частиц и даже больших молекул ДНК. Однако в большинстве случаев причины предпочтительного распределения вирусов среди тех или иных групп хозяев остаются неизвестными. Например, сложно сказать, почему РНК-вирусы столь распространены среди растений и животных, но не среди прокариот; далее в этой главе мы вернемся к рассмотрению этого вопроса с другой, эволюционной позиции.
Метагеномика вирусов, экспериментальная вирусология, агенты переноса генов и повсеместное распространение вирусов
Вирусы — вездесущие спутники клеточных форм жизни: при более-менее детальном изучении любой клеточный организм оказывается населенным вирусами. У тех организмов — таких как нематоды, — у которых настоящие вирусы пока не были открыты, в геном встроены многочисленные мобильные элементы[104].
С недавних пор изучение виромов (всего множества вирусов, обнаруживаемых в данной среде обитания) стало областью науки, испытывающей бурное развитие (Edwards and Rohwer, 2005; Kristensen et al., 2010). Выделение вирома — методически довольно незамысловатая операция, так как вирусные частицы (по крайней мере подавляющее большинство, исключая, возможно, их гигантские разновидности) проходят сквозь фильтры, непроницаемые даже для мельчайших клеток. Так что достаточно просто собрать частицы в фильтрате и анализировать состав. Изучение виромов принесло большие неожиданности. Первой оказалась сама концентрация вирусных частиц. Поразительно, но по крайней мере в морской среде вирусы (прежде всего бактериофаги) оказались наиболее распространенной из биологических форм, причем в совокупности количество вирусных частиц превосходит количество клеток по меньшей мере на порядок. Это не вполне корректный способ сравнения, так как одна инфицированная вирусом клетка способна продуцировать сотни вирусных частиц, и тем не менее такие данные показывают, что вирусы исключительно широко распространены и активны в окружающей среде. Сейчас вирусы рассматривают в качестве крупных геохимических действующих сил, так как уничтожение вирусами микроорганизмов оказывает сильное влияние на формирование органического осадка (Suttle, 2007, 2005). Вторая крупная неожиданность — это огромное генетическое разнообразие виромов, а также их непредвиденная генетическая композиция. Набор генов в ДНК-вироме резко отличается от набора генов известных бактериофагов. Виром в основном состоит из редких и уникальных генов, для которых не удается найти и подобрать гомологичных последовательностей в имеющихся базах данных. Хотя гены, специфичные для бактериофагов, встречаются в виромах чаще, чем в микробиомах, они представляют ничтожное меньшинство; большинство генов, для которых обнаружены гомологи, представляются случайным набором бактериальных генов. Исключив возможность массивной контаминации, которая, учитывая тщательно разработанные протоколы, используемые для выделения виромов, маловероятна, мы приходим к выводу, что в виромах в основном представлены не обычные вирусы, а некие другие биологические объекты (Khristensen et al., 2010).
Какова природа этой «темной материи», которая доминирует в виромах? На самом деле мы этого не знаем, однако легко предложить правдоподобную гипотезу. Вспомним агенты переноса генов (АПГ), которые уже обсуждались в главах 5 и 9. Агенты переноса генов — особая разновидность псевдовирусов (Lang and Beatty, 2007). Они образуют вирусные (фагоподобные) частицы, которые состоят из белков, кодируемых дефектным профагом, находящимся в соответствующей бактериальной или архейной хромосоме. Однако частицы АПГ не содержат ДНК профага (так что они не являются настоящими бактериофагами), но вместо этого служат оболочками для, судя по всему, случайных фрагментов бактериальной хромосомы. Легко представить, что темная материя виромов в основном состоит из АПГ (Kristensen et al., 2010), так что по сути дела это всего лишь «псевдовиромы». Эта простая гипотеза, которая, конечно, нуждается в эмпирической проверке, имеет далеко идущие следствия. В самом деле, если вирусные (или вирусоподобные) частицы — самые распространенные биологические объекты на Земле и большая часть из них — агенты переноса генов, тогда неизбежен логический вывод, что в биосфере доминируют АПГ. В сочетании с присутствием многочисленных, часто «скрытых» профагов и других мобильных элементов в бактериальных и архейных геномах (Cortez et al., 2009) и еще большим изобилием (чаще неактивных) эгоистичных элементов в геномах многих эукариот (включая человека), эти наблюдения наводят на мысль, что мир вирусов в большой степени «выстраивает» геномы клеточных форм жизни и таким образом определяет эволюцию жизни в целом. Необходимо еще тщательно продумать и изучить в дальнейшем фундаментальные следствия такого вывода, и мы не раз вернемся к ним в этой и следующих главах.
Хотя метагеномика вирусов — еще молодая область науки, она уже замечательным образом преобразила наше понимание мира вирусов, причем это далеко не сводится к удивительным данным по генетическому составу виромов. Одна группа открытий была сделана в результате анализа последовательностей, полученных в ходе глобального исследования океана, грандиозного метагеномного проекта Крейга Вентера (Yooseph et al., 2007). Оказывается, что помимо несметного числа бактериальных последовательностей (основной цели проекта) база данных глобального исследования океана содержит многочисленные последовательности, гомологичные консервативным генам больших нуклеоцитоплазматических ДНК-вирусов (NCLDV, подробнее о них позже в этой главе), заражающих эукариот. Независимо от конкретного источника вирусной ДНК (гигантские вирусы, зараженные пикоэукариоты, проходящие бактериальные фильтры, или, вероятнее всего, и те и другие), разнообразие нескольких семейств NCLDV вышло далеко за границы, очерченные традиционной вирусологией (Monier et al., 2008). Вторая группа замечательных открытий сделана в метагеномике морских РНК-вирусов: были открыты многочисленные РНК-вирусы, заражающие одноклеточных морских эукариот, и весьма неожиданно оказалось, что все они принадлежат к одному суперсемейству вирусов, ранее обнаруженных у животных и растений, — пикорнаподобным вирусам (Koonin et al., 2008). В совокупности эти открытия показывают, что наши представления о мире вирусов пока крайне поверхностны — быть может, истинные размеры и разнообразие этого мира превзойдут самые смелые фантазии[105].
Эволюция вирусов: полифилия, монофилия и гены-сигнатуры
В предыдущих разделах этой главы было дано представление о мире вирусов и показано, что он соизмерим по масштабу с миром клеточных форм жизни — и, возможно, количественно доминирует в биосфере. Более того, вирусный мир оказывает определяющее воздействие на эволюцию клеток с помощью агентов переноса генов и огромного арсенала мобильных элементов. Так что изучать эволюцию вирусов необходимо, если мы стремимся к хоть сколько-нибудь глубокому пониманию эволюции жизни в целом.
Сравнительная геномика не свидетельствует в пользу монофилетического происхождения всех вирусов. Здесь мы понимаем под «монофилией» «происхождение от общего предкового вируса или вирусоподобного эгоистического элемента» (Koonin et al., 2006). У многих групп вирусов просто нет общих генов, что позволяет решительно отмести любые идеи об общем происхождении. Идея «общих генов», будучи приложена к вирусам, оказывается непростой: в мире вирусов общность не обязательно ограничивается однозначными отношениями ортологии между генами, которые легко выявить благодаря очевидному сходству последовательностей. Вместо этого, как будет изложено в следующих разделах, отдаленные отношения гомологии между вирусными белками, а также между вирусными белками и их гомологами из клеточных форм жизни могут дать более сложные, но важные свидетельства об эволюции вирусов. Несмотря на эту сложность, среди больших групп вирусов изобилуют случаи, когда между ними либо нет никаких гомологичных генов, какое бы определение мы ни взяли, либо имеются лишь отдаленно родственные домены, чьи эволюционные траектории очевидно различны. Например, большинство вирусов гипертермофильных кренархеот не имеют общих генов ни с какими другими вирусами (Prangishvili et al., 2006b), а РНК-вирусы разделяют с ДНК-вирусами и плазмидами, реплицирующимися способом «катящегося кольца», лишь чрезвычайно отдаленно родственные домены в соответствующих белках репликации.
С другой стороны, можно уверенно показать монофилию нескольких крупных групп вирусов, включая обширные совокупности РНК-вирусов и сложных ДНК-вирусов (см. табл. 10-1). Некоторые из этих монофилетических классов переходят даже границы геномных стратегий: так, монофилетический класс элементов, размножающихся через стадию обратной транскрипции, включает в себя как РНК-вирусы, так и вирусы, мобильные элементы и плазмиды с ДНК-геномами. В классе элементов, реплицирующихся по механизму катящегося кольца, сочетаются вирусы, содержащие одноцепочечную или двухцепочечную ДНК, а также плазмиды. Более того, на основании сходства в структурах комплексов, участвующих в репликации РНК, а также присутствия гомологии, пусть и отдаленной, в ферментах репликации была предложена правдоподобная гипотеза об общем происхождении вирусов, содержащих позитивную РНК-нить, вирусов, содержащих дцРНК, и элементов, способных к обратной транскрипции (Ahlquist, 2006). Однако в целом вывода, что вирусы имеют много различных линий происхождения, по-видимо му, невозможно избежать (Koonin et al., 2006; см. табл. 10-1).
Краткая естественная история вирусных генов
Анализ последовательностей выявил несколько категорий вирусных генов, принципиально отличающихся по происхождению (Koonin et al., 2006). Можно обсуждать, какая степень дробности классификации оптимальна, но четко различаются по меньшей мере пять классов, укладывающихся в три более крупные категории.
Гены с четко опознаваемыми гомологами у клеточных форм жизни:
1. Гены с близкими гомологами у клеточных организмов (обычно это хозяева данного вируса) присутствуют у узкой группы вирусов.
2. Гены, консервативные среди большой группы вирусов или даже нескольких групп и имеющие относительно отдаленные клеточные гомологи.
Вирус-специфичные гены:
3. Гены-сироты[106] — гены без выявленных гомологов, кроме как у близкородственных вирусов.
4. Вирус-специфические гены, консервативные для (относительно) широкой группы вирусов, но не имеющие гомологов у клеточных форм жизни.
5. Гены — вирусные сигнатуры.
Вирусные сигнатуры — это гены, общие для многих разнообразных групп вирусов, имеющие лишь отдаленные гомологи у клеточных организмов, для которых имеются убедительные свидетельства в пользу монофилии (общего происхождения) всех вирусных членов соответствующих семейств генов. Словосочетание «гены-сигнатуры» было использовано, чтобы подчеркнуть, что эти гены являются признаками «вирусного состояния».
Относительный вклад каждого из этих классов генов в наборы генов различных вирусов зависит от размера вирусного генома и его генетической сложности, которые разнятся более чем на три порядка (см. рис. 10-1). Вирусы с малыми геномами, такие как большинство РНК-вирусов, часто обладают лишь несколькими генами, большинство из которых принадлежат к классу генов-сигнатур. Напротив, у вирусов с большими геномами, например поксвирусов, широко представлены все пять классов. Чтобы проиллюстрировать разнообразие генетического состава, на рис. 10-2 показана разбивка набора генов трех вирусов, обладающих малым, средним и крупным геномами соответственно, на пять классов генов. Интересно, что крупные и среднего размера геномы бактериофагов и вирусов архей заполнены «сиротами», которые зачастую составляют более 80 процентов генов этих вирусов. Быстро эволюционирующие «гены-сироты» фагов, возможно, поставляют много, если не большинство «генов-сирот», которые находят в геномах прокариот (несмотря на отсутствие выявляемой консервативности последовательностей), так что они играют ключевую роль в эволюции прокариот (Daubin and Ochman, 2004).
