Физиолого-генетические проблемы сложных форм поведения в норме и патологии[46]
Физиолого-генетические проблемы сложных форм поведения в норме и патологии[46]
Поведение высших позвоночных животных и человека чрезвычайно гетерогенно. С. Н. Давиденков (1947) указывал на то, что вопрос о «вариациях нормы» высшей нервной деятельности человека и отношение этих нормальных вариаций к патологии высшей нервной деятельности является основным положением невропатологии. На чрезвычайно большую гетерогенность как нормального, так и патологического поведения указывает также Оливерио (1975).
Генетика поведения является той наукой, в которой соприкасаются важнейшие проблемы ряда крупных дисциплин: генетики, физиологии, биохимии, биологии развития, эволюционного учения. Генетика поведения, без которой невозможно изучать гетерогению поведения, является тем фундаментом, который лежит в основе невропатологии и психиатрии. Одной из важнейших задач, стоящих перед генетикой поведения, является объективное экспериментальное изучение рассудочной деятельности.
И. М. Сеченов настоятельно указывал на то, что при изучении основ мышления необходимо исследовать простейшие психические акты у животных.
Задача настоящей статьи — рассмотреть роль центральных механизмов нервной системы в проявлении и выражении нормальных и патологических особенностей поведения.
Метод исследования — детальное изучение феногенетики признаков поведения с выделением элементарных поведенческих реакций и основных физиологических функций нервной системы, во взаимодействии которых осуществляется формирование поведения.
Сформулированная нами ранее гипотеза (Крушинский, 1938, 1960, 1977) и на сегодняшний день способствует пониманию интеграции нормального и патологического поведения как животных, так и человека. Конкретно эта гипотеза заключается в следующем: проявление и выражение любых генов, принимающих участие в формировании как нормальных, так и патологических поведенческих актов тем выше, чем выше общая возбудимость нервной системы; механизмом, препятствующим проявлению и выражению как нормальных, так и патологических признаков, является тормозной процесс.
Возбуждение и торможение контролируется генотипом и находится в относительно простых эпистатических отношениях.[47] Результатом их взаимодействия определяется уровень возбудимости.
Положением о роли возбудимости в проявлении и выражении любых признаков поведения можно, по-видимому, объяснить, почему талантливые люди и их родственники нередко страдают различными формами нервно-психических заболеваний. Это может происходить потому, что в семьях повышенно возбудимых людей любые особенности поведения, как нормальные, так и патологические должны иметь наиболее яркую форму своего выражения.
Проблема генетической связи между талантливостью и нервно-психическими заболеваниями, обсуждаемая в течение нескольких десятилетий (Геккер, 1924; Ганушкин, 1934; Эфроимсон, 1976), до настоящего времени не получила экспериментального физиолого-генетического развития.
В основу настоящей работы положены результаты планомерного феногенетического анализа нормальных и патологических признаков поведения.
Отправной точкой экспериментального исследования послужила попытка объяснить причину появления чрезвычайно трусливого потомства при скрещивании двух пород собак: немецких овчарок и восточносибирских (гиляцких) лаек, которые сами не были трусливыми. Исследования показали, что в основе такого гетерозиса лежит комбинация повышенной возбудимости (передающейся от немецких овчарок) с генетически обусловленной трусостью, которая не проявлялась у гиляцких лаек. При повышении возбудимости в результате инъекций кокаина эти собаки делались трусливыми (Крушинский, 1938).
Дальнейшие исследования, проводимые на собаках и крысах, показали, что уровень возбудимости является весьма существенным фактором в проявлении и выражении различных актов поведения, которые могут быть отнесены к категории безусловных рефлексов (инстинктов). Чем выше возбудимость, тем больше вероятность проявления различных особенностей поведения в фенотипе. Было показано, что акты поведения, формирующиеся под ведущим влиянием обучения, также в значительной степени зависят от уровня возбудимости нервной системы.
В наших экспериментах одним из показателей возбудимости служила двигательная активность собак. Была обнаружена положительная корреляция между успехом обучения и уровнем двигательной активности собак (Крушинский, 1960).
Большое значение в проявлении и выражении признаков поведения имеет сила нервной системы. Под силой нервной системы понимается «работоспособность нервной системы», определяемая пределом возбуждения нейрона мозга и, как показали наши исследования, возможностью тормозного процесса купировать застойное возбуждение (Крушинский, 1960).
В Институте физиологии им. И. П. Павлова нами была проведена обработка данных по исследованию силы нервной системы у собак. Анализ результатов скрещивания, а также вычисление коэффициентов корреляции между силой нервной системы братьев и сестер (r=+0,34±0,10) показал генотипическую обусловленность силы нервной системы. Уровень силы тормозного процесса как одного из компонентов возбудимости нервной системы связан с проявлением некоторых генотипически обусловленных форм поведения. Так, например, собаки, обладающие слабым типом нервной системы, в подавляющем большинстве (15:1) были трусливыми (Крушинский, 1947).
Сила нервной системы имеет значение и при выработке индивидуально приобретенных навыков. При легких условиях обучения (когда дни обучения чередуются с днями отдыха) между успехом обучения и силой нервной системы не наблюдалась положительная корреляция. Однако в более трудных условиях удавалось обнаружить положительную корреляционную связь.
Таким образом, на первом этапе проводимого нами исследования собранный фактический материал не опровергал гипотезу о значении уровня возбудимости и силы тормозного процесса в проявлении и выражении нормальных поведенческих актов, какими являются инстинкты и условные рефлексы.
Экспериментальное изучение роли возбуждения и торможения в патологии нервной деятельности было проведено нами на модели рефлекторной (или аудиогенной) эпилепсии крыс.
В результате селекции[48] нам с Л. Н. Молодкиной удалось вывести популяцию крыс (КМ), у которых практически в 100 % при действии звукового раздражителя развиваются интенсивные судорожные припадки (вместо 10–15 % у исходной популяции). Популяция крыс КМ дала возможность изучать в модельных опытах разные патофизиологические признаки: аудиогенную подкорковую эпилепсию, миоклонические корковые судорожные припадки и нарушения сердечно-сосудистой системы (с летальными исходами) при действии звуковых раздражителей.
Методом диаллельного скрещивания было показано, что высокая чувствительность крыс к действию звукового раздражителя обусловливается системой рецессивных генов (Романова, 1975).
Проведенное исследование различных патологий на полученной нами генетической модели подтвердило высказанную гипотезу о значении возбуждения и торможения в проявлении и выражении ряда тяжелых функциональных патологий мозга и сердечно-сосудистых нарушений. При звуковой эпилепсии крыс первым симптомом патологического процесса является бурное двигательное возбуждение, которое заканчивается судорожным припадком. Применение фармакологических препаратов, вызывающих возбуждение (кофеин, стрихнин), оказывало однозначный эффект: сокращение латентного периода двигательного возбуждения и усиление интенсивности судорожного припадка. Применение транквилизаторов и седативных средств (бром, люминал и др.) оказывало обратное действие.
Электрофизиологическое исследование показало наличие резких изменений биопотенциалов в коре и подкорковых структурах мозга. Патологическое возбуждение при судорожном припадке начинается в продолговатом мозге, откуда оно распространяется по специфическим (слуховым) и неспецифическим путям в верхние отделы центральной нервной системы. В коре больших полушарий патологическая активность регистрируется только во время миоклонических судорог, которые появляются после многократных звуковых воздействий. Проведенный физиолого-генетический анализ показал значение взаимодействия между возбуждением и торможением в формировании припадков рефлекторной эпилепсии, которое обусловливает «фенотипический узор» патологического процесса.
В развитии судорожного комплекса экспериментальной эпилепсии у крыс выступают основные компоненты феногенетического процесса, выделенного Н. В. Тимофеевым-Ресовским (1930): проявление, выражение и специфичность.
Важнейшим этапом в проводимой нами работе была селекция крыс линии КМ на затяжное возбуждение, которое продолжается после выключения звука в течении 20–30 мин. Когда путем селекции была выведена линия крыс с затяжным возбуждением, у них обнаружилась значительная гетерогения в реакции на применяемые звуковые раздражители. Эта гетерогения поведения выражалась в появлении фазовых состояний нервной системы. В норме реакция организма на внешний раздражитель подчиняется закону силовых отношений: в ответ на сильное раздражение возникает сильная реакция, на слабый — слабая. При резком возбуждении мозга закон силовых отношений перестает действовать и появляются новые отношения между раздражителями и ответной реакцией организма (Крушинский и др., 1952).

Рис. 1. Парадоксальная стадия при регистрации импульсной активности нейрона продолговатого мозга
А — увеличение импульсации на звук при полутороминутной экспозиции; Б, В — силовые; Г, Д — парадоксальные отношения

Рис. 2. Кривые, характеризующие изменения процессов возбуждения и торможения и их взаимодействующей
И. П. Павлов, обнаружив это явление на собаках, назвал его запредельным торможением и рассматривал в качестве защитной реакции нервной системы. Крупный отечественный психиатр В. Л. Осипов посредством запредельного торможения объяснял симптомокомплекс ряда тяжелых психических заболеваний человека, в том числе шизофрении (Ленц, 1925).
Полученная на крысах генетическая модель запредельного торможения открыла практически неограниченные возможности в изучении закономерностей развития перевозбуждения мозга и тех защитных механизмов, которые используются нервной системой против его вредоносного действия. Феногенетическое описание запредельного торможения на поведенческом уровне у крыс подтвердилось при изучении суммарной и импульсной активности нейронов продолговатого мозга (Семиохина, Цурита, 1974) (рис. 1). Можно предположить, что все многообразие фаз запредельного торможения, полученное на генетической модели, сводится к простым количественным изменениям взаимоотношения между процессами возбуждения и торможения во времени.
Нами была высказана гипотеза, что многообразие фаз запредельного торможения может быть объяснено при допущении, что возбуждение по мере действия стимулирующего раздражителя нарастает в линейной зависимости от времени, а торможение, индуцируемое возбуждением, — по экспоненте (рис. 3). Возбудимость мозга, которая определяет все многообразие обнаруженных фаз запредельного торможения, выступает как разность уровней возбуждения и торможения. Проверка предполагаемых взаимоотношений между возбуждением и торможением была осуществлена при помощи построения математической модели Г. В. Савиновым (1962). Технически построенная модель была осуществлена на электронной машине.
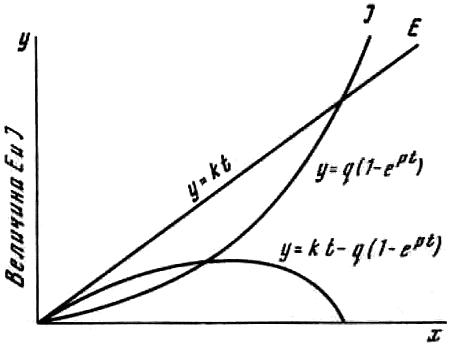
Рис. 3. Осциллограмма, полученная на модели. Воспроизведение всех фаз парабиоза, обнаруженных в опытах со звуковой экспозицией крыс
Стадии: I — силовая; II — уравнительная; III — парадоксальная; IV — затяжное возбуждение; V — инвертированная
На блоке усилителя набрана нелинейная характеристика, которая соответствует кривой изменения возбудимости во времени (см. рис. 3). На осциллограмме, полученной на модели (рис. 2), описанная схема воспроизводит зарегистрированные в опыте на животных взаимоотношения между раздражителем и ответной реакцией организма. Полученные данные с построением математической модели изменения функционального состояния системы по времени хорошо объясняют гетерогению проявления и выражения нормальных и патологических реакций поведения в фенотипе и находятся в соответствии с высказанной рабочей гипотезой.
