8.3.3. Пространственная структура популяций растений и животных
8.3.3. Пространственная структура популяций растений и животных
Типы распределения особей в пространстве. Занимаемое популяцией пространство предоставляет ей средства к жизни. Каждая территория может прокормить лишь определенное число особей. Естественно, что полнота использования имеющихся ресурсов зависит не только от общей численности популяции, но и от размещения особей в пространстве. Это наглядно проявляется у растений, площадь питания которых не может быть меньше некоторой предельной величины. Перехватывая корнями питательные вещества и воду, затеняя пространство, выделяя ряд активных веществ, каждое растение распространяет свое влияние на определенную площадь, поэтому оптимальным для популяции является такой интервал между соседними экземплярами, при котором они не влияют отрицательно друг на друга, но при этом не остается и недоиспользованного пространства.
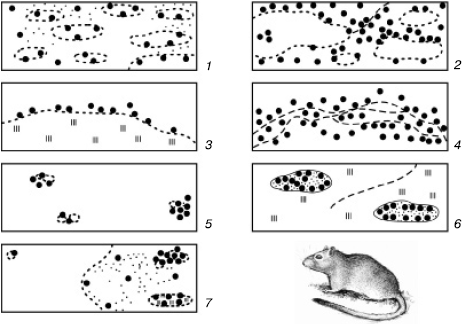
Рис. 104. Основные варианты размещения колоний больших песчанок (по Е. В. Ротшильду, 1966):
1– сплошные равномерные поселения; 2 – сплошные кружевные поселения; 3 – узколенточные; 4 – широколенточные; 5 – мелкоостровные; 6 – крупноостровные; 7 – отдельные скопления колоний
В природе изредка встречается почти равномерное упорядоченное распределение особей на занимаемой территории, например в уплотненных популяциях сидячих морских полихет, в чистых зарослях некоторых растений. Однако чаще всего члены популяции распределяются в пространстве неравномерно (рис. 104), что обусловлено двумя причинами: во-первых, неоднородностью занимаемого пространства, а во-вторых, некоторыми особенностями биологии видов, которые способствуют возникновению скоплений индивидуумов. У растений такая агрегированность возникает, например, при вегетативном размножении, при слабом распространении семян и прорастании их вблизи материнской особи; у животных – при групповом образе жизни семьями, стадами, колониями, при концентрациях для размножения, перезимовки и т. п.
Неравномерное размещение членов популяции может проявляться в двух крайних вариантах со всевозможными переходами между ними: 1) в резко выраженной мозаичности с незанятым пространством между отдельными скоплениями особей и 2) в распределении случайного, диффузного типа. Примером первого могут служить гнездовья грачей, поселяющихся колониями в рощах или парках, к которым примыкают благоприятные кормовые угодья. Диффузное распределение встречается в природе, если члены популяции относительно независимы друг от друга и обитают в однородной для них среде. Таково, например, размещение мучных хрущаков Tribolium confusum в муке, личинок-поденок в воде ручья, норок пауков-каракуртов на лугах и т. п.
В каждом конкретном случае тип распределения в занимаемом пространстве оказывается приспособительным, т. е. позволяет оптимально использовать имеющиеся ресурсы. Способы, которыми достигается рациональный характер размещения, определяются системой взаимоотношений между членами популяции.
Растения в ценопопуляции чаще всего распределены крайне неравномерно, образуя более или менее изолированные группы, скопления, так называемые микроценопопуляции, субпопуляции или ценопопуляционные локусы. Эти скопления отличаются друг от друга числом особей, плотностью, возрастной структурой, протяженностью. Часто более плотный центр скопления окружен особями, расположенными менее плотно.
Пространственная неоднородность ценопопуляции связана с характером развития скоплений во времени. У люцерны серповидной, например, семена обычно опадают в непосредственной близости от материнского растения, поэтому около обильно плодоносящих средневозрастных генеративных растений образуются скопления молодых (проростков, ювенильных и имматурных). Эти скопления характеризуются большой плотностью. По мере того как особи переходят в следующие возрастные состояния, скопления меняют возрастную структуру, изреживаются. Одновременно происходит приживление новых зачатков в пределах скопления, оно уплотняется, усложняется его структура, расширяется занятая им территория. Если приживление зачатков происходит за пределами скопления, то возникает новое. Скопления могут частично сливаться друг с другом, т. е. достигать более высокого уровня агрегированности.