Происхождение пяти классов вирусных генов, скорее всего, различно. Два класса генов, имеющих легко распознаваемые гомологи в клеточных формах жизни, вероятно, представляют собой соответственно относительно недавние (класс 1) и древние (класс 2) заимствования из геномов клеточных хозяев. Откуда произошли вирус-специфические гены — это значительно более трудный и интересный вопрос. Первая возможность заключается в том, что эти гены произошли от других генов вирусов или хозяев, но резкое ускорение эволюции, связанное с появлением новых, специфических вирусных функций, стерло все следы их происхождения. Это соображение согласуется с тем фактом, что многие (возможно, большинство) генов 4-го класса (вирус-специфических генов, консервативных среди группы вирусов) — компоненты вириона, то есть несут важнейшую для вируса функцию, для которой нет эквивалента у клеточных форм жизни. Мы пока отложим обсуждение других путей возникновения и эволюции вирус-специфических генов и обсудим эволюцию вирусных генов-сигнатур. Гены-сигнатуры, пересекающие границы, пролегающие между самыми разными эволюционными линиями вирусов, — глубоко интересный и значимый объект для понимания эволюции и происхождения вирусов.
Таблица 10-1
Наиболее крупные монофилетические классы вирусов и эгоистичных генетических элементов
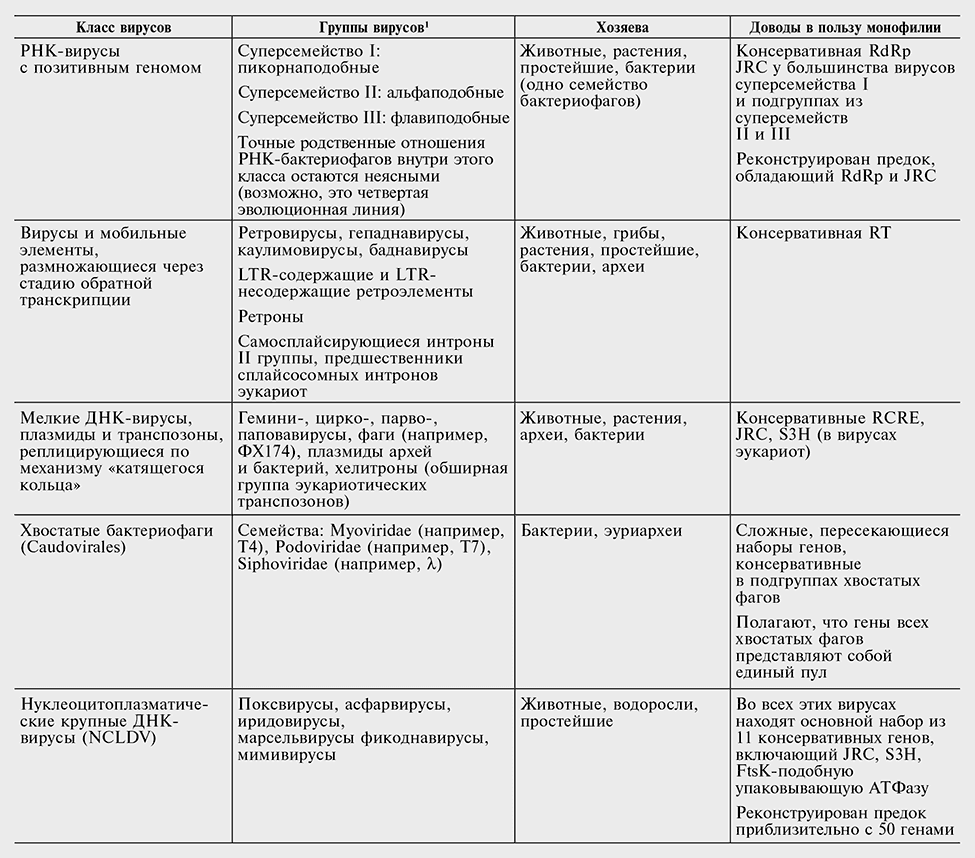
Сокращения: JRC — белок капсида с укладкой типа рулета; LTR — long terminal repeat (длинный концевой повтор); RdRp — РНК-зависимая РНК-полимераза; RCRE — (инициирующая) эндонуклеаза репликации по механизму «катящегося кольца»; RT — обратная транскриптаза; S3H — геликаза суперсемейства 3.
—————
1 Уже после выхода этой книги автору посчастливилось участвовать в работе, где из горячего источника в Йеллоустонском парке, населенного исключительно гипертермофильными кренархеотами, методами метагеномики был выделен и секвенирован РНК-геном нового вируса, очень отдаленно родственного разным семействам позитивных РНК-вирусов эукариот (Bolduc B, Shaughnessy DP, Wolf YI, Koonin EV, Roberto FF, Young M. Identification of novel positive-strand RNA viruses by metagenomic analysis of archaea-dominated Yellowstone hot springs. J Virol. 2012 May;86(10):5562–73). Скорее всего, этот вирус представляет новое суперсемейство позитивных РНК-вирусов, которые, возможно, размножаются в археях. Прямые данные, подтверждающие (или отвергающие) это предположение, еще предстоит получить. Если гипотеза подтвердится, наши представления об эволюции этого класса вирусов могут потребовать существенного пересмотра (см. ниже).