Нет необходимости говорить здесь о биологическом значении описанной системы саморегуляции. Она, несомненно, весьма целесообразна. Пока эта система работает сбалансированно, опасности для жизни организма практически нет. Однако когда эта сбалансированность нарушается, резкое возбуждение мозга может привести к летальному исходу. В наших опытах это достигалось в тех случаях, когда после звуковой экспозиции делался перерыв. В этом случае 12 %подопытных животных гибнут от острых нарушений кровообращения. Почти у всех погибших особей наблюдаются тяжелые кровоизлияния в мозг (Крушинский, Флесс, Молодкина, 1952; Крушинский, Пушкарская, Молодкина, 1953). Если перед началом звуковой экспозиции повысить возбудимость нервной системы введением кофеина или тироксина, летальные исходы значительно учащаются (Крушинский, Доброхотова, 1957) (табл. 1). И наоборот, если звуковую экспозицию проводить в камере с повышенным содержанием СO2, который снижает возбудимость, летальных исходов не бывает (Крушинский и др., 1960). На этом примере хорошо видно, какое значение имеет повышенная возбудимость для течения патологического процесса: она является тем фоном, на котором проявляются самые различные патологии, связанные с функциональным состоянием нервной системы.
Таблица 1. Влияние гипертиреоидизации на частоту смертей при нервной травме

Экспериментальное изучение каких-либо генетических или физиологических проявлений талантливости в модельных опытах на животных невозможно. Однако животные, так же как и человек, обладают элементарной рассудочной деятельностью, уровень развития которой является важнейшим компонентом талантливости.
Наша лаборатория в течение 25 лет изучает объективными физиологогенетическими методами биологические основы рассудка в экспериментах на животных. Предпосылкой для проведения таких исследований послужило то, что нам удалось определить наиболее существенные черты рассудочной деятельности. Мы считаем, что поведенческий акт может быть оценен как рассудочный, если он выполняется адекватно в новой обстановке в результате оперирования эмпирически уловленными законами, связывающими явления и предметы внешнего мира.
Нам удалось выделить три формы поведения, которые выполняются как человеком, так и животными на основе оперирования эмпирически уловленными законами природы: первая связана со способностью к экстраполяции (Крушинский, 1958); вторая — с оперированием пространственными отношениями между геометрическими фигурами (Крушинский, 1968); третья — с определением закономерности перемещения объекта в пространстве (Крушинский, Попова, 1978).
В результате проведенных исследований удалось изучить отдельных представителей всех классов позвоночных животных, от рыб до обезьян и дельфинов, и начать исследования на людях.
Исследование показало несомненную роль генотипических факторов в полиморфизме по рассудочной деятельности. Об этом говорят исследования, проведенные на крысах (Крушинский, 1977а). При сравнении лабораторных крыс с пасюками и гибридами, несущими долю генотипа пасюков, наблюдаются ясные различия в успехе решения предъявляемого теста и развитие неврозов после правильного решения задачи.
Удобным объектом для изучения генетики поведения являются мыши различных инбредных линий. Нашей лабораторией совместно с отделом эмбриологии ИЭМ АМН СССР было проведено исследование на мышах нормального кариотипа и мышах, несущих в своем кариотипе робертсоновские транслокации (Крушинский и др., 1976). Оказалось, что у мышей разных линий доля особей, правильно решивших задачу, недостоверно отличается от 50 %-ного (случайного) уровня. Однако мыши, гомозиготные по транслокации Т1 ИЭМ (Rb/8,17/T1 ИЭМ), в достоверном большинстве случаев решают задачу на экстраполяцию правильно как при первом, так и при многократных предъявлениях.
Исследования, проводимые по селекции мышей на различный размер мозга, также показали, что генотипические факторы оказывают влияние на способность к экстраполяции у животных разных линий. Селекция на большой и малый вес мозга была начата на основе гетерогенной популяции, полученной путем скрещивания шести инбредных линий лабораторных мышей. Критерием для селекции служила регрессия, связывающая два признака — все тела и вес мозга. Уже с четвертого поколения установилось достоверное (р<0,001) различие среднего веса мозга у сублиний «большой» и «малый мозг». К 14-му поколению это различие достигло 63 мг. Доля правильных решений у сублинии «малый мозг» оказалась достоверно выше 50 %-ного случайного уровня, но начиная с 13-го поколения селекции правильные решения у обеих сублиний снизились до 50 %-ного случайного уровня. На основании проведенных исследований может быть сделан вывод, что различие в весе мозга не является ведущим фактором у особей, принадлежащих к одному виду, в решении элементарной логической задачи. Однако полученные данные указывают на изменение способности к экстраполяции у мышей селекционированных линий.
Проведенные опыты на крысах, мышах и сравнение способности к экстраполяции красных лисиц (Vulpes vulpes) и серебристо-черных (Vulpes fulvus) показали влияние генотипа на элементарную рассудочную деятельность у животных (Сотская, 1974). Сравнение уровня рассудочной деятельности диких и доместицированных форм показало, что доместицированные животные, как правил, обладают менее развитой рассудочной деятельностью, чем их дикие предки (Крушинский, 1977). Наиболее вероятной причиной такого различия является ослабление пресса естественного отбора у доместицированных животных. Ослабление действия естественного отбора приводит к увеличению изменчивости фенотипа и распаду полигенных систем, обеспечивающих наиболее высокие формы приспособления организмов к многообразным изменениям среды обитания. Такая дезинтеграция коадаптированных генетических систем является результатом дестабилизирующего отбора (Беляев, 1972).
При исследовании рассудочной деятельности у животных, которое проводится разными методами, мы столкнулись с неожиданным фактом появлением при решении предъявляемых тестов неадекватных форм поведения. Практически среди животных всех исследованных таксономических групп встречаются особи, у которых после нескольких правильных решений поведение во время эксперимента настолько меняется, что дает основание предполагать развитие какой-то патологии:
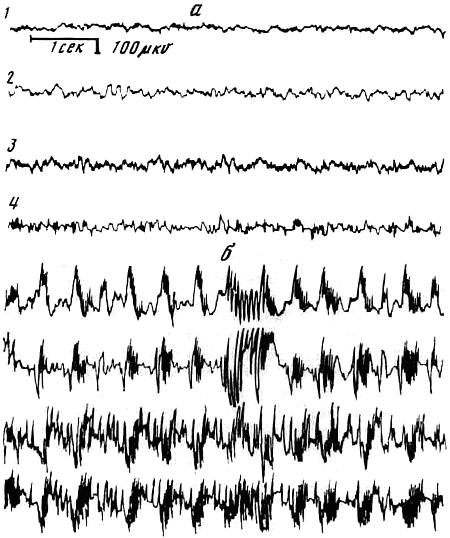
Рис. 4. Электроэнцефалограмма крысы после пяти правильных решений
а — фон; б — после решений задачи; 1 — сенсомоторная область коры; 2 — дорсальный гиппокамп, 3 — септум; 4 — латеральное поле гипоталамуса
Электрофизиологические исследования, проводимые на крысах и черепахах, показали, что при решении предъявляемых задач наблюдаются высоковольтные разряды и комплексы пик-волна, которые характерны для эпилептического припадка. Патологические разряды регистрировались одновременно как в корковых, так и подкорковых структурах мозга (Семиохина и др., 1976) (рис. 4).
Результаты этих исследований позволяют прийти к заключению, что при решении незнакомых для животных логических задач может развиваться резкое возбуждение мозга, которое широко иррадиирует по отдельным мозговым структурам. Оно приводит к появлению неадекватных реакций поведения: животное начинает бояться экспериментальной обстановки, убегает от нее и т. д. Очевидно, можно предположить, что не только у животных, но и у людей напряжение рассудочной деятельности, которое неизбежно в период творческой работы, может привести к резкому возбуждению мозга и появлению ряда патологических симптомов.
Таблица 2. Правильные решения у серых крыс, селекционированных на высокий уровень способности к экстраполяции, %

Удивительная, но вполне объяснимая связь между рассудочной деятельностью и патологией выступает при селекции. В лаборатории постнатального онтогенеза ИБР АН СССР у двух доместицированных линий пасюков (Rattus norvegicus) Н. Л. Крушинской и И. Л. Дмитриевой ведется селекция на способность крыс к экстраполяции. Отбор проведен в четырех поколениях (см. табл. 2).
Никакого эффекта улучшения рассудочной деятельности не обнаружено. Наоборот, было установлено, что от поколения к поколению уменьшается число хорошо решающих крыс и увеличивается число особей, с которыми трудно проводить опыты. Доля правильных решений резко уменьшается после первого предъявления задачи. Это обусловлено большой «нервозностью» крыс, которые плохо адаптируются к экспериментальной обстановке: стремятся убежать из нее или забиваются в угол камеры. Подобное же явление наблюдалось ранее и в нашей лаборатории.
Полученные данные о появлении невротиков в линиях крыс, селекционированных по признаку высокой рассудочной деятельности, едва ли случайны. Наиболее логичное объяснение этого факта сводится к следующему. При селекции на высокий уровень рассудочной деятельности отбираются в первую очередь наиболее возбудимые особи. Такие животные весьма предрасположены к всевозможным формам неврозов. Развитие неврозов мешает выявлению рассудочной деятельности, что и выражается в уменьшении процента в селекционируемых линиях крыс, хорошо решающих задачи. Сказанное предположение может быть проверено экспериментально. Если оно справедливо, то введение селекционированным крысам седативных средств должно уменьшить уровень возбудимости и ослабить или полностью купировать неадекватные формы их поведения.
Одним из ярких выражений неадекватности поведения явилась парадоксальная реакция на отодвигающийся за ширмой корм в опытах с экстраполяцией. В этом случае крысы шли не за кормом, а в противоположном направлении. Введение таким крысам мепробамата полностью купировало эту парадоксальную форму поведения. Ряд крыс начал адекватно решать предъявленную задачу.
Приведенные данные являются убедительной иллюстрацией значения уровня возбудимости как модификатора поведения и согласуются с высказанной рабочей гипотезой.
Таким образом, поднятая проблема о значении центральных механизмов нервной деятельности в фенотипическом выражении отдельных, частных, генотипически обусловленных особенностей переросла в более общую проблему — о регуляторном значении центральных механизмов нервной системы в фенотипической гетерогении поведения. В основе этой гетерогении лежит взаимодействие основных нервных процессов возбуждения и торможения, которое в каждый текущий момент определяет состояние возбудимости нервной системы.
Фенотипическая гетерогения поведения может быть разделена на две принципиально разные группы. К первой следует отнести гетерогению, являющуюся отражением взаимодействия основных нервных процессов: возбуждения и торможения. Основной характеристикой этого типа гетерогении являются парабиотические стадии, которые проявляются при разном уровне возбудимости нервной системы. В этом случае защитные механизмы, приводящие к стабилизации системы, являются источником гетерогении поведения. Ко второй группе мы относим гетерогению, обусловленную тем, что разный уровень возбудимости нервной системы оказывает модифицирующее влияние на проявление и выражение специфических особенностей поведения.
Проведенное исследование показало огромную роль центральных физиологических механизмов в фенотипической изменчивости как нормальных поведенческих актов, так и патологических реакций организма. Вместе с тем оно дало возможность убедиться в том, что в основе фенотипической корреляции между нормальными и патологическими признаками поведения лежит универсальный центральный механизм нервной системы, интегральное выражение которого определяется уровнем ее возбудимости.
Вероятно, одним из наиболее актуальных направлений регуляции фенотипической изменчивости поведения как в норме, так и при патологии является управление центральными физическими механизмами функций нервной системы; управление уровнем возбудимости целого мозга.
Нам кажется, что эта проблема имеет не только научно-познавательное (теоретическое) значение, но может представлять интерес и для медицины. В связи с этим нами начаты исследования на людях по изучению элементарных компонентов невербальной формы мышления (Крушинский, Попова, 1976). Работа проводится в настоящее время в двух направлениях: во-первых, изучается феноменологическая структура логической задачи, которая может быть предъявлена как животным, так и человеку; во-вторых, изучается роль генотипа в невербальной форме мышления человека. Для решения предъявляемой задачи человек или животное должны уловить закон перемещения предмета в пространстве. Задача состоит из двух основных компонентов: необходимо на основании проб и ошибок уловить закономерность перемещения объекта в пространстве при каждом последующем предъявлении задачи; определить шаг перемещения объекта. Первый этап — улавливание направления перемещения предмета осуществляется на основе способности к экстраполяции. Второй этап — улавливание шага перемещения объекта. Для этого индивидуум должен оперировать понятием размера («больше», «меньше», «равно»). Оказалось, что улавливание шага перемещения является более трудной задачей, чем определение направления перемещения объекта.
Задача была предложена 1073 испытуемым в возрасте от 2 до 25 лет. Показано, что успех решения задачи зависит от возраста. Некоторые дети уже в возрасте от 3 лет начинают решать задачу. Постепенно доля испытуемых, решающих задачи, увеличивается и достигает к 17–20 годам 80 % (у студентов). Следует отметить, что период школьного образования не оказывает существенного влияния на кривую успеха решения задачи от возраста.
Анализ проведен на 140 парах монозиготных и дизиготных близнецов и 70 парах неродственных особей, возраст которых соответствовал возрасту близнецов.[49] Было обнаружено большое сходство в решении задач у близнецов. Статистически достоверное сходство в решении (р<0,001) наблюдается лишь в монозиготных парах. Результаты анализа полученного материала дают основание допустить роль генотипических факторов в невербальном мышлении человека.
Необходимо подчеркнуть, что одной из важнейших задач физиологогенетического исследования является экспериментальное изучение роли центральных физиологических механизмов нервной деятельности в проявлении и выражении как нормального, так и патологического поведения.
Уровень возбудимости мозга является важнейшим фактором, обусловливающим проявление и выражение различных форм поведения в фенотипе.
Несомненно, что перевозбужденный мозг — главнейший источник весьма тяжелых патологий (как нервно-психических, так и сердечно-сосудистых). Поэтому управление уровнем — возбудимости мозга является важнейшей задачей, которая может быть решена только совместными усилиями специалистов в области физиологии, медицинской генетики, патофизиологии, биохимии и фармакологии.
Использование генетических моделей, полученных на животных, явится важнейшим этапом в разработке методов управления уровнем возбудимости мозга.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
7. РОЛЬ БЕССОЗНАТЕЛЬНОГО ПРИ НЕКОТОРЫХ ФОРМАХ ПАТОЛОГИИ
7. РОЛЬ БЕССОЗНАТЕЛЬНОГО ПРИ НЕКОТОРЫХ ФОРМАХ ПАТОЛОГИИ Формированием условного рефлекса на неосознаваемые внешние стимулы объясняется нервный механизм пространственной ориентации слепых людей [Бериташвили, 1969]. Существенная роль звуковых раздражений для ощущения
8. ПРОЕКЦИЯ ИНДИВИДУАЛЬНОГО ОПЫТА НА СТРУКТУРЫ МОЗГА В НОРМЕ И ПАТОЛОГИИ
8. ПРОЕКЦИЯ ИНДИВИДУАЛЬНОГО ОПЫТА НА СТРУКТУРЫ МОЗГА В НОРМЕ И ПАТОЛОГИИ 8.1. Зависимость проекции индивидуального опыта от особенностей индивидуального развития Из всего сказанного ранее с очевидностью следует, что с позиций системной психофизиологии проблема
8.4. Изменение проекции индивидуального опыта в условиях патологии
8.4. Изменение проекции индивидуального опыта в условиях патологии Проекция опыта на структуры мозга может изменяться не только при нормальном ходе индивидуального развития. Повреждающие воздействия, такие, например, как перерезка нервов или ампутация пальцев,
8.6. Значение материала патологии для изучения системной организации поведения
8.6. Значение материала патологии для изучения системной организации поведения Таким образом, проекция индивидуального опыта на структуры мозга изменяется в филогенезе, определяется историей обучения в процессе индивидуального развития и модифицируется при
Порочные круги во внутриклеточных системах как основа клеточной патологии
Порочные круги во внутриклеточных системах как основа клеточной патологии Вводные замечания можно распространить и на порочные потенциально патогенетические круги во внутриклеточных системах. Следует сказать, что в клетке труднее, чем в физиологических системах,
Метаболиты в норме и при патологии
Метаболиты в норме и при патологии В живой клетке ежесекундно образуются сотни метаболитов. Однако их концентрации поддерживаются на определенном уровне, который является специфической биохимической константой или референтной величиной. При болезнях происходит
Глава 21. Обмен сложных липидов
Глава 21. Обмен сложных липидов К сложным липидам относят такие соединения, которые, помимо липидного, содержат и нелипидный компонент (белок, углевод или фосфат). Соответственно существуют протеолипиды, гликолипиды и фосфолипиды. В отличие от простых липидов,
Патологии системы свертывания крови.
Патологии системы свертывания крови. ГемофилииГемофилин – наследственные заболевания, обусловленные отсутствием определенных факторов свертывания крови. Гемофилия А связана с дефицитом фактора VIII, гемофилия В (болезнь Кристмаса) – фактора IX, гемофилия С – фактора XI.
Патологии органов мужской половой системы
Патологии органов мужской половой системы Аномалии строения полового членаВ ряде случаев у кобелей встречаются аномалии в виде укорочения, искривления, сужения крайней плоти (фимоз), недоразвития дистального отдела мочеиспускательного канала, при котором его наружное
ОБЩЕЕ ВВЕДЕНИЕ ПОЧЕМУ ДЛЯ РЕШЕНИЯ СЛОЖНЫХ ВОПРОСОВ ДОСТАТОЧНО ПРОСТЫХ ИССЛЕДОВАНИЙ
ОБЩЕЕ ВВЕДЕНИЕ ПОЧЕМУ ДЛЯ РЕШЕНИЯ СЛОЖНЫХ ВОПРОСОВ ДОСТАТОЧНО ПРОСТЫХ ИССЛЕДОВАНИЙ В этой книге я предлагаю провести семь экспериментов, которые могли бы изменить наше представление об окружающем мире. Эти эксперименты способны вывести нас за пределы современного
2.1. Вопрос нормы и патологии. Определение нормальной психики
2.1. Вопрос нормы и патологии. Определение нормальной психики В работе с животными, проявляющими аномальное поведение, одним из базовых понятий, на которое должны опираться в своей деятельности специалисты в области поведенческой медицины, это различение нормы и
Зачатки высших форм поведения
Зачатки высших форм поведения Многощетинковые кольчатые черви относятся к наиболее развитым в психическом отношении низшим беспозвоночным. Их поведение отличается подчас большой сложностью и представляет особый интерес в том смысле, что содержит ряд элементов
Патологии беременности и послеродового периода
Патологии беременности и послеродового периода Выпадение влагалища наблюдается во второй половине беременности при погрешностях кормления и отсутствии моциона. Оно обусловливается повышением внутрибрюшного давления в сочетании с расслаблением сфинктера преддверия
Глава 5. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ КАК ОСНОВА ОРГАНИЗАЦИИ СЛОЖНЫХ ФУНКЦИЙ
Глава 5. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ КАК ОСНОВА ОРГАНИЗАЦИИ СЛОЖНЫХ ФУНКЦИЙ Анализ процессов пищеварения и транспорта на современном уровне и в особенности эволюции этих процессов открывает новые горизонты для понимания принципов организации сложных
Эволюционно-генетические проблемы поведения животных и человека[51]
Эволюционно-генетические проблемы поведения животных и человека[51] Известно, что одним из открытий И. П. Павлова явилось обнаружение патологии поведения животных (а также человека) при чрезмерном напряжении высшей нервной деятельности. Большой вклад в исследование
Генетические методы и проблемы разведения домашних животных
Генетические методы и проблемы разведения домашних животных Существует несколько классификаций методов разведения домашних животных. У американского ученого В. Райса пять: поглощение крови, прилитие крови, скрещивание, разведение по линиям и инбридинг. Академик Е. Ф.