3.5. Мейоз
3.5. Мейоз
Современные представления о цитологических основах наследственности сформировались только после выяснения генетического смысла процесса мейотического деления клеток.
Мейоз – это процесс образования гаплоидных клеток, т. е. клеток, имеющих половинный набор хромосом. Его можно рассматривать как второй тип деления клеток. Мейоз также можно рассматривать и как специфичный вариант клеточной дифференцировки. Таким способом образуются половые клетки (гаметы) и споры.
Гамета – это клетка, способная сливаться с другой гаметой с образованием диплоидной клетки (зиготы), дающей новый организм.
Спора – это клетка, способная самостоятельно развиваться в новый организм.
В результате процесса мейоза из одной диплоидной клетки образуется 4 гаплоидных (гаметы или споры). У большинства организмов мейоз протекает принципиально сходно. Он состоит из двух последовательных делений: редукционное деление (мейоз-1) и эквационное деление (мейоз-2). В каждом из них различают 4 фазы: профазу, метафазу, анафазу и телофазу. Таким образом, весь процесс мейоза условно можно разбить на 8 этапов, плавно переходящих один в другой. Если другие пути на специализацию начинаются после М-периода клеточного цикла, то мейоз начинается после S-периода, т. е. после репликации хромосом.
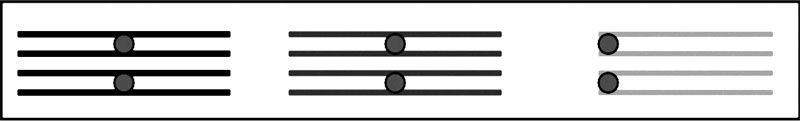
Рис. 3.4. Синапсис гомологичных хромосом с образованием бивалентов в профазе мейоза
Профаза-1. Наиболее сложная, длительная и важная стадия мейоза. Помимо процессов, аналогичных процессам профазы митоза (спирализация хромосом, разрушение ядерной мембраны, исчезновение ядрышка, образование веретена деления), определяющее значение для всего последующего процесса имеет конъюгация гомологичных хромосом – синапсис. Соединенные пары гомологов называются бивалентами (рис. 3.4).
Гомологичные хромосомы связывает особая структура, образованная из белков кариоплазмы – синаптонемный комплекс (СК). В бивалентах гомологичные хромосомы могут обмениваться гомологичными участками. Такой процесс называется кроссинговером. Механизм кроссинговера довольно сложен. Кроссинговер вносит большой вклад в повышение генетического разнообразия, играет важную эволюционную роль и активно изучается на протяжении всей истории генетики. Однако до сих пор он сохраняет свои загадки.
В связи с длительностью и многообразием процессов профазы-1 ее обычно подразделяют на 5 подстадий.
Лептотена – начало спирализации и уплотнения хромосом.
Зиготена – начало (с отдельных участков) и завершение синапсиса гомологичных хромосом. Происходит формирование СК.
Пахитена – укорочение и утолщение бивалентов (стадия толстых нитей).
Диплотена – гомологичные хромосомы бивалентов начинают расходиться (разрушается СК), но они связаны в нескольких зонах контакта – хиазмах. Число хиазм в биваленте может быть различным (обычно 2–3), в длинных хромосомах больше, чем в коротких. Хиазмычасто показывают, что между хроматидами происходит кроссинговер.
Диакинез – хромосомы достигают максимальной спирализации. Исчезают хиазмы, и к концу диакинеза хромосомы остаются связанными только в теломерных участках.
В конце профазы-1 центриоли расходятся к полюсам клетки.
Метафаза-1. Завершается формирование веретена деления. Биваленты концентрируются в экваториальной плоскости клетки.
Анафаза-1. Гомологичные хромосомы расходятся к полюсам клетки. Каждая хромосома состоит из двух хроматид, соединенных общей центромерой.
Телофаза-1. Обычно очень короткая. У полюсов клетки группируются гаплоидные наборы хромосом, в которых представлен только один из парыгомологов. Восстанавливаются структура ядра и ядерная мембрана. Происходит частичная деспирализация хромосом. В конце телофазы-1 наступает цитокинез и образуются две клетки с гаплоидным набором хромосом.
После телофазы-1 вновь образованные клетки сразу вступают в мейоз-2, который проходит по типу обычного митоза.
Профаза-2. Частично деспирализованные хромосомы хорошо различимы. Начинается процесс обратной спирализации хромосом. Разрушается ядерная мембрана, формируется веретено деления, центриоли начинают расходиться к полюсам клетки.
Метафаза-2. Хромосомы выстраиваются в экваториальной плоскости. Центромеры прикрепляются к микротрубочкам образованного веретена деления.
Анафаза-2. Происходит разделение центромер, и каждая хроматида становится самостоятельной хромосомой. Дочерние хромосомы направляются к полюсам клетки.
Телофаза-2. Формируются новые ядра с гаплоидным набором хромосом. Хромосомы деконденсируются. Наступает цитокинез.
Основное биологическое значение мейоза заключается в обеспечении постоянства числа хромосом на протяжении поколений при половом размножении. Важным следствием мейоза является обеспечение генетического разнообразия гамет в результате рекомбинации хромосом и кроссинговера.
Механизм распределения неядерных генетических структур (митохондрий, хлоропластов) при митозе и мейозе пока неизвестен.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Мейоз и митоз
Мейоз и митоз Митоз – это деление клетки. Как известно, почти все клетки нашего организма время от времени делятся, но это не банальное деление пополам, а сложный многофазный процесс. Однако прежде чем говорить о митозе (и о другом варианте клеточного деления – мейозе),
3.5. Мейоз
3.5. Мейоз Современные представления о цитологических основах наследственности сформировались только после выяснения генетического смысла процесса мейотического деления клеток.Мейоз – это процесс образования гаплоидных клеток, т. е. клеток, имеющих половинный набор
Мейоз
Мейоз Мейоз – это процесс образования гаплоидных клеток, т. е. клеток, имеющих половинный набор хромосом. Примером гаплоидных клеток являются гаметы (половые клетки) и споры.Гамета – это клетка, способная объединяться с себе подобной клеткой с образованием зиготы –
20. Образование половых клеток. Мейоз
20. Образование половых клеток. Мейоз Вспомните!Где в организме человека происходит образование половых клеток?Какой набор хромосом содержат гаметы? Почему?Для осуществления полового размножения необходимы специализированные клетки – гаметы, содержащие одинарный