Гены — вирусные сигнатуры: сигналы из древнего мира вирусов
Помимо больших классов, перечисленных в табл. 10-1, отчетливых вертикальных родственных отношений между крупными группами вирусов не существует. Однако значительное число генов, кодирующих белки, которые играют ключевую роль в репликации генома, экспрессии и морфогенезе вирионов, входят в пересекающиеся наборы генов, принадлежащие группам вирусов, казалось бы не родственных ни в каком другом отношении, хотя ни один из этих генов не присутствует у всех вирусов (см. табл. 10-2). Большинство генов — вирусных сигнатур не имеют высококонсервативных гомологов у клеточных форм жизни (исключения — легко распознаваемые провирусы и мобильные генетические элементы), хотя отдаленные гомологи бывают. Два наиболее широко распространенных среди вирусов гена кодируют белок капсида с так называемой укладкой типа рулета и геликазу суперсемейства 3. Оба этих белка пересекают барьер между ДНК— и РНК-вирусами и появляются в поразительно широком ассортименте групп вирусов, от некоторых самых мелких РНК-вирусов с позитивным геномом до крупных нуклеоцитоплазматических ДНК-вирусов, класса вирусов, включающего гигантский мимивирус (см. табл. 10-2). Уточняя, можно сказать, что белок капсида с укладкой типа рулета — главный строительный блок икосаэдрических (сферических) вирусных капсидов, наиболее частой формы капсидов, которые крайне различны по величине, но довольно сходны по симметрии и общей форме среди всего огромного диапазона вирусов, которые используют разнообразные стратегии репликации-экспрессии и заражают хозяев, представляющих все или почти все разнообразие клеточной жизни. Так, геликаза суперсемейства 3 участвует в репликации геномов огромного разнообразия РНК— и ДНК-вирусов.
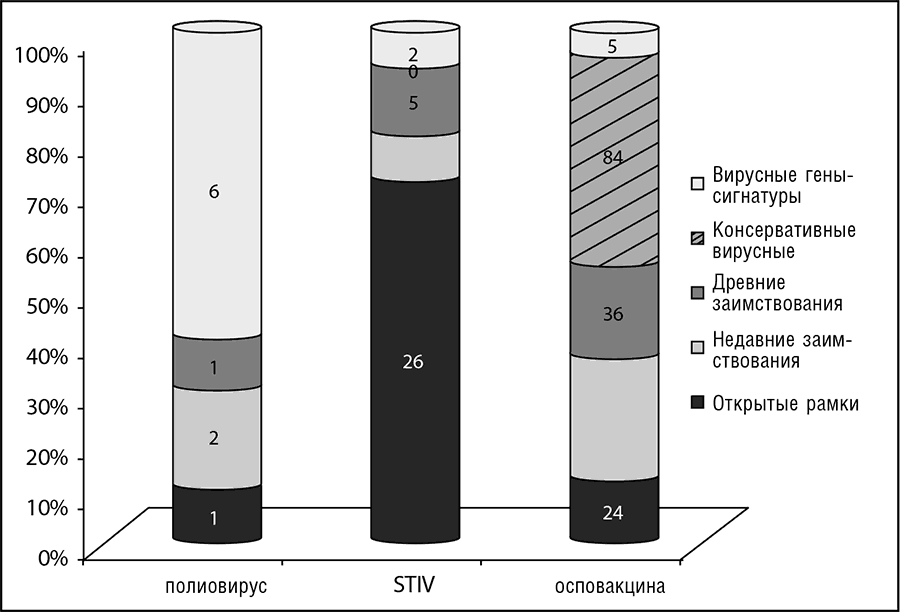
Рис. 10-2. Разбиение вирусных генов на пять эволюционных классов: вирус, обладающий малым геномом: полиовирус (7,4 Кб); вирус, обладающий геномом промежуточного размера: Sulfolobus turreted icosahedral virus (STIV); вирус, обладающий крупным геномом: вирус осповакцины (195 Кб). Данные по Koonin et.al., 2006.
Другие белки, перечисленные в табл. 10-2, не столь широко распространены, как белок капсида с укладкой типа рулета и геликаза суперсемейства 3, но все же за их счет формируются множественные непредвиденные связи между группами вирусов, во всех других отношениях казавшихся неродственными. В качестве примера можно рассмотреть эндонуклеазу инициации репликации по механизму «катящегося кольца», которая объединяет огромное разнообразие небольших репликонов одноцепочечных и двухцепочечных ДНК, включая вирусы, плазмиды и транспозоны, которые размножаются в животных, растениях, бактериях и археях. Детальный анализ аминокислотных последовательностей показал, что ДНК-связывающий домен репликативного белка вирусов полиомы и папилломы (например, T-антигена SV40) представляет собой неактивную производную форму эндонуклеазы инициации репликации способом «катящегося кольца» (Iyer et al., 2005). Таким образом, посредством детального исследования одного из белков-сигнатур хорошо известная связь между различными небольшими репликонами оцДНК (вирусов и плазмид) распространилась также на группу репликонов дцДНК такого же размера. Подобное расширение набора вирусных групп, охватываемых определенным геном-сигнатурой, произошло также в результате детального анализа АТФазы, отвечающей за упаковку вирусной ДНК в капсид, и праймазы архей и эукариот, участвующей в инициации репликации ДНК (Iyer et al., 2005; Iyer et al., 2004b; см. табл. 10-2).
Репликация генома РНК-вирусов с позитивным геномом, дцРНК-вирусов, вирусов с негативным геномом и обратно транскрибирующихся вирусов (элементов) катализируется другим классом ферментов — вирусных сигнатур: РНК-зависимыми РНК-полимеразами и обратной транскриптазой. Полимеразы РНК-вирусов с позитивным геномом и обратная транскриптаза образуют монофилетическую группу внутри обширного класса так называемых palm-доменов («домены-ладони»), характерных для различных полимераз (Iyer et al., 2005; Koonin et al., 2008). РНК-зависимые РНК-полимеразы дцРНК-вирусов и вирусов с негативным геномом, скорее всего, сильно измененные производные того же полимеразного домена (Delarue et al., 1990; Gorbalenya et al., 2002; Koonin et al., 1989). Этот ген-сигнатура может привести нас к самым ранним этапам эволюции жизни, к миру РНК (см. гл. 11 и 12 — там гораздо подробней) и началу мира вирусов. Palm-домен, вероятно, является изначальным белком-полимеразой, сменившим рибозимные полимеразы (гипотетического) мира РНК. Это предположение поддерживается не только широким распространением palm-домена среди современных форм жизни, но еще и структурной, а следовательно, и эволюционной связью между palm-доменом и доменом, содержащим РНК-распознающий мотив (RRM, RNA Recognition Motif), древний РНК-связывающий домен, который первоначально, возможно, способствовал репликации рибозимов (Aravind et al., 2002). РНК-зависимые РНК-полимеразы и обратные транскриптазы исключены из основного хода репликативного цикла клеточных форм жизни, хотя большинство эукариотических геномов, особенно растений и животных, включает в себя множество копий RT-кодирующих ретроэлементов; у прокариот тоже есть немного подобных элементов (см. также гл. 5 и 7). Эти элементы, однако, эгоистичны и с эволюционной точки зрения принадлежат миру вирусов. Возможно, наиболее впечатляющее вторжение обратной транскриптазы в мир клеток — это каталитическая субъединица эукариотической теломеразы, важнейшего фермента, участвующего в репликации концов хромосом[107]. Конечно, не стоит забывать, что все интроны эукариот произошли от ретроэлементов прокариот (см. гл. 7). Примечательно, что единственная другая известная РНК-зависимая РНК-полимераза, не родственная полимеразам, содержащим palm-домен, и являющаяся компонентом системы РНК-интерференции эукариот (см. гл. 7), также, по-видимому, имеет вирусное происхождение (Iyer et al., 2003).
Таблица 10-2
Белки, кодируемые наиболее распространенными генами — вирусными сигнатурами
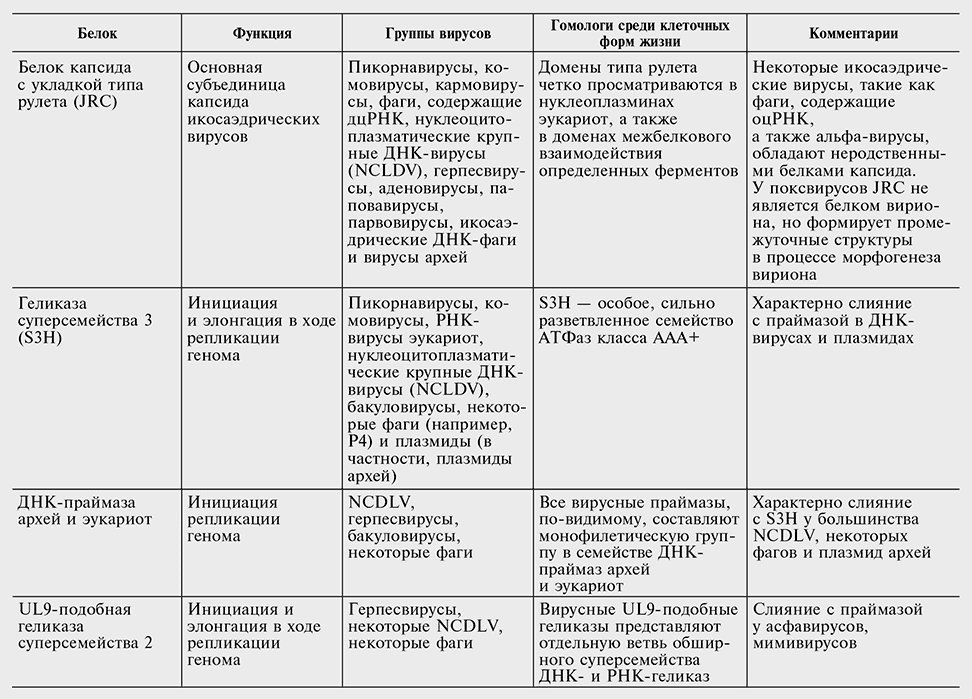
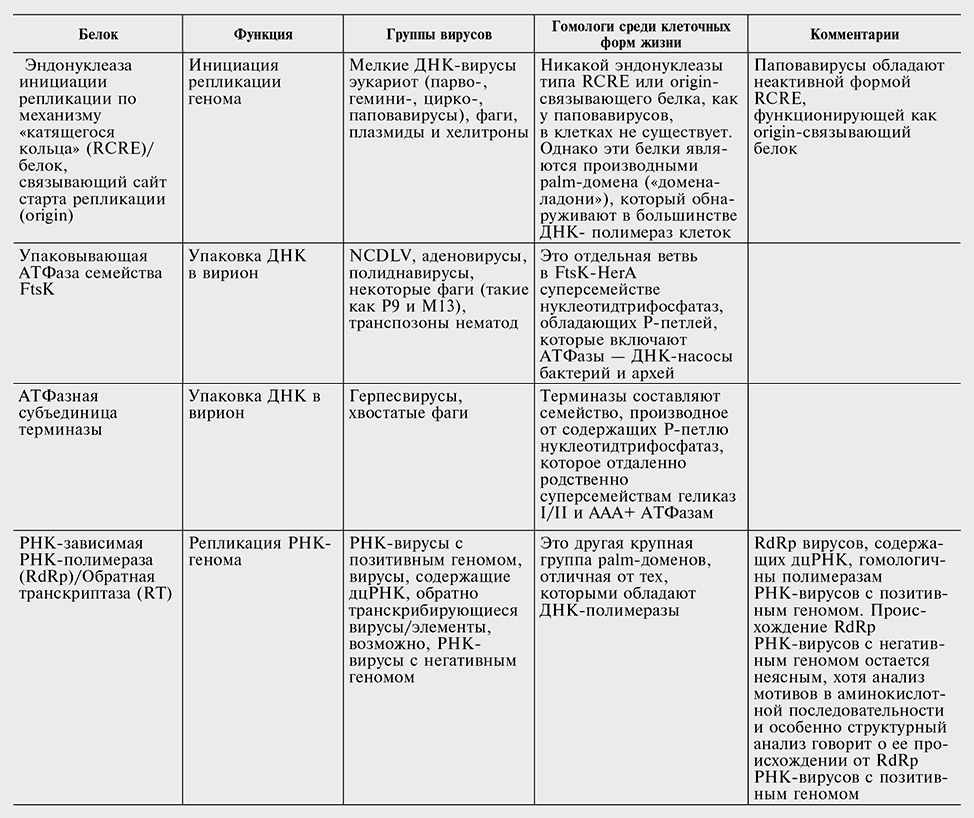
Перечень генов — вирусных сигнатур в табл. 10-2 консервативен. Вероятнее всего, другие гены также заслуживают статуса сигнатур, но отыскать явные свидетельства в пользу этого непросто. Секвенирование новых вирусных геномов в совокупности с всесторонним сравнительным анализом могло бы помочь выявить новые гены, которые, несмотря на относительно узкое распространение среди вирусов, могут считаться «сигнатурами». В самом деле, так может обстоять дело со многими, если не с большинством, генов класса 4, вирусных генов, консервативных для больших групп вирусов, но не клеточных форм жизни.