Таким образом, жизнь ценопопуляции протекает в форме несинхронных возрастных изменений различных локусов, при этом меняется и ее пространственная структура, ибо изменяются конфигурация, протяженность локусов и их местоположение в фитоценозе.
У животных благодаря их подвижности способы упорядочивания территориальных отношений более разнообразны по сравнению с растениями. Даже у сидячих форм имеется ряд приспособлений к рациональному размещению в пространстве. У асцидий и мшанок растущий край колонии, наталкиваясь на колонию другого вида, растет поверх нее и в конечном счете подавляет ее. Но если встречаются колонии одного вида, каждая из них тормозит рост соседней и они начинают распространяться в другом направлении. При полном окружении колониями своего вида вегетативное размножение прекращается, но усиливается образование половых продуктов и подвижных личинок.
Личинки устриц, морских желудей, сидячих полихет, руководствуясь химическими стимулами, оседают обычно в местах, где уже имеются особи того же вида. Для осевшей личинки перед окончательным прикреплением характерно специальное поисковое поведение, позволяющее ей занять определенное место в скоплении. По наблюдениям, циприсовидные личинки морского желудя Balanus balanoides обычно прикрепляются на расстоянии не менее 2,5 мм от молодых и не менее 2 мм от старых особей своего вида. В то же время они свободно оседают в близком соседстве и даже на поверхности представителей других видов.
У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение – реакция на местонахождение других членов популяции. Инстинкты, поддерживающие размещение по территории отдельных особей или групп в популяциях, существуют у птиц, млекопитающих, пресмыкающихся, ряда рыб, в меньшей степени – у амфибий. Они выражены также у многих видов беспозвоночных со сложной нервной системой – ряда насекомых, пауков, крабов, осьминогов и др.
По типу использования пространства все подвижные животные делятся на две основные группы: оседлых и кочевых.
При оседлом существовании животное в течение всей или большей части жизни использует довольно ограниченный участок среды. Такие животные отличаются инстинктами привязанности к своему участку, а в случае вынужденного переселения – стремлением вернуться на хорошо знакомую территорию. Это «чувство дома» получило в экологии название «хоминг» (от англ. home– дом). Многие виды возвращаются к месту размножения даже после длительных и дальних миграций. Известно, например, что одна и та же пара скворцов из года в год может занимать «свой» скворечник. Хоминг голубей использовали даже в практических целях – для переноса почты.
Оседлый образ жизни имеет значительные биологические преимущества. На хорошо знакомой территории животное свободно ориентируется, тратит меньше времени на поиски корма, кратчайшим путем спасается в известные ему укрытия. Кроме того, многие оседлые виды создают систему запасов, прокладывают тропинки, строят дополнительные гнезда, норы, что помогает им выжить. Например, у белок есть основное гнездо, где выводится молодняк, и несколько дополнительных, в которые зверьки скрываются от непогоды или врагов. Белки создают также серии кладовых, запасая на зиму орехи, семена, грибы. Длительное использование определенной территории помогает более полному ее освоению.
На чужом участке поведение животного меняется. Наблюдения за сусликами показали, например, что такие зверьки отличаются суетливостью, неуверенностью передвижения, часто осматриваются, убежища находят лишь случайно и поэтому чаще гибнут, чем хозяева территории.
Уменьшение вероятности гибели от хищников на знакомой территории доказано и в экспериментах. Например, при выпускании ушастой совы в комнату, где находились хомячки, оказалось, что зверьков, ранее ознакомившихся с обстановкой комнаты, сова ловила в пять раз реже, чем тех, которые попадали в нее впервые.
Однако оседлый образ жизни таит в себе угрозу быстрого истощения ресурсов, если плотность популяции окажется слишком высокой. У оседлых видов выработались такие приспособительные особенности поведения, которые обеспечивают разграничение мест обитания между отдельными особями, семьями или другими внутрипопуляционными группировками (рис. 105). Общая площадь, занимаемая популяцией, оказывается поделена на отдельные индивидуальные или групповые участки, чем достигается упорядоченное использование запасов пищи, естественных укрытий, мест для размножения и т. п.
