3.1.4. Температурные адаптации гомойотермных организмов
3.1.4. Температурные адаптации гомойотермных организмов
Гомойотермия – принципиально иной путь температурных адаптаций, возникший на основе резкого повышения уровня окислительных процессов у птиц и млекопитающих в результате эволюционного совершенствования кровеносной, дыхательной и других систем органов. Потребление кислорода на 1 г массы тела у теплокровных животных в десятки и сотни раз больше, чем у пойкилотермных.
Основные отличия гомойотермных животных от пойкилотермных организмов: 1) мощный поток внутреннего, эндогенного тепла; 2) развитие целостной системы эффективно работающих терморегуляторных механизмов, и в результате 3) постоянное протекание всех физиологических процессов в оптимальном температурном режиме.
Гомойотермные сохраняют постоянный тепловой баланс между теплопродукцией и теплоотдачей и соответственно поддерживают постоянную высокую температуру тела. Организм теплокровного животного не может быть временно «приостановлен» так, как это происходит при гипобиозе или криптобиозе у пойкилотермных.
Гомойотермные животные всегда вырабатывают определенный минимум теплопродукции, обеспечивающий работу кровеносной системы, органов дыхания, выделения и других, даже находясь в покое. Этот минимум получил название базального метаболизма. Переход к активности усиливает выработку тепла и соответственно требует усиления теплоотдачи.
Теплокровным свойственна химическая терморегуляция – рефлекторное увеличение теплопродукции в ответ на понижение температуры среды. Химическая терморегуляция полностью отсутствует у пойкилотермных, у которых, в случае выделения дополнительного тепла, оно генерируется за счет непосредственной двигательной активности животных.
В противоположность пойкилотермным при действии холода в организме теплокровных животных окислительные процессы не ослабевают, а усиливаются, особенно в скелетных мышцах. У многих животных сначала наблюдается мышечная дрожь – несогласованное сокращение мышц, приводящее к выделению тепловой энергии. Кроме того, клетки мышечной и многих других тканей выделяют тепло и без осуществления рабочих функций, приходя в состояние особого терморегуляционного тонуса. При дальнейшем снижении температуры среды тепловой эффект терморегуляционного тонуса возрастает.
При продуцировании дополнительного тепла особенно усиливается обмен липидов, так как нейтральные жиры содержат основной запас химической энергии. Поэтому жировые запасы животных обеспечивают лучшую терморегуляцию. Млекопитающие обладают даже специализированной бурой жировой тканью, в которой вся освобождающаяся химическая энергия, вместо того чтобы переходить в связи ЛТФ, рассеивается в виде тепла, т. е. идет на обогревание организма. Бурая жировая ткань наиболее развита у животных – обитателей холодного климата.
Поддержание температуры за счет возрастания теплопродукции требует большого расхода энергии, поэтому животные при усилении химической терморегуляции либо нуждаются в большом количестве пищи, либо тратят много жировых запасов, накопленных ранее. Например, землеройка бурозубка крошечная имеет исключительно высокий уровень обмена. Чередуя очень короткие периоды сна и активности, она деятельна в любые часы суток и в день съедает корма в 4 раза больше собственной массы. Частота сердцебиения у бурозубок до 1000 в мин. Также и птицам, остающимся на зиму, нужно много корма: им страшны не столько морозы, сколько бескормица. Так, при хорошем урожае семян ели и сосны клесты зимой даже выводят птенцов.
Усиление химической терморегуляции, таким образом, имеет свои пределы, обусловленные возможностью добывания пищи. При недостатке корма зимой такой путь терморегуляции экологически невыгоден. Он, например, слабо развит у всех животных, обитающих за полярным кругом: песцов, моржей, тюленей, белых медведей, северных оленей и др. Для обитателей тропиков химическая терморегуляция также мало характерна, поскольку у них практически не возникает необходимости в дополнительном продуцировании тепла.