Комбинация свойств белков — вирусных сигнатур во многом необычна и требует эволюционистского объяснения. В самом деле, все гены-сигнатуры без исключения ответственны за важнейшие, центральные аспекты вирусного жизненного цикла, включая репликацию генома, формирование вириона и упаковку геномной ДНК в вирион (см. табл. 10-2). Обладание этими генами связывает между собой совершенно различные классы вирусов, которые часто имеют совершенно разные стратегии самовоспроизведения и различаются по размеру генома на три порядка. Наконец, у всех генов — вирусных сигнатур есть отдаленные гомологи в клеточных формах жизни (см. табл. 10-2), но вирусные версии, по-видимому, имеют общее происхождение.
Две сразу возникающие гипотезы о происхождении вирусных генов-сигнатур предлагают противоречащие друг другу эволюционные сценарии, ответственные за существование и распространение этих генов (Koonin et al., 2006).
1. Гены-сигнатуры — наследие последнего универсального общего предка вирусов (Last Universal Common Ancestor of Viruses, LUCAV). Этот сценарий предполагает, что, невзирая на все свидетельства в пользу противоположного (см. выше), все ныне живущие вирусы на самом деле монофилетичны, хотя их дальнейшая эволюция включала в себя масштабную потерю генов в некоторых эволюционных линиях, а также обильное заимствование новых генов от хозяев в других.
2. Напротив, в рамках гипотезы о полифилетическом происхождении вирусов распространение генов-сигнатур по всему диапазону групп вирусов может объяснить горизонтальный перенос генов.
При более детальном рассмотрении ни одна из этих гипотез не кажется правдоподобным всеобъемлющим объяснением существования и распределения вирусных генов-сигнатур. Действительно, относительно малое число и мозаичное распространение генов-сигнатур (см. табл. 10-2) вряд ли свидетельствует о существовании LUCAV, хотя очевидно, что множество разнообразных вирусов, если не все они, разделяют некую общую историю. С другой стороны, исключительно отдаленное (но все же различимое) сходство между белками-сигнатурами из различных групп вирусов, обладающих абсолютно разными стратегиями репликации, мало совместимо со сценарием ГПГ.
Ниже в этой главе мы рассмотрим сценарий происхождения и эволюции вирусов, который не включает (в традиционном понимании) LUCAV, но сочетает в себе аспекты общего происхождения и гипотезу ГПГ и естественным образом связан с определенными моделями эволюции клеток. Наиболее простое объяснение того факта, что белки-сигнатуры, участвующие в вирусной репликации и формировании вириона, присутствуют в широком спектре вирусов, но, очевидно, отсутствуют в какой-либо из клеточных форм жизни, заключается в том, что у последних таких генов просто-напросто никогда и не было. Вместо этого наиболее правдоподобный сценарий постулирует, что гены-сигнатуры предшествуют клеткам и происходят непосредственно из первичного, доклеточного пула генов. Как можно представить, в таком первичном пуле отбор будет действовать прежде всего в отношении функций, непосредственно связанных с репликацией, что согласуется со свойствами большинства генов-сигнатур (см. табл. 10-2). Если учесть распространение генов-сигнатур среди многочисленных групп резко различных вирусов, важнейший вывод будет заключаться в том, что крупные классы вирусов и сами возникли на доклеточных этапах эволюции. Этот вывод — ключевой момент концепции древнего мира вирусов. Основная черта мира вирусов состоит в непрерывном потоке генетической информации через огромное разнообразие эгоистичных элементов от доклеточного этапа эволюции до наших дней.