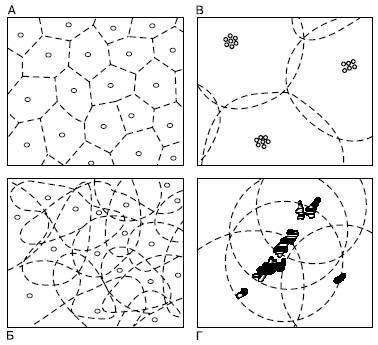
Puc. 105. Основные типы домовых участков у животных (по Н. П. Наумову, 1972):
А – одиночные разобщенные охраняемые;
Б – одиночные перекрывающиеся;
В – групповые разобщенные;
Г – групповые, перекрывающие друг друга (у колоний птиц на островах)
Территориальное поведение животных. Несмотря на территориальное обособление членов популяции, между ними поддерживается связь с помощью системы различных сигналов и непосредственных контактов на границах владений. Территориальное поведение животных включает два типа активности: 1) направленную на обеспечение собственного существования (поиск пищи, исследование территории, рытье нор и т. п.) и 2) на установление отношений с соседними особями (охрана участков, сигнализация, мечение и др.). Занимаемые участки не бывают больше тех пределов, при которых связь между соседними особями нарушается. Минимальные же размеры индивидуальной территории зависят от кормовых ресурсов среды. При такой системе размещения каждая оседлая особь не встречает конкуренции на своем участке, это повышает шансы на выживание и оставление потомства (рис. 106).
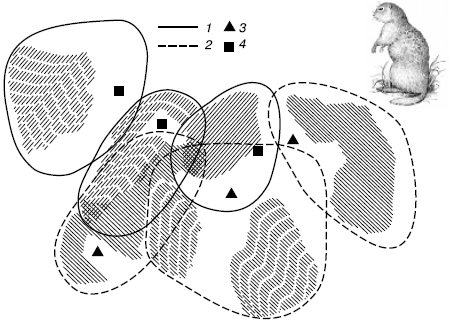
Рис. 106. Индивидуальные участки шести малых сусликов в Нижнем Поволжье (по А. Н. Солдатовой, 1955):
1– границы индивидуальных участков у самок; 2 – то же у самцов; 3 – гнездовые норы самцов; 4 – выводковые норы самок. Заштрихованы предпочитаемые каждым зверьком места кормежки
«Закрепление участка» достигается разными способами: 1) охраной границ занимаемого пространства и прямой агрессией по отношению к чужаку (рис. 107); 2) особым ритуальным поведением, демонстрирующим угрозу; 3) системой специальных сигналов и меток, свидетельствующих о занятости территории.
Прямая агрессия и нанесение конкуренту физических повреждений встречаются довольно редко. Перед нападением обычно демонстрируется поведение угрозы. Часто «схватка» представляет собой лишь систему ритуальных поз и заканчивается, как правило, изгнанием пришельца. В территориальных конфликтах в большинстве случаев побеждает хозяин участка, даже если ему противостоит более сильный соперник. В поведении животных на своей территории преобладает рефлекс активной обороны, на чужой – ориентировочный. Малый суслик немедленно бросается на другого, если тот забегает на его участок, и преследует нарушителя до границы своей территории. Если же, увлекшись погоней, он попадает на участок соседа, роли меняются и хозяин активно изгоняет зверька, от которого только что спасался сам.
Такое же право первого осуществляется и в территориальных притязаниях у большинства птиц. Гнезда мухоловок-пеструшек в эксперименте передвигали в переносных дуплянках на гнездовые участки соседних пар. При этом возникали драки, в которых всегда побеждали хозяева участка. Если гнездо победившей пары оказывалось на территории соседей, исход драки решался в пользу владельцев участка.