В пределах некоторого диапазона внешних температур гомойотермные поддерживают температуру тела, не тратя на это дополнительной энергии, а используя эффективные механизмы физической терморегуляции, позволяющие лучше сохранять или отводить тепло базального метаболизма. Этот диапазон температур, в пределах которого животные чувствуют себя наиболее комфортно, называется термонейтральной зоной. За нижним порогом этой зоны начинается химическая терморегуляция, за верхним – траты энергии на испарение.
Физическая терморегуляция экологически выгодна, так как адаптация к холоду осуществляется не за счет дополнительной выработки тепла, а за счет сохранения его в теле животного. Кроме того, возможна защита от перегрева путем усиления теплоотдачи во внешнюю среду.
Способы физической терморегуляции множественны. В филогенетическом ряду млекопитающих – от насекомоядных к рукокрылым, грызунам и хищникам механизмы физической терморегуляции становятся все более совершенными и разнообразными. К ним следует отнести рефлекторное сужение и расширение кровеносных сосудов кожи, меняющее ее теплопроводность, изменение теплоизолирующих свойств меха и перьевого покрова, противоточный теплообмен путем контакта сосудов при кровоснабжении отдельных органов, регуляцию испарительной теплоотдачи.
Густой мех млекопитающих, перьевой и особенно пуховой покров птиц позволяют сохранять вокруг тела прослойку воздуха с температурой, близкой к температуре тела животного, и тем самым уменьшить теплоизлучение во внешнюю среду. Теплоотдача регулируется наклоном волос и перьев, сезонной сменой меха и оперения. Исключительно теплый зимний мех млекопитающих Заполярья позволяет им в холода обходиться без существенного повышения обмена веществ и снижает потребность в пище. Например, песцы на побережье Северного Ледовитого океана зимой потребляют пищи даже меньше, чем летом.
У морских млекопитающих – ластоногих и китов – слой подкожной жировой клетчатки распределен по всему телу. Толщина подкожного жира у отдельных видов тюленей достигает 7–9 см, а общая его масса составляет до 40–50 % от массы тела. Теплоизолирующий эффект такого «жирового чулка» настолько высок, что под тюленями, часами лежащими на снегу, снег не тает, хотя температура тела животного поддерживается на уровне 38 °C. У животных жаркого климата подобное распределение жировых запасов приводило бы к гибели от перегрева из-за невозможности выведения избытка тепла, поэтому жир у них запасается локально, в отдельных частях тела, не мешая теплоизлучению с общей поверхности (верблюды, курдючные овцы, зебу и др.).
Системы противоточного теплообмена, помогающие поддерживать постоянную температуру внутренних органов, обнаружены в лапах и хвостах у сумчатых, ленивцев, муравьедов, полуобезьян, ластоногих, китов, пингвинов, журавлей и др. При этом сосуды, по которым нагретая кровь движется от центра тела, тесно контактируют со стенками сосудов, направляющих охлажденную кровь от периферии к центру, и отдают им свое тепло.
Немаловажное значение для поддержания температурного баланса имеет отношение поверхности тела к его объему, так как в конечном счете масштабы продуцирования тепла зависят от массы животного, а теплообмен идет через его покровы.
Связь размеров и пропорций тела животных с климатическими условиями их обитания была подмечена еще в XIX в. Согласно правилу Бергмана (1848), если два близких вида теплокровных животных отличаются размерами, то более крупный обитает в более холодном, а более мелкий – в теплом климате. Бергман подчеркивал, что эта закономерность проявляется лишь в том случае, если виды не отличаются другими приспособлениями к терморегуляции. Из проанализированных им 75 групп птиц в фауне Германии около трети удовлетворяло найденному правилу. Д. Лллен в 1877 г. подметил, что у многих млекопитающих и птиц северного полушария относительные размеры конечностей и различных выступающих частей тела (хвостов, ушей, клювов) увеличиваются к югу – правило Аллена. Терморегуляционное значение отдельных участков тела далеко не равноценно. Выступающие части имеют большую относительную поверхность, которая выгодна в условиях жаркого климата. У ряда млекопитающих, например, особое значение для поддержания теплового баланса имеют уши, снабженные, как правило, большим количеством кровеносных сосудов (рис. 18). Огромные уши африканского слона, маленькой пустынной лисички-фенека, американского зайца превратились в специализированные органы терморегуляции. При адаптации к холоду проявляется закон экономии поверхности, так как компактная форма тела с минимальным отношением площади к объему наиболее выгодна для сохранения тепла.

Рис. 18. Африканский длинноухий тушканчик (по В. Е. Соколову и др., 1977)
Если температура среды превышает верхнюю границу термонейтральной зоны, животным приходится затрачивать дополнительную энергию на испарительную терморегуляцию.
Эффективным механизмом отдачи тепла служит испарение воды путем потоотделения или через влажные слизистые оболочки полости рта и верхних дыхательных путей. Способность к образованию пота у разных видов очень различна. Человек при сильной жаре может выделить до 12 л пота в день, рассеяв при этом тепла в десять раз больше по сравнению с нормой. Выделяемая вода, естественно, должна возмещаться через питье. У некоторых животных испарение идет только через слизистые. У собаки, для которой одышка – единственный способ испарительной терморегуляции, частота дыхания при этом доходит до 300–400 вдохов в минуту. Регуляция температуры через испарение требует траты организмом воды и поэтому возможна не во всех условиях существования.
Поведенческие способы регуляции теплообмена для теплокровных животных не менее важны, чем для пойкилотермных, и также чрезвычайно разнообразны – от изменения позы и поисков укрытий до сооружения сложных нор, гнезд, осуществления ближних и дальних миграций.
В норах роющих животных ход температур сглажен тем сильнее, чем больше глубина норы. В средних широтах на расстоянии 150 см от поверхности почвы перестают ощущаться даже сезонные колебания температуры. В особенно искусно построенных гнездах также поддерживается ровный, благоприятный микроклимат (рис. 19). В войлокообразном гнезде синицы-ремеза, имеющем лишь один узкий боковой вход, тепло и сухо в любую погоду.
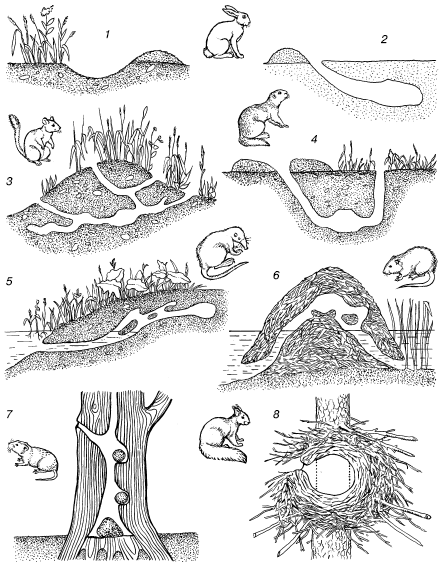
Рис. 19. Особенности строения нор и расположения гнезд разных видов млекопитающих (по Н. П. Наумову, 1963, И. И. Барабаш-Никифорову, Л. Н. Формозову, 1963):
1– логово зайца-русака в песчаных дюнах; 2 – снежная нора зайца-русака; 3 – летняя нора полуденной песчанки; 4 – нора малого суслика; 5 – нора выхухоли; 6 – хатка ондатры; 7 – гнезда рыжей полевки в дупле дуба; 8 – зимнее гнездо обыкновенной белки
В ряде случаев гомойотермные животные используют в целях терморегуляции групповое поведение. Например, некоторые пингвины в сильный мороз и бураны сбиваются в плотную кучу, так называемую черепаху. Особи, оказавшиеся с краю, через некоторое время пробиваются внутрь, и «черепаха» медленно кружится и перемещается. Внутри такого скопления температура поддерживается около +37 °C даже в самые сильные морозы. Обитатели пустынь верблюды в жару также сбиваются вместе, прижимаясь друг к другу боками, но этим достигается противоположный эффект – предотвращение сильного нагревания всей поверхности тела солнечными лучами. Температура в центре скопления животных равна температуре их тела, 39 °C, тогда как шерсть на спине и боках крайних животных нагревается до 70 °C.
Сочетание эффективных способов химической, физической и поведенческой терморегуляции при общем высоком уровне окислительных процессов в организме позволяет гомойотермным животным поддерживать свой тепловой баланс на фоне широких колебаний внешней температуры.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Эволюция механизмов адаптации
Эволюция механизмов адаптации Процесс приспособления, или адаптациогенез, составляет главное содержание эволюции. Он может идти на разных уровнях: строения организма, его поведения, структуры коллектива. На всех этих уровнях можно наблюдать постепенный переход от
Депрессия — часть процесса адаптации?
Депрессия — часть процесса адаптации? Депрессия, подобно гневу отверженности, может казаться нам непродуктивной. В чем же смысл мук и страданий, которые приносит нам потеря любимого? Неужели тратить энергию на рыдания выгоднее, чем активно заниматься ее
2.2. Адаптации организмов
2.2. Адаптации организмов Приспособления организмов к среде носят название адаптации. Под адаптациями понимаются любые изменения в структуре и функциях организмов, повышающие их шансы на выживание.Способность к адаптациям – одно из основных свойств жизни вообще, так как
3.1.1. Температурные границы существования видов
3.1.1. Температурные границы существования видов В среднем активная жизнедеятельность организмов требует довольно узкого диапазона температур, ограниченного критическими порогами замерзания воды и тепловой денатурации белков, примерно в пределах от 0 до +50 °C.Границы
3.1.3. Температурные адаптации пойкилотермных организмов
3.1.3. Температурные адаптации пойкилотермных организмов Температура пойкилотермных изменяется вслед за температурой окружающей среды. Они преимущественно эктотермны, выработки и сохранения собственного тепла у них недостаточно для противостояния тепловому режиму
Глава 4. ОСНОВНЫЕ СРЕДЫ ЖИЗНИ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ
Глава 4. ОСНОВНЫЕ СРЕДЫ ЖИЗНИ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ На нашей планете живые организмы освоили четыре основные среды обитания, сильно различающиеся по специфике условий. Водная среда была первой, в которой возникла и распространилась жизнь. В последующем живые
4.1. Водная среда обитания. Специфика адаптации гидробионтов
4.1. Водная среда обитания. Специфика адаптации гидробионтов Вода как среда обитания имеет ряд специфических свойств, таких, как большая плотность, сильные перепады давления, относительно малое содержание кислорода, сильное поглощение солнечных лучей и др. Водоемы и
Глава 10 Адаптации экспрессии генов в процессе развития
Глава 10 Адаптации экспрессии генов в процессе развития Жизнь -это сила, которая проделывает бесчисленное множество экспериментов, пытаясь организовать себя ... мамонт и человек, мышь и мегатерий, мухи и отцы церкви - все это результаты более или менее успешных попыток
Эмбриональные адаптации
Эмбриональные адаптации Эволюционная история любого вида определяет те его структуры и процессы, на которые может действовать естественный отбор. История и адаптация переплетаются друг с другом. Это привело к поразительному разнообразию личиночных морфологии. В
Идеалы и адаптации
Идеалы и адаптации Влияя на поведение живого существа, инстинкты влияют тем самым на его шансы выжить и оставить потомство, а следовательно — являются такими же адаптациями, как пищеварительная, кровеносная, терморегулирующая системы организма, и прочие приспособления
IV. Инстинкты адаптации к эволюционной среде обитания
IV. Инстинкты адаптации к эволюционной среде обитания Эволюционная среда обитания, она же — среда эволюционной адаптации, СЭА (в англоязычной литературе употребляется аббревиатура EEA) — среда, в которой происходила большая часть эволюции наших предков после их
10. Адаптации организмов к условиям обитания как результат действия естественного отбора
10. Адаптации организмов к условиям обитания как результат действия естественного отбора Вспомните!На основании собственных наблюдений приведите примеры приспособленности организмов к условиям существования.В течение многих веков в естествознании господствовало