Конкурирующие концепции происхождения и эволюции вирусов
Прежде чем мы обсудим возникающую концепцию происхождения вирусов из первичного пула генов во всей ее полноте, нам надо вкратце рассмотреть существующие гипотезы происхождения и эволюции вирусов. Традиционно эти идеи вращаются вокруг трех тем (см. рис. 10-3):
1. Происхождение вирусов из первичных генетических элементов.
2. Дегенерация одноклеточных паразитов до вирусного состояния.
3. Сценарий «сбежавших генов», который возводит вирусы к генам клеточных организмов, которые сбежали из клеточного генома и переключились на эгоистичный режим самовоспроизводства.
«Первичная» гипотеза была довольно модной в самые первые дни вирусологии, и примечательно, что Феликс д’Эррель (D’Herelle, 1922), первооткрыватель бактериофагов и один из основателей вирусологии, еще в 1922 году предположил, что фаги могут быть эволюционными предшественниками клеток. Несколько лет спустя, в 1928 году, Дж. Б. С. Холдейн предложил ту же гипотезу в классическом эссе на тему происхождения жизни (Haldane, 1928, мы вернемся к поистине пророческим идеям Холдейна в гл. 11). Однако, как только стало ясно, что все вирусы — облигатные внутриклеточные паразиты, «первичная» гипотеза была, по сути, отвергнута в силу простого и, при поверхностном рассмотрении, неопровержимого аргумента, что внутриклеточные паразиты не могут предшествовать появлению полноценных клеток. Напротив, присутствие во многих вирусах (особенно с большими геномами) многочисленных генов, произошедших от хозяев (в противоположность вирус-специфическим генам), может быть истолковано в поддержку гипотезы «сбежавших генов» или даже «дегенерации клетки». Во дни расцвета молекулярной биологии, когда фундаментальные различия между вирусами и клетками были четко осознаны — так что происхождение вирусов от клеток (пусть и дегенерировавших) было сочтено маловероятным, — гипотеза сбежавших генов стала, в большей или меньшей степени благодаря исключению прочих, общепринятой концепцией происхождения вирусов (Luria and Darnell, 1967). Однако недавно открытие гигантских вирусов и особенно тот факт, что эти вирусы обладают некоторыми важнейшими «клеточными» генами, например генами для множества компонентов системы трансляции, привели к воскрешению гипотезы клеточной дегенерации (Claverie, 2006). В самом деле, в терминах размера генома и генетической сложности открытие гигантских вирусов уничтожает границу между вирусами и клеточными формами жизни.
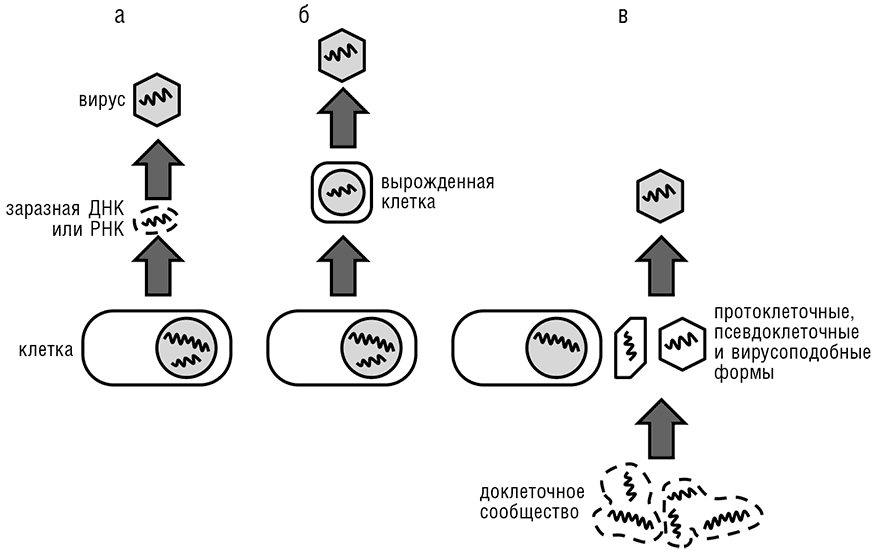
Рис. 10-3. Три конкурирующие гипотезы происхождения вирусов: а — сценарий сбежавших генов; б — сценарий клеточной дегенерации; в — сценарий первичного пула генов.
Невзирая на все эти аргументы, существование вирусных генов-сигнатур, по-видимому, успешно фальсифицирует как гипотезу дегенерации клетки, так и гипотезу сбежавших генов (или, по крайней мере, вызывает серьезные сомнения в них). Что касается гипотезы дегенерации клетки, давайте рассмотрим NCDLV (Koonin and Yutin, 2010), класс крупных вирусов, к которым эта концепция легче всего приложима и на самом деле уже прилагалась вскоре после открытия гигантского мимивируса (см. табл. 10-1). Среди девяти генов, которые объединяют (практически) все NCDLV, три наиболее важных (белок капсида с укладкой типа рулета, геликаза суперсемейства 3 и упаковывающая АТФаза) — вирусные гены-сигнатуры. Даже простейшая предковая форма NCDLV не могла бы без них функционировать. Соответственно, для обоснования клеточного происхождения этого предкового NCDLV потребовалось бы привлечь ad hoc определенно неэкономные сценарии, например согласованную потерю всех генов-сигнатур у всех известных клеточных форм жизни или их происхождение от вымершей крупной эволюционной линии клеток. Та же логика в основном отвергает концепцию сбежавших генов, ввиду того что у генов-сигнатур никогда не было клеточного «дома», из которого они бы сбежали. Иными словами, чтобы спасти гипотезу «сбежавших генов», нужно постулировать существование вымерших клеточных доменов, откуда могли бы сбежать гены-сигнатуры.
Таким образом, наиболее экономный сценарий эволюции вирусов, по-видимому, включает доклеточный мир вирусов. Наиболее вероятным кажется, что основные классы вирусов — по крайней мере все стратегии репликации и экспрессии генома — возникли уже в доклеточную эру. Возможно, называть гипотетические первичные эгоистичесные элементы вирусами нецелесообразно, учитывая отсутствие клеток на этом этапе их эволюции. Однако, если их называть «вирусоподобными» агентами или как-то вроде этого, это никак не изменит того факта, что нет ни следа клеточного происхождении вирусов, и ни в коей мере не опровергнет гипотезу древнего мира вирусов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава VII Половая эволюция
Глава VII Половая эволюция Эволюция всякого живого существа происходит двояко: 1) его онтогения, или вся последовательность развития индивидуума, как такового, считая от зачатия вплоть до его естественной смерти, обусловленной старостью, и 2) его филогения, или развитие его
Глава 6 Эволюция
Глава 6 Эволюция Естественный отборУченым, который открыл научному миру эволюционный механизм, был английский натуралист Чарлз Роберт Дарвин (1809—1882), внук Эразма Дарвина, упомянутого выше.В молодости Дарвин пытался изучать медицину, а позже подумывал о посвящении в
Глава 7. Созидающая эволюция
Глава 7. Созидающая эволюция Многие люди полагают, что естественный отбор — это лишь отрицающая сила, способная к искоренению причуд и неудач, но неспособная к созиданию сложности, красоты и эффективности проекта. Действительно ли она просто отнимает от того, что уже
Глава 2. Эволюция
Глава 2. Эволюция Путь от амебы к человеку казался философам очевидным прогрессом. Хотя неизвестно, согласилась бы с этим мнением амеба. Б. Рассел (1872–1970), английский философ, лауреат Нобелевской премии 1950 г. Теория эволюции – это не только общебиологическая теория, но и
Глава 4 Эволюция на наших глазах
Глава 4 Эволюция на наших глазах Часто для того, чтобы принять факт, нам нужно его увидеть. Факт эволюции некоторым людям принять непросто именно потому, что увидеть эволюцию воочию очень трудно. Ведь по меркам человеческой жизни эволюция — чудовищно медленный процесс.
Глава 5. Биологическая эволюция
Глава 5. Биологическая эволюция Теория эволюции, объясняющая механизмы исторического развития живой природы, не просто общебиологическая теория. В значительной степени это мировоззренческая система, уровень разработанности которой имеет решающее значение для
Глава 3 Морфологическая и молекулярная эволюция
Глава 3 Морфологическая и молекулярная эволюция Я думаю, что отец наш небесный выдумал человека, потому что разочаровался в обезьяне. Марк
Глава 8. Эволюция биологических форм
Глава 8. Эволюция биологических форм 8.1. Неодарвинистская теория эволюции Фактически очень мало известно, если вообще когда-нибудь может быть известно, о деталях эволюции в прошлом. Также эволюцию нелегко наблюдать в настоящем. Даже на шкале времени в миллионы лет начало
Глава 11. Наследование и эволюция поведения
Глава 11. Наследование и эволюция поведения 11.1. Наследование поведения Согласно гипотезе формативной причинности, наследование поведения зависит от генетической наследственности, а также от морфогенетических полей, которые контролируют развитие нервной системы и
Глава IV ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Глава IV ЭВОЛЮЦИЯ ЧЕЛОВЕКА Так не скорби о том, что ты умер, Пелид благородный! Гомер. Одиссея. Мысль может порождать Субстанцию, и населять свои миры Созданьями, что ярче, чем живые, и давать Дыханье формам, которые всю плоть переживут. Байрон. Сон. Я начал писать эту книгу
Глава 3 ЭВОЛЮЦИЯ ПСИХИКИ И АНТРОПОГЕНЕЗ
Глава 3 ЭВОЛЮЦИЯ ПСИХИКИ И АНТРОПОГЕНЕЗ С древнейших времен человек пытался как-то осознать свое отношение к миру животных, искал признаки сходства и различия в поведении последних. Об этом свидетельствует та исключительная роль, которую играет поведение животных в
Глава VII. Эволюция людей
Глава VII. Эволюция людей 7.1. Эволюция приматов; возможные пути эволюции гоминин после их отделения от шимпанзе 7.1.1. Ранние предшественники современных людей Плацентарные млекопитающие появились по разным данным 150–100 млн лет тому назад (Kumar and Hedges, 1998; Archibald et al., 2001; Douzery et
Глава 7 Созидательная эволюция
Глава 7 Созидательная эволюция Иногда люди думают, что естественный отбор — сила исключительно отрицательная, способная только устранять уродцев и неудачников, но никак не создавать сложность, красоту и эффективность конструкции. Разве он не удаляет просто-напросто
Глава 15 Эволюция интеллекта
Глава 15 Эволюция интеллекта 1. Общие принципы эволюции интеллекта 2. Млекопитающие 3. Птицы 4. Приматы 5. Гоминиды 6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens Вернемся теперь к