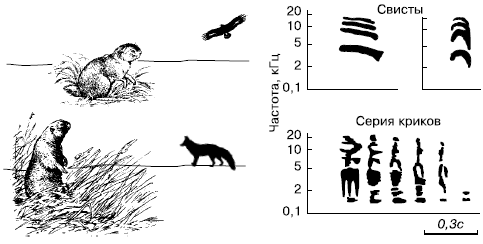
Рис. 107. Спектрограммы сигналов тревоги, издаваемых длиннохвостым арктическим сусликом. Сигналы слышны соседям (по А. И. Константинову, В. Н. Мовчану, 1985)
Многие животные вообще редко вступают в конфликты, активно избегая чужие участки. Широко развита система маркировки, предупредительных сигналов о занятости территории. У птиц это в основном сигнализация звуками. Поющий самец зарянки, например, все время передвигается по границам своего участка, задерживаясь в тех местах, откуда грозит вторжение соседа. Млекопитающие часто оставляют запаховые метки: мочу, кал, выделения специальных желез. У сурков и многих других видов для мечения участка служат анальные железы. У соболей пахучий секрет выделяется на подошвах ног, у серн – позади рогов, у антилопы гарна – спереди от внутреннего края глаз. Серны и антилопы оставляют запаховые следы на кустарниках и нижних ветвях деревьев. Собачьи, кошачьи и лемуры метят территорию мочой. Лемуры для этой цели ладонями втирают капли мочи в подошвы ног.
Ряд крупных животных оставляют метки, воспринимаемые зрительно (рис. 108). Например, медведи и дикие коты оставляют глубокие царапины на деревьях.
Обычная реакция на территориальные метки – избегание – закреплена у животных наследственно. Биологическая выгода такого типа поведения очевидна. Если бы овладение территорией решалось только исходом физической борьбы, появление каждого более сильного пришельца грозило бы хозяину потерей участка и устранением от размножения. Большая часть энергии особей тратилась бы на постоянные стычки, что уменьшило бы шансы выведения потомства и увеличило смертность.

Рис. 108. Зубр, маркирующий дерево (по И. И. Барабаш-Никифорову, А. Н. Формозову, 1963)
Охрана участка может осуществляться не на всей его территории, а лишь вблизи биологического центра – норы, гнезда и т. п. В таких случаях владения отдельных особей или групп могут перекрываться, иногда широко, и часть пространства животные используют совместно. На нейтральной территории инстинкт агрессии угасает. Так, мухоловки-пеструшки, гнезда которых искусственно сближают на границе соседних участков, мирно живут рядом и выкармливают птенцов.
Частичное перекрывание индивидуальных территорий служит способом поддержания контактов между членами популяции. Соседние особи часто поддерживают устойчивую обоюдовыгодную систему связей: взаимное предупреждение об опасности, совместную защиту от врагов (рис. 109). Нормальное поведение животных включает активный поиск контактов с представителями своего вида, который часто усиливается в период падения численности.
Территориальное поведение животных особенно ярко выражено в период размножения. Оно тесно связано с их физиологическим состоянием и регулируется гормональным путем. Вне сезона размножения у многих видов распределение по индивидуальным участкам сменяется групповым образом жизни с иным типом поведения. Большинство воробьиных птиц после вылета птенцов объединяются в стаи, совершающие кочевки.
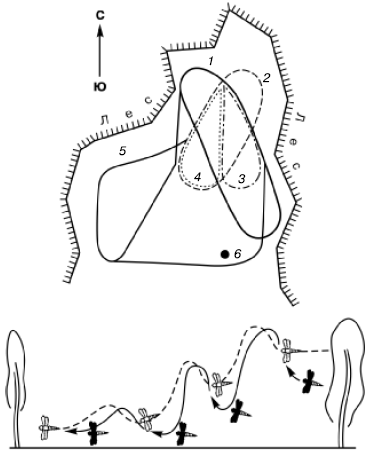
Рис. 109. Поведение стрекозы-коромысла на индивидуальном участке, ограниченном лесной поляной (по В. В. Заике, 1977): вверху – охотничий участок, внизу – самец-хозяин (светлый) изгоняет самца-нарушителя (темный), стараясь прижать его к земле.
1–5– разные траектории полета; 6 – одиночное дерево, избираемое местом отдыха
У ряда перелетных птиц территориальное поведение проявляется и на местах зимовок. Хищные и мелкие мышевидные грызуны обладают участками на протяжении всей жизни. Появившийся молодняк сначала остается на территории родителей, но затем выселяется, либо занимая освободившиеся участки, либо проникая в соседние популяции, либо формируя новые на незанятой территории.
