1.3. Общность фундаментальных процессов ассимиляции пищи
1.3. Общность фундаментальных процессов ассимиляции пищи
Итак, формирование трофологии стало возможным благодаря достижениям ряда современных наук, после того как было продемонстрировано, что все разнообразие известных в биологии трофических процессов на микро-, макро- и мегауровнях сводится к некоторым общим закономерностям, которые будут рассмотрены ниже. Забегая вперед, следует сказать, что процессы питания базируются на двух фундаментальных принципах — принципе универсальности строительных блоков и принципе универсальности функциональных блоков (см. 1.4). Лишь благодаря этому возможно построение трофических цепей. Таким образом, имеет место сложнейшая интеграция отдельных явлений и свойств природы в грандиозный комплекс, именуемый биосферой в целом, с его большим биотическим круговоротом.
Как отмечено выше, успехи молекулярной биологии, мембранологии, цитологии, физиологии и биохимии позволили установить общие закономерности строения и функционирования систем, обеспечивающих ассимиляцию пищи всеми без исключения группами организмов: бактериями, грибами, простейшими, растениями и животными (обзоры: Уголев, 1983, 1985, 1987а, 1989, 1990). Это относится как к экзотрофии — утилизации пищевых веществ, поступающих из окружающей среды, так и к эндотрофии — утилизации веществ, содержащихся в депо или структурах различных клеток, т. е. к использованию внутренних ресурсов организма.
Экзо- и эндотрофия включают два фундаментальных этапа. Первый из них — собственно пищеварение, или деполимеризация (трансформация) пищевых веществ, в результате чего разрушаются крупные молекулы и надмолекулярные комплексы пищи и образуются транспортируемые и метаболизируемые формы. Второй — транспорт этих веществ, а также других, которые поступают во внутреннюю среду организма без предварительной обработки в пищеварительной системе или в отдельных клетках. Все разнообразие процессов деполимеризации пищи у всех живых организмов — от бактерий до млекопитающих, как стало известно в конце 50-х годов, сводится к трем основным типам пищеварения: внеклеточному, внутриклеточному и мембранному (рис. 1.1).
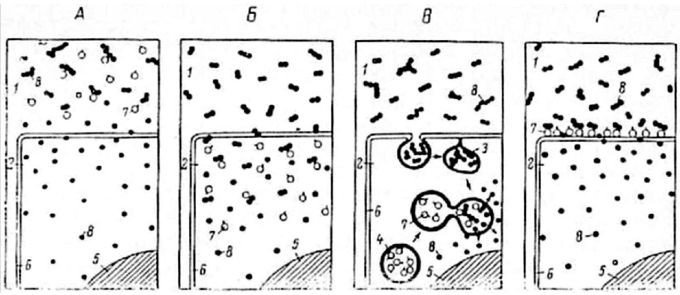
Рис. 1.1. Основные типы пищеварения (по: Уголев, 1985).
А — внеклеточное дистантное пищеварение;
Б — внутриклеточное цитоплазматическое пищеварение;
В — внутриклеточное везикулярное, или внеплазматическое, пищеварение, связанное с эндоцитозами (пино- и фагоцитозом);
1- внеклеточная среда; 2 — внутриклеточная среда; 3 — внутриклеточная пищеварительная вакуоль; 4 — лизосома; 5 — ядро; 6 — мембрана; 7 — ферменты; 8 — субстраты и продукты из гидролиза.
Важно, что эти три типа пищеварения являются основными как при экзо-, так и при эндотрофии. Детальная характеристика мембранного и других основных типов пищеварения представлена во многих сводках, в том числе и наших (Уголев, 1963, 1967, 1972, 1985; Мембранный гидролиз…, 1986; Membrane digestion…, 1989).
1.3.1. Внеклеточное пищеварение
Этот тип пищеварения характеризуется тем, что синтезированные в клетке ферменты выделяются за ее пределы во внеклеточную среду, где реализуется их гидролитический эффект. При внеклеточном пищеварении ферменты растворены в водной фазе и их распределение определяется законами теплового движения. В связи с этим возможна любая ориентация активных центров ферментов по отношению к субстратам, а структурная организация ферментных систем ограничена или невозможна. Растворенные в водной фазе ферменты атакуют поглощенные организмом субстраты, разрушая, в частности, крупные молекулы и надмолекулярные агрегаты, и обеспечивают главным образом начальные этапы пищеварения. Внеклеточное пищеварение обнаружено у всех организмов, включая бактерий. У человека и высших животных этот тип пищеварения называется полостным, так как реализуется в специальных полостях — ротовой полости, полостях желудка и тонкой кишки. В тонкой кишке высших организмов полостное пищеварение сочетается с мембранным и иногда внутриклеточным.
1.3.2. Внутриклеточное пищеварение
Под внутриклеточным пищеварением понимаются все случаи, когда нерасщепленный или частично расщепленный субстрат проникает внутрь клетки, где подвергается гидролизу не выделяемыми за ее пределы ферментами. Внутриклеточное пищеварение может быть разделено на два подтипа — молекулярный и везикулярный. Молекулярное внутриