Влияние ускорений на высшую нервную деятельность собак
Влияние ускорений на высшую нервную деятельность собак
Влияние ускорений на реакции центральной нервной системы изучались целым рядом отечественных и зарубежных исследователей. Опубликованные данные были обобщены в нескольких обзорах (Chambers, 1963; Климовицкий, Лившиц, Родионов, 1967; и др.).
Полученный материал говорил о существенных изменениях под влиянием ускорений психических реакций животных.
Ускорения +0,5g в течение 3 мин. снижали процент правильных реакций крыс (Herrick et al., 1958). Процент снижения таких реакций был существеннее при ускорении +2g, действующем от 1 до 5 мин. (De Marko, Geller, 1964). При поперечных ускорениях вида +2g в течение 30 мин. условный оборонительный рефлекс шимпанзе, когда нажим на рычаг прекращал болевое раздражение, увеличивался и, наоборот, уменьшался, когда ускорение становилось большим (+4g). У Барера (1962а, б) ускорения вида +4g в течение 15 сек. вызывали нарушения условных рефлексов, а когда величина ускорений достигала +7g и +10g в течение 1 мин., наблюдался полный срыв условнорефлекторной деятельности.
Для ответа на вопрос о глубине изменений реакций центральной нервной системы Барер (1962а, б) воспользовался методом исследования процесса восстановления. Восстановление условнорефлекторной деятельности крыс при действии на них ускорением вида +7g началось через 10–15 мин., а при +10g через 22–30 мин. Полностью животные отреагировали после ускорений вида +10g на подаваемый стереотип раздражителей только через 60 мин. Все это позволяло связать степень разрушения условнорефлекторной деятельности с силой воздействия ускорений. Такой вывод делал необходимым изучение реакций центральной нервной системы животных при величинах ускорений, ожидаемых в космическом полете.
Н.Н. Лившиц и Е.С. Мейзеров (1967) подробно исследовали характер нарушений реакций центральной нервной системы у крыс. Они установили, что нарушения заключаются в выпадении некоторых рефлекторных ответов на положительные раздражители, в удлинении латентного периода условных рефлексов, в снижении силы реакций на положительные раздражители. Некоторые из этих нарушений также были характерны для высшей нервной деятельности собак: снижались положительные условные рефлексы, нарушались индукционные отношения и т. д. (Кисляков, 1956).
В некоторых исследованиях были показаны резкие изменения рефлекторной деятельности в зависимости от характера подкрепляющих воздействий: болевых, кислотных, пищевых, питьевых (Herrick, 1961; и др.). Очевидно, разрушаемость и сохранность рефлексы, нарушались индукционные отношения и т. д.
О механизмах, лежащих в основе нарушений реакций центральной нервной системы под действием ускорений, можно думать, что они создавали предпосылки для повышения возбудимости нервной системы. Она проявлялась в таких неспецифических реакциях, как учащение дыхания, появление тахикардии. В электроэнцефалограмме регистрировалась десинхронизация, активация ?-ритма. Кроме того, усиливалась реакция экзальтации ?-ритма на закрывание глаз, увеличивалась амплитуда вызванных потенциалов в зрительной коре при стимуляции глаза мелькающим светом (Барер, 1962а, б; Изосимов, Разумеев, 1962а, б; Росин и др. 1959; и др.). Усиление возбудимости также доказывалось увеличением лабильности и проводимости рефлекторных дуг, что выражалось в значительных изменениях латентного периода условных рефлексов при относительно постоянной или несущественно изменяемой величине рефлексов. После периода повышения и ослабления возбудимости понижается не только возбудительный, но и тормозной процесс. Об этом свидетельствовало нарушение нормальных индукционных отношений (Кисляков, 1956), а также случаи ультрапарадоксальных реакций (Лившиц, Мейзеров, 1967). Нарушения возбудительного и активного тормозного процесса приводили к преобладанию запредельного тормозного. Развивалось состояние разлитого торможения, вызывающее иногда сон (Васильев, Герд, 1964).
В некоторых опытах наблюдался другой порядок развития всех этих явлений: условные рефлексы на первые раздражители отсутствовали, потом оказывались нормальными, в конце опыта опять регистрировалось их выпадение — циркуляторные нарушения нервной деятельности.
Можно говорить о наличии определенного параллелизма между изменениями гемодинамических показателей и некоторыми функциональными показателями реакций нервной системы. Значительные нарушения возникали в случаях, если направление ускорения совпадало с направлением основных кровеносных сосудов. Характеризуя общее действие ускорений на нервную систему, профессор Н.Н. Лившиц указывала, что ускорения вызывали резкие, но кратковременные сдвиги.
Дальнейшие исследования были проведены в плане отбора и подготовки собак к запускам в космос и входили как составная часть в более широкую группу изучения собак при действии на них ускорений (Котовская, Юганов, 1962).
Центростремительные ускорения переменной величины до +10g создавались на центрифуге радиусом 3,7 м и действовали в поперечном направлении к оси тела грудь — спина. Величины ускорений, скорость их нарастания и время действия воспроизводились соответственно графикам выведения космических кораблей на орбиту.
Опыты с воздействием перегрузок чередовались с контрольными, в которых сохранялись все условия эксперимента, за исключением вращения животных. Изучению реакций центральной нервной системы в связи с воздействиями ускорений предшествовало приучение животных находиться в фиксированном состоянии в кабине на центрифуге. Систему выработанных условных рефлексов проверяли непосредственно за 10–15 мин. до воздействия ускорений, через 5–7 мин. и через 25 мин. после центрифугирования. На рис. 27 показана собака Кама в кабине центрифуги.
Полученные данные показали, что при обследовании высшей нервной деятельности через 5–7 мин. после воздействия глубокие нарушения были обнаружены у трех собак: резкое удлинение скрытого периода рефлексов, выпадение большого числа ответных реакций.
Собаки Малек и Звездочка отреагировали более чем на половину подаваемых сигналов. При этом латентный период у них был замедлен относительно несущественно. Одно животное вначале подачи стереотипа раздражителей реагировало гораздо быстрее, чем обычно, а с пятого положительного сигнала, наоборот, его реакции удлинялись. Менее всего центрифугирование повлияло на количество ответных реакций и величину латентного периода у Невы: собака не отреагировала только на два сигнала, следующих за дифференцировкой. Однако у нее оказались значительно нарушенными дифференцировки. То же самое было характерно и еще для четырех собак.
Таким образом, полученный материал показал, что не только у разных собак, но и у одного и того же животного по одним показателям реакций центральной нервной системы регистрировалось явное и резкое ухудшение условнорефлекторной деятельности, а по другим, наоборот, реакции были нормальными (табл. 15). Следовательно, у собак под влиянием ускорения в одних случаях нарушался процесс возбуждения (выпадение рефлексов в ответ на положительный раздражитель, замедление латентного периода), в других — процесс торможения (срывы дифференцировок, появление большого числа межсигнальных реакций и т. д.).

Рис. 27. Собака Кама в кабине центрифуги
Собака одета в одежду, предохраняющую электроды для записи физиологических функций, и привязана для создания особым образом направленных перегрузок
Картина еще осложнялась различным характером изменений при первом и втором испытаниях. Через 25 мин. после центрифугирования у двух собак происходило дальнейшее нарушение корковой деятельности по всем показателям: они не реагировали на сигналы, безусловные раздражители были вялыми, в конце опыта животные засыпали. У одной собаки во время подачи стереотипа раздражителей период возбуждения, судя по межсигнальным реакциям, сменялся глубоким разлитым торможением (сном).
У трех животных на 25-й минуте после центрифугирования наблюдалось некоторое улучшение условнорефлекторной деятельности, скрытый период рефлексов уменьшался, собаки реагировали на большее, чем через 5 мин. после центрифугирования, число сигналов. Зато по другим показателям — реакциям на тормозные раздражители, наоборот, отмечалось ухудшение.
После воздействия перегрузок нарушался характер самих двигательных реакций. Животные (особенно Малек и Пушинка) в ответ на условный раздражитель стали нажимать на педаль не 1–2 раза, как обычно, а несколько раз; причем нажимы быстро следовали один за другим. Движения отличались беспорядочностью, лапы двигались не вдоль педали, а поперек ее и выше, иногда по панели со световыми сигналами. Нередко подобные движения были неадекватными по своей силе (таких нарушений не наблюдалось только у Невы).
Таблица 15. Число положительных условных реакций, дифференцировок и межсигнальных реакций у собак после воздействия перегрузок
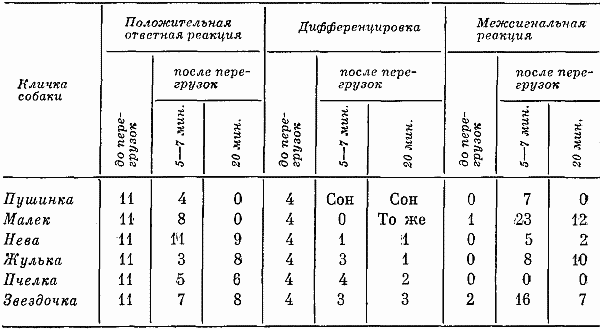
Изменения процессов, лежащих в основе реакций центральной нервной системы, очевидно, определялись характером сдвигов в области рефлекторной деятельности. Удлинение латентных периодов реакций и выпадение условных рефлексов в ответ на положительные раздражители свидетельствовали об ослаблении процесса возбуждения, а срывы дифференцировок и появление межсигнальных реакций — процесса торможения. О такой возможности в своем исследовании также говорит В.А. Кисляков (1956).
У Невы при относительной сохранности возбудительного процесса нарушался процесс внутреннего торможения. У Малька наряду с повышенной возбужденностью отмечалась быстрая истощаемость нервных процессов. Возбуждение этой собаки носило разлитой характер и захватывало ряд разных анализаторов: животное реагировало не только на световые, по и на звуковые сигналы, реакции его были особенно хаотичными, сочетались со стремлением освободиться, издавать звуки и т. д.
Реакции собак на отрицательные тормозные раздражители, появление межсигнальных реакций, хаотичное поведение — все свидетельствовало об ослаблении дифференцировочного, т. е. внутреннего торможения у многих животных. Однако эти же явления могли бы быть результатом развития в коре мозга пассивного разлитого торможения. Такое предположение подтверждалось вялостью некоторых собак. Значит, соотношение возбуждения и торможения изменялось в сторону преобладания торможения.
Для всех животных без исключения было характерно раннее нарушение внутреннего процесса торможения (резкое сокращение правильных реакций на дифференцировочные раздражители, увеличение числа межсигнальных реакций и т. д.). И.П. Павлов считал внутреннее торможение наиболее хрупким и нежным видом тормозных реакций.
Можно говорить об определяющей роли индивидуальных особенностей нервной системы собак. Нервная система Невы мало подвергалась влиянию перегрузок. У трех собак изменения по типу охранительного торможения наступали при первом обследовании, при втором у них отмечалась тенденция к восстановлению условнорефлекторных реакций. У Пушинки и Малька охранительное торможение развилось лишь к 25-й минуте после центрифугирования. Полной нормализации деятельности коры головного мозга в указанные сроки наблюдения ни у одного животного не наступило.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Деятельность риккетсий
Деятельность риккетсий Риккетсии принадлежат к числу самых заклятых врагов человека. Их активными помощниками бывают паразитирующие насекомые: вши, блохи, клещи. Мы знаем о риккетсиях всего каких-то 50 лет, хотя уже с XVI века известно, что они сеяли смерть по всей Европе. И
Орудийная деятельность
Орудийная деятельность Давно прошли те времена, когда изготовление и использование орудий считались уникальными свойствами человека. Сегодня известно множество видов животных, использующих орудия в повседневной жизни, причем в ход идут как неизмененные природные
10.5. Орудийная деятельность животных
10.5. Орудийная деятельность животных Именно изучение орудийной деятельности человекообразных обезьян положило начало проблеме мышления у животных. С развитием этологии список видов, к которым приложимо понятие орудийной деятельности, постоянно расширялся. Среди
4. ВЛИЯНИЕ ЭМОЦИЙ НА ДЕЯТЕЛЬНОСТЬ И ОБЪЕКТИВНЫЕ МЕТОДЫ КОНТРОЛЯ ЭМОЦИОНАЛЬНОГО СОСТОЯНИЯ ЧЕЛОВЕКА
4. ВЛИЯНИЕ ЭМОЦИЙ НА ДЕЯТЕЛЬНОСТЬ И ОБЪЕКТИВНЫЕ МЕТОДЫ КОНТРОЛЯ ЭМОЦИОНАЛЬНОГО СОСТОЯНИЯ ЧЕЛОВЕКА Сам факт генерирования эмоций в ситуации прагматической неопределённости предопределяет и объясняет их адаптивное компенсаторное значение. Дело в том, что при
2. ОРИЕНТИРОВОЧНО-ИССЛЕДОВАТЕЛЬСКАЯ ДЕЯТЕЛЬНОСТЬ
2. ОРИЕНТИРОВОЧНО-ИССЛЕДОВАТЕЛЬСКАЯ ДЕЯТЕЛЬНОСТЬ Ориентировочную реакцию (как настройку анализаторов на лучшее восприятие нового стимула) следует отличать от исследовательских реакций и ориентировочно-исследовательского поведения. Последние могут следовать за ОР –
Изучение собак Почему нужно изучать собак?
Изучение собак Почему нужно изучать собак? Биологический вид, к которому принадлежат домашние собаки, Canis familiaris, можно смело назвать успешным, даже чрезвычайно успешным.[1] Это означает, что, трансформировавшись по сравнению со своими предками, волками, они и теперь
Влияние на собак 7-часового пребывания в камере
Влияние на собак 7-часового пребывания в камере Вегетативные показатели: после трех-четырехразового пребывания в 7-часовых опытах наблюдалось снижение пульса (на 5–11 ударов в минуту), числа дыхательных движений в минуту (на 2–3), уменьшалась (на 0,03°) температура тела.
Влияние на собак 12-часового пребывания в камере
Влияние на собак 12-часового пребывания в камере Состояние животных при опытах такой длительности изучалось после первого и второго 12-часового опыта, а также при последних (9–11-м) опытах такой длительности. Последнее позволяло судить о приученности животных находиться в
Влияние на собак 19-часового пребывания в камере
Влияние на собак 19-часового пребывания в камере Вегетативные показатели. В первых 19-часовых опытах, начиная с 12-го часа, когда в предыдущих экспериментах собаки выпускались из кабины и бежали на прогулку, частота пульса и дыхательных движений увеличивалась или
Влияние на высшую нервную деятельность собак вибраций
Влияние на высшую нервную деятельность собак вибраций Вибрации даже в относительно небольших дозах, по мнению ряда исследователей, являлись сильным раздражителем для центральной нервной системы (Андреева-Галанина, 1956; Лукьянова, 1964; и др.).У собак действие местных
Влияние на реакции центральной нервной системы собак запусков в космос
Влияние на реакции центральной нервной системы собак запусков в космос Изучение реакции центральной нервной системы в экстремальных условиях проводилось отечественной физиологией в работах школы И.П. Павлова. Внимание таким исследованиям уделяли и зарубежные
Влияние запуска в космос на поведение и психическое состояние собак
Влияние запуска в космос на поведение и психическое состояние собак Поведение и психическое состояние собак может быть использовано как наглядный индикатор общего состояния живых организмов в необычных условиях и в том числе при полетах на космических кораблях, при
2.1. Деятельность пищеварительного аппарата
2.1. Деятельность пищеварительного аппарата Процессы экзотрофии у большинства животных и человека реализуются пищеварительной системой. На протяжении XIX в. было дано множество технологических описаний этой системы. Одно из самых ярких в 1897 г. сделал И.П. Павлов, который
ПРИРОДООХРАННАЯ ДЕЯТЕЛЬНОСТЬ
ПРИРОДООХРАННАЯ ДЕЯТЕЛЬНОСТЬ В 1979 году в Англии окончательно исчезла большая голубая бабочка (Large Blue butterfly), количество особей которой в течение предшествующих десятилетий неуклонно сокращалось. Детальные исследования выявили причину сокращения численности этого вида,
Рассудочная деятельность
Рассудочная деятельность Экспериментальное изучение рассудочной деятельности на животных практически только начинается. Несмотря на то, что Ч. Дарвин еще в 1871 г. с совершенной определенностью высказал свое мнение о том, что животные обладают «рассуждающей