В устойчивых семейных группах территориальные отношения между членами семьи могут быть достаточно сложными. Тигры, которым присуща полигамия, живут семьями, состоящими из самца, двух-трех самок и молодых разного возраста. Площадь семейной территории определяется плотностью основных жертв и при 14–15 особей копытных на 1000 га составляет около 50 тыс. га на семью (Лазовский заповедник). Территория, посещаемая самками, значительно меньше – около 10 тыс. га. Самец периодически обходит всю семейную территорию по постоянному маршруту, оставляя пахучие метки, экскременты и мочу на выступах скал, выворотах деревьев, нагребаемых кучах земли, снега и т. п. Участки молодых тигров частично перекрываются с территорией, посещаемой самками. Некоторые части семейных владений посещают лишь старые и молодые самцы. У каждого члена семьи есть излюбленные места для охоты. Через 3–5 лет молодые покидают семейную территорию родителей.
У оседлых видов все варианты общей пространственной структуры популяций можно свести к четырем основным типам: диффузному, мозаичному, пульсирующему и циклическому.
В популяциях диффузного типа животные распределены в пространстве дисперсно, не образуя обособленных поселений. Это возможно лишь при относительно равномерном размещении по территории кормов, мест, пригодных для размножения, и укрытий. Диффузный тип пространственной структуры популяций преобладает, например, у мелких млекопитающих сухих открытых пространств – большой песчанки и мохноногого тушканчика в монотонных песчаных ландшафтах, даурской пищухи в полынной и карагановой степи, малого суслика в различных растительных ассоциациях сухих степей и полупустынь.
Мозаичный тип размещения оседлых животных возникает в том случае, если пригодные для заселения биотопы распределены в пространстве резко неравномерно. Например, поселения малого суслика в волго-уральской полупустыне приурочены только к пятнам степной растительности, встречающимся среди сплошных песков. Обыкновенный хомяк в полупустыне обитает только в тростниковом поясе озер и на склонах озерных котловин.
Пульсирующий тип пространственной структуры характерен для популяций с резкими колебаниями численности. В годы депрессии популяция состоит из обособленных поселений, а в годы подъема численности занимает всю пригодную территорию, меняя мозаичный тип распределения на диффузный. В период резкого падения численности животные собираются на наиболее благоприятных участках, которые получили название «стаций переживания». Например, полевки-экономки в лесостепи в засушливые годы заселяют только березово-осиновые колки и заболоченные берега озер. Во влажные годы возникает обширная сеть временных поселений практически во всех биотопах, включая посевы. Эти временные поселения сохраняются обычно лишь до следующей засухи.
Пульсирующий тип структуры популяций можно отметить и у многих видов панцирных клещей и коллембол, заселяющих лесную подстилку (рис. 110). В сухое время эти членистоногие концентрируются лишь возле комлей деревьев, где режим влажности стабильнее, а во влажные периоды их популяции занимают весь покров растительных остатков на поверхности лесной почвы.
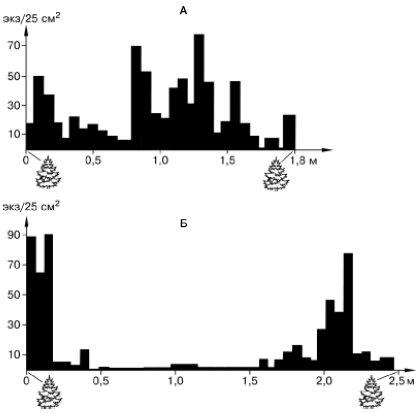
Рис. 110. Распределение панцирных клещей Nothrus silvestris в лесной подстилке между комлями двух елей в дождливый (А) и засушливый (Б) периоды. У комлей деревьев увлажнение более устойчиво
Циклический, или переложный, тип пространственной структуры популяций оседлых животных характеризуется закономерным попеременным использованием территории в течение года. Обский и копытный лемминги на острове Врангеля зимуют на сухих прибрежных возвышенностях. Летом зверьки переселяются на кочкарные и разнотравно-злаково-лишайниковые участки тундр, где и располагаются их летние норы. Лемминги отличаются высокой динамичностью пространственного распределения, отсутствием постоянных поселений, сменой мест зимнего и летнего обитания. Такой переложный характер использования территории в условиях тундр имеет большое приспособительное значение. Восстановление нарушенной растительности в тундрах происходит чрезвычайно медленно, длительное существование на одном месте за счет растительных кормов практически невозможно и грозило бы не только голодом зверькам, но и необратимыми изменениями растительного покрова. При переложном типе использования пространства сохраняется баланс между потреблением кормов и их ежегодным возобновлением, а тем самым и возможность существования дальнейших поколений.
Циклический тип освоения территории оседлыми животными, по существу, сходен с использованием ресурсов кочевыми популяциями.
Преимущества кочевого образа жизни заключаются в том, что животные не зависят от запасов корма на конкретной территории. Постоянные передвижения одиночных особей, однако, увеличивают вероятность гибели от хищников. Поэтому кочевой образ жизни практически несвойствен одиночным животным, кочуют группы: стада, стаи. Индивидуальные участки отдельных особей в таких группах не обособляются, но широко развита ориентация на соседей. В пасущихся стадах копытных между отдельными животными поддерживается определенное среднее расстояние, при котором они не мешают друг другу, обладая свободой передвижения. У птиц и рыб расположение в стае улучшает аэро– и гидродинамические условия полета и плавания: обеспечивается такое взаимодействие вихревых следов движения каждой особи, которое уменьшает общую величину сопротивления среды. У рыб, плывущих в середине стаи, частота движения хвостового плавника меньше, чем у тех, которые находятся впереди. С изменением скорости дистанция между членами движущейся стаи меняется.
Некоторые виды образуют широко кочующие группы, не привязанные к определенной территории. Таково поведение многих видов рыб во время нагульных миграций. Масштабы и длительность таких миграций зависят от обилия корма и численности стада. Чем крупнее стадо, тем длиннее его миграционный путь, так как быстрее потребляются встреченные корма. Норвежская сельдь Clupea harengus после нереста у берегов Норвегии кочует на тысячи километров на север и восток, держась в местах скопления планктона, который служит ей пищей.
В некоторых случаях, при резких нерегулярных изменениях запасов корма, возможны столь же нерегулярные перемещения популяции и временное «оседание» ее в тех местах, где запасы пищи еще не исчерпаны. Типичные примеры таких популяций без постоянных территорий можно обнаружить у клестов, всецело зависящих от неустойчивого урожая семян хвойных. У клестов постоянно меняются места гнездования и сроки размножения. В кормных лесах птицы выводят птенцов не только летом, но и зимой. Во внегнездовое время они широко кочуют в поисках лесов с богатым урожаем семян.
Перемещения многих других видов ограничены определенной территорией, которая может быть очень большой. Табуны зебр в Серенгети кочуют в сухой сезон на участке в 400–600 км2, во влажный общая площадь их перемещения занимает 300–400 км2. На этой территории у них есть определенные места отдыха, пастьбы и водопоев, которые они используют регулярно в определенные сроки. Периферические части таких территорий могут широко перекрываться с участками других стад. Регулярное перемещение по площади обычно происходит за сроки, достаточные для восстановления кормов на пастбищных участках. Сходным образом используют территорию стада других копытных, слонов, обезьян. Таким образом, при кочевом образе жизни животные совершают более или менее закономерные циклические перемещения по обширной территории с повторным использованием одних и тех же участков через определенные сроки.
У кочующих видов возможно также обособление оседлых популяций или временный переход к оседлому существованию. Северные олени на островах Северного Ледовитого океана зимуют там же, не делая попыток перехода на материк. Таймырские стада совершают обширные миграции более чем на 1000 км, но отдельные мелкие группы остаются на местах летнего нагула на северном Таймыре, как и на местах зимовок в лесотундре и северной части тайги.
Между разными способами использования территории нет абсолютных разграничений. Пространственная структура популяции очень динамична. Она подвержена сезонным и другим адаптивным перестройкам в соответствии с местом и временем. Однако масштабы возможных изменений и общий тип использования территории определяются биологическими особенностями вида, в первую очередь характером внутривидовых связей. Особенно большую роль в поддержании структуры популяций играет поведение особей по отношению друг к другу, система взаимных контактов.
