Определение особенностей условнорефлекторной деятельности собак
Определение особенностей условнорефлекторной деятельности собак
Исследование высшей нервной деятельности собак во всех сериях опытов проводилось с помощью методики и аппарата, описанных в начале книги.
Создавая методику, мы исходили из того, что она должна была выявлять наиболее тонкие изменения в высшей нервной деятельности. Поэтому в качестве индикатора использовались сенсомоторные реакции собак, близкие по своему выражению к двигательным пищедобывательным (Воронин, 1957). Вырабатываемые рефлексы имели сложную эффекторную часть, требующую тонкой координации. Поэтому они были более подвержены изменениям, чем слюнные, электрокожные и кислотные. По своему выражению и процессу выработки получаемые рефлексы были ближе всего к так называемым произвольным движениям. По нашему мнению, это должно было способствовать выявлению у животных в период воздействия на них экстремальных факторов скрытых тонких изменений в реакциях центральной нервной системы.
Изучению рефлекторной деятельности предшествовало исследование у подопытных собак особенностей реакций их центральной нервной системы. Известно, что определение типа собак даже по малому стандарту испытаний в школе И.П. Павлова проходило на протяжении многих месяцев и включало в себя изучение процесса выработки условных рефлексов, исследование угашения с подкреплением и без подкрепления, переделку сигнальных раздражителей, пробы с применением фармакологических средств и т. д.
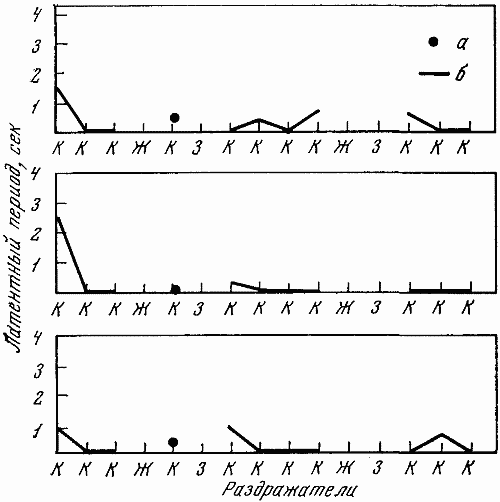
Рис. 22. Обследование высшей нервной деятельности путем трехкратной подачи выработанного стереотипа раздражителей
Собака Стрелка (2. VI 1960 г.); а, б — рефлексы на положительные сигналы
В наших условиях использование существующих лабораторных стандартов для определения типов было не применимо по ряду причин. Использование дрессировки лишало возможности охарактеризовать процесс выработки условных рефлексов. Угашение с подкреплением и без подкрепления и т. д. мы не могли применить, так как в каждый момент необходимо было иметь хорошо закрепленные реакции на стереотип раздражителей и т. д. В литературе (Паршутин и др., 1954; Флес, 1958) описывались приемы более простой и доступной оценки типологических особенностей животных. Но эти методы также не могли быть применены в наших условиях.
В связи со сказанным нам пришлось на основании принципов павловских функциональных проб продумать свою систему проверки особенностей высшей нервной деятельности собак. Оценочная роль такой системы не могла быть детально изучена. Поэтому целью исследований было охарактеризовать, хотя бы частично, процессы возбуждения, торможения и их баланса. Это позволяло сделать обоснованное заключение об отсутствии у подопытных животных патологических черт реакций центральной нервной системы.
Обследование нервной деятельности животных включало трехкратную подачу выработанного стереотипа, пробу для определения силы возбудительного процесса, две пробы для определения тормозного процесса и две — для испытания баланса возбудительных и тормозных процессов.
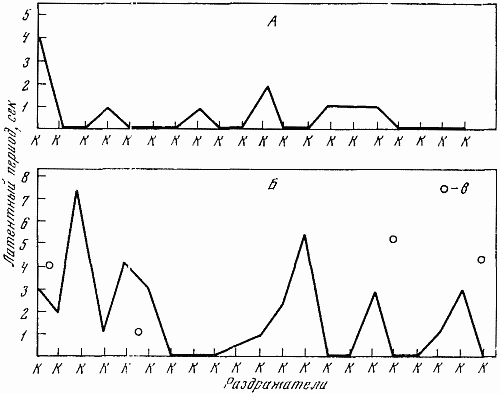
Рис. 23. Обследование процесса возбуждения путем подачи положительных раздражителей через укороченный в два раза интервал, т. е. через 15 сек.
А — Белка (10. V 1960 г.); Б — Звездочка (16. I 1960 г.); в — межсигнальные реакции
Трехкратная с интервалами в 10–17 мин. подача выработанного стереотипа раздражителей позволяла точнее оценить условнорефлекторный фон и провести статистическую обработку некоторых показателей реакции центральной нервной системы. У собаки Стрелки, отобранной для космического полета, такая подача раздражителей не вызывала нарушений условнорефлекторной деятельности, как это видно из рис. 22.
На рис. 22–26 показана условнорефлекторная деятельность подопытных собак при перечисленных выше пробах. Действие положительных сигналов обозначалось буквой «К», тормозных — «Ж» и «3» (свет большой и средней интенсивности), рефлексы на положительный раздражитель — точкой, на тормозной и срывы дифференцировок — кружочком, обведенным двумя линиями. Межсигнальные реакции обозначались кружочком, обведенным одной линией. Расположение этих обозначений по оси ординат примерно соответствует той секунде, на которой появлялись данные реакции.
Для определения силы возбудительного процесса — способности испытывать сильное возбуждение — использовалось многократное (20-разовое) повторение положительных сигналов при уменьшении интервалов между раздражителями (интервал в 30 сек. заменялся интервалом в 15 сек.). Проведение таких проб показано на рис. 23. Считалось, что животные обладают сильным возбудительным процессом, если уменьшение интервалов не сопровождалось запредельным торможением, существенным изменением латентного периода, появлением большого числа межсигнальных реакций.
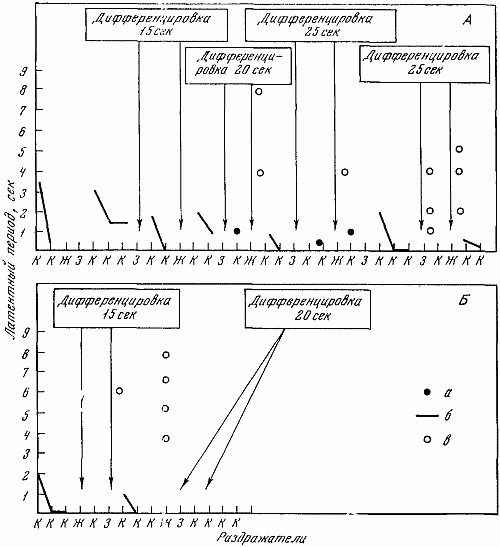
Рис. 24. Испытание тормозного процесса с помощью удлинения (до 15–25 сек.) дифференцированного тормозного раздражителя
А — Белка (7. V 1660 г.); Б — Жулька (18. Х 1960 г.). Остальные обозначения те же, что на рис. 22, 23
Сила тормозного процесса — способность выдерживать длительное концентрированное торможение — испытывалась с помощью традиционной для павловской школы пробы с удлинением до 15 и 25 сек. (вместо 10 сек.) действия тормозного раздражителя (рис. 24). В этом варианте раздражители подавались в том же порядке, что и обычно. Первые две дифференцировки — свет большой и средней интенсивности — удлинялись до 15 сек., а вторые две — до 20 сек. С целью охарактеризовать процесс торможения также применялась показанная на рис. 25 проба с удлинением до 60 сек. (вместо 30 сек.) в обычном стереотипе интервала перед тормозным раздражителем.
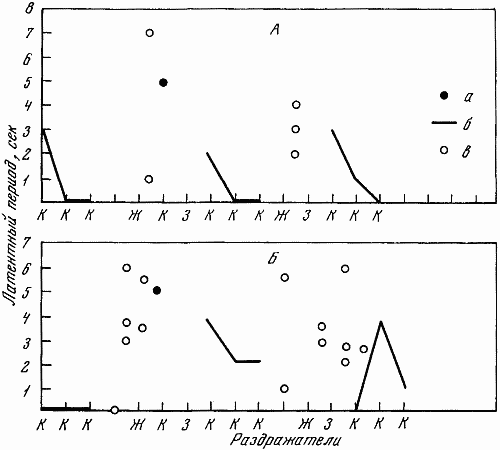
Рис. 25. Испытание тормозного процесса с помощью удлинения (до 60 сек.) интервала перед тормозным раздражителем
А — Жулька (18. Х 1960 г.); В — Звездочка (14. I 1961 г.). Остальные обозначения те же, что на рис. 22, 23
Для испытания баланса возбудительного и тормозного процессов использовалась проба с перемежающейся подачей положительных и тормозных раздражителей (рис. 26). В качестве дифференцировок, чтобы исключить сильно действующий свет, применялся только свет средней интенсивности. 25 раздражителей подавалось в следующем порядке: три положительных и дифференцировка, затем один положительный и дифференцировка, потом два положительных и дифференцировка, вслед за этим два раза подряд один положительный и дифференцировка, затем после положительного две дифференцировки, потом опять положительный и дифференцировка и 2 раза подряд положительный и две дифференцировки. Заканчивалась система подаваемых сигналов положительным раздражителем. Результаты обследования с помощью этой пробы показаны на рис. 26.
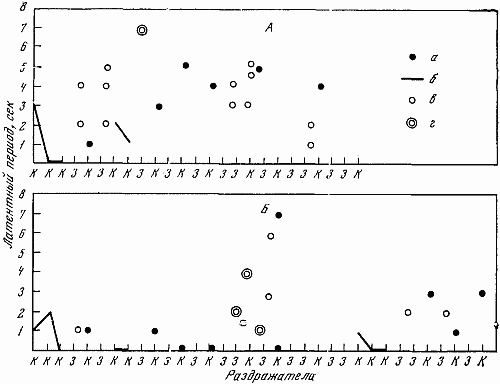
Рис. 26. Испытание баланса возбудительного и тормозного процессов путем перемежающейся подачи дифференцировочных и положительных раздражителей
А — Звездочка (13. II 1961 г.); Б — Жулька (15. VIII 1966 г.); г — срывы дифференцировки. Остальные обозначения те же, что на рис. 22, 23
Кроме приема с перемеживающейся подачей положительных и тормозных раздражителей, для испытания баланса нервных процессов применялась также проба с увеличенным интервалом (45 сек. вместо 30 сек.) Раздражители следовали в том же порядке, как и в обычном стереотипе. Увеличенные интервалы предваряли 3, 10 и 13–15-й раздражители. Перед 11-м раздражителем (тормозным большей силы) интервал был нормальным.
Опыты проводились на собаках, отобранных по характеру их поведения и высшей нервной деятельности. Брали животных, которые в клетках вивария и на прогулках в двигательном и голосовом отношениях вели себя спокойно. Исследование влияния на реакции центральной нервной системы перегрузок, вибраций, шума, космического полета проводилось на 12 собаках основной группы, выбранных из 37 животных; 25 животных было выбраковано в связи с тем, что у них при испытании особенностей условнорефлекторной деятельности были обнаружены случаи выпадений реакций в ответ на положительные сигналы, срывов дифференцировок, большие вариации в латентном периоде и т. д. В группу собак, отобранных для запуска в космос, вошли животные, условнорефлекторная деятельность которых заслуживала наиболее высоких оценок не только во время проб на возбудительный и тормозной процесс, но также и при испытании баланса возбудительного и тормозного процессов.
На основании полученных фактов можно было высказать предположение, что, например, собака Белка относилась к числу животных с сильными и уравновешенными нервными процессами.
Данные, полученные при троекратном обследовании выработанного стереотипа у всех 12 собак, показали общую устойчивость условнорефлекторного фона. Собаки реагировали на 96% подаваемых раздражителей. Латентный период в среднем был равен 1,4 при сигме 0,5. Сила нажима на педаль и направленность движений отвечали предъявляемым к ним требованиям в 99,6%. Неправильные нажимы (у Малька и Быстрого) были единичными случаями. У большинства собак дифференцировки были абсолютными.
Испытание возбудительного процесса показало, что собаки реагировали на 87% положительных сигналов. У семи из них было 100% правильных ответов, у пяти отмечалось выпадение небольшого числа реакций. Латентный период не изменялся или незначительно колебался, уменьшаясь и увеличиваясь (в среднем на 0,571 сек.). Укорочение интервала приводило к тому, что у половины собак межсигнальных реакций не было, у остальных они возникали в относительно небольшом числе.
При испытании тормозного процесса было сохранено 68,7% тормозных реакций. У трех собак они были абсолютными, остальные правильно реагировали на три, два и один тормозной раздражитель. У всех собак наблюдалось большое количество (в среднем 12) межсигнальных реакций. Такие реакции чаще появлялись после дифференцировок и особенно в том случае, когда два тормозных раздражителя следовали друг за другом. Удлинение дифференцировок незначительно влияло на положительные реакции: отмечалось выпадение 7,5% ответов на сигналы. Скрытый период реакций несущественно колебался.
Примерно такие же результаты были получены при испытании тормозного процесса с помощью другой пробы. Было сохранено 66,6% дифференцировок, т. е. всего на 2,1% меньше, чем при предыдущей. Регистрировалось выпадение 6% реакций на положительные и 27% на тормозные сигналы. Большое количество межсигнальных реакций наблюдалось в периоды удлиненных интервалов между отрицательными раздражителями и особенно, когда друг за другом следовало два тормозных сигнала.
Абсолютные дифференцировки и реакции на все положительные раздражители имели место у трех животных, у других трех они также были весьма благоприятными: собаки реагировали на 10 из 11 положительных сигналов и на 3–4 отрицательных из четырех. У следующих трех положительные реакции нарушались относительно мало, а дифференцировки — существенно. У Звездочки, наоборот, при полной сохранности реакций на дифференцировочные раздражители отмечалось выпадение реакций на положительный сигнал.
Все это позволяло сделать не только обоснованный вывод об отсутствии у подопытных животных патологических изменений в реакциях центральной нервной системы, но также и отнести Лисичку, Быстрого, Белку, Каму к собакам с сильными и уравновешенными процессами возбуждения и торможения. В нервной системе Невы, Малька и Пушинки, очевидно, при сильном раздражительном процессе обнаруживалась некоторая слабость тормозного. Она могла быть как результатом большей силы возбудительного процесса, так и слабости тормозного. Изменения латентного периода и число межсигнальных реакций в различных функциональных пробах давали возможность предположить у Невы первое, а у двух других животных — второе.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Наследование анатомических особенностей, связанных с привычкой сидеть на корточках
Наследование анатомических особенностей, связанных с привычкой сидеть на корточках В книге Habit and Heritage Вуд Джоунз обсуждал также манеру сидеть на корточках в разных популяциях человека. У многих азиатских народов принято сидеть на корточках так, что ступни стоят плоско
Предполагаемая недостаточность естественного отбора для объяснения начальных стадий полезных особенностей строения.
Предполагаемая недостаточность естественного отбора для объяснения начальных стадий полезных особенностей строения. Известный зоолог Ст. Джордж Майварт сделал недавно сводку всех возражений, когда-либо выдвинутых мною и другими против теории естественного отбора в
Причины, препятствующие приобретению полезных особенностей путем естественного отбора.
Причины, препятствующие приобретению полезных особенностей путем естественного отбора. М-р Майварт спрашивает затем (в этом и заключается его второе возражение): если естественный отбор столь могуществен и если способность обрывать сидящие высоко листья дает столь
Описание медико-психологических особенностей детей с проблемой гиперактивности
Описание медико-психологических особенностей детей с проблемой гиперактивности В 1970-х годах в Чехословакии проблемой так называемых гиперактивных детей (с двигательной расторможенностью) занималась группа чехословацких врачей, педагогов и психологов во главе с
Описание медико-психологических особенностей детей с ДЦП
Описание медико-психологических особенностей детей с ДЦП Детский церебральный паралич — заболевание незрелого мозга, которое возникает под влиянием различных вредных факторов, действующих в период внутриутробного развития, в момент родов и на первом году жизни. При
Описание медико-психологических особенностей детей с аутизмом
Описание медико-психологических особенностей детей с аутизмом В настоящее время аутизм рассматривается с точки зрения патологии развития. Больные аутизмом не понимают символического уровня, соответствующего их интеллектуальному возрасту. Человек с аутизмом
Описание медико-психологических особенностей детей с проблемой гиперактивности
Описание медико-психологических особенностей детей с проблемой гиперактивности В 1970-х годах в Чехословакии проблемой так называемых гиперактивных детей (с двигательной расторможенностью) занималась группа чехословацких врачей, педагогов и психологов во главе с
Описание медико-психологических особенностей детей с ДЦП
Описание медико-психологических особенностей детей с ДЦП Детский церебральный паралич — заболевание незрелого мозга, которое возникает под влиянием различных вредных факторов, действующих в период внутриутробного развития, в момент родов и на первом году жизни. При
Описание медико-психологических особенностей детей с аутизмом
Описание медико-психологических особенностей детей с аутизмом В настоящее время аутизм рассматривается с точки зрения патологии развития. Больные аутизмом не понимают символического уровня, соответствующего их интеллектуальному возрасту. Человек с аутизмом
Приложение III. Определение возраста собак Л. Г. Уткин
Приложение III. Определение возраста собак Л. Г. Уткин У собак, как и у других животных, возраст можно определить главным образом по зубам. Собака имеет 42 зуба, а именно: 12 резцов, 4 клыка, 2 вставных зуба в нижней челюсти и 24 коренных зуба. Зубная формула собаки выражается
Эволюционные истоки морфологических особенностей человека
Эволюционные истоки морфологических особенностей человека Как с эволюционной точки зрения интерпретировать переход к прямохождению, исчезновение волос на теле, нетипичное для обезьян распределение жировой ткани, стремительное увеличение размеров головного мозга и
Изучение собак Почему нужно изучать собак?
Изучение собак Почему нужно изучать собак? Биологический вид, к которому принадлежат домашние собаки, Canis familiaris, можно смело назвать успешным, даже чрезвычайно успешным.[1] Это означает, что, трансформировавшись по сравнению со своими предками, волками, они и теперь
2 Подготовка собак для поиска взрывчатых веществ, взрывных устройств, оружия и боеприпасов на основе игровой деятельности
2 Подготовка собак для поиска взрывчатых веществ, взрывных устройств, оружия и боеприпасов на основе игровой деятельности 2.1. Игровая потребность и игровая деятельность Потребность в игре, по мнению П. В. Симонова, относится к группе потребностей саморазвития [4] и
6. ВЫБОР МЕТОДИКИ И ТАКТИКИ ЛЕЧЕНИЯ В ЗАВИСИМОСТИ ОТ КЛИНИЧЕСКИХ ОСОБЕННОСТЕЙ, СТЕПЕНИ ТЯЖЕСТИ ОСНОВНОЙ И СОПУТСТВУЮЩЕЙ ПАТОЛОГИИ
6. ВЫБОР МЕТОДИКИ И ТАКТИКИ ЛЕЧЕНИЯ В ЗАВИСИМОСТИ ОТ КЛИНИЧЕСКИХ ОСОБЕННОСТЕЙ, СТЕПЕНИ ТЯЖЕСТИ ОСНОВНОЙ И СОПУТСТВУЮЩЕЙ ПАТОЛОГИИ Среди вышеописанных методик РДТ наиболее часто применяется полное (влажное) голодание, эту методику можно считать даже классической, ибо
РОЛЬ ОСОБЕННОСТЕЙ КУЛЬТУРЫ
РОЛЬ ОСОБЕННОСТЕЙ КУЛЬТУРЫ Разные авторы пришли к выводу, что ощущения, вызванные галлюциногенами, включают в себя ряд взаимодействующих особенностей, таких, как позиция, ожидания, мотивация, настроение, черты характера индивидуума, а также его физическое состояние.