Многочисленные конкретные формы проявления территориальных взаимоотношений животных можно расположить в постепенно усложняющийся ряд. Самая простая пространственная структура популяций – система неперекрывающихся охраняемых участков оседлых особей; более сложная – система перекрывающихся территорий, при которой усиливаются внутривидовые контакты. Наконец, групповое использование участков возможно только на основе строго упорядоченных отношений внутри группы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
ПРОИСХОЖДЕНИЕ ДОМАШНИХ ЖИВОТНЫХ И КУЛЬТУРНЫХ РАСТЕНИЙ
ПРОИСХОЖДЕНИЕ ДОМАШНИХ ЖИВОТНЫХ И КУЛЬТУРНЫХ РАСТЕНИЙ История органического мира с появлением человека приняла новые формы. Это воздействие человека на развитие органического мира выражается прежде всего в создании домашних животных и культурных растений.Первым
3.4. Отличия растений от животных
3.4. Отличия растений от животных Как считают большинство биологов, различия между растениями и животными можно разделить на три группы: 1) по структуре клеток и их способности к росту; 2) способу питания; 3) способности к движению. Отнесение к одному из царств проводится не
Происхождение растений и животных Три мира живой природы
Происхождение растений и животных Три мира живой природы Первые самые простые существа, или живые белки, которые образовались на земле, ещё нельзя было отнести ни к растениям, ни к животным. Эти живые белки возникали из более простых неживых белков, и за их же счёт
3. Учение об искусственном отборе (селекции) домашних животных и культурных растений
3. Учение об искусственном отборе (селекции) домашних животных и культурных растений Дарвин подчеркивает, что ему никогда не удалось бы раскрыть тайну изменения видов, если бы он не занялся сельским хозяйством, вопросом о выведении новых пород сельскохозяйственных
Сложные отношения всех животных и растений друг к другу в борьбе за существование.
Сложные отношения всех животных и растений друг к другу в борьбе за существование. Известно много случаев, показывающих, как сложны и неожиданны препятствия и отношения между органическими существами, которые должны бороться одновременно в одной и той же стране. Приведу
7. СТРУКТУРА И ДИНАМИКА СУБЪЕКТИВНОГО МИРА ЧЕЛОВЕКА И ЖИВОТНЫХ
7. СТРУКТУРА И ДИНАМИКА СУБЪЕКТИВНОГО МИРА ЧЕЛОВЕКА И ЖИВОТНЫХ 7.1. Историческая детерминация уровневой организации систем Представления о закономерностях развития многими авторами разрабатываются в связи с идеями уровневой организации (см. в [Анохин, 1975, 1980; Роговин, 1977;
7.2.2. Пространственная структура биоценоза
7.2.2. Пространственная структура биоценоза Тот участок абиотической среды, которую занимает биоценоз, называют биотопом, т. е., иначе, битоп – место обитания биоценоза (от лат. биос – жизнь, топос – место).Пространственная структура наземного биоценоза определяется
8.3. Биологическая структура популяций
8.3. Биологическая структура популяций Основные показатели структуры популяций – численность, распределение организмов в пространстве и соотношение разнокачественных особей.Индивидуальные черты каждого организма зависят от особенностей его наследственной программы
8.3.1. Половая структура популяций
8.3.1. Половая структура популяций Соотношение особей по полу и особенно доля размножающихся самок в популяции имеют большое значение для дальнейшего роста ее численности. У большинства видов пол будущей особи определяется в момент оплодотворения в результате
8.3.2. Возрастная структура популяций
8.3.2. Возрастная структура популяций С возрастом требования особи к среде и устойчивость к отдельным ее факторам закономерно и весьма существенно изменяются. На разных стадиях онтогенеза могут происходить смена сред обитания, изменение типа питания, характера
8.4. Этологическая структура популяций животных
8.4. Этологическая структура популяций животных Закономерности поведения животных составляют предмет особой науки – этологии. Систему взаимоотношений между членами одной популяции называют поэтому этологической или поведенческой структурой популяции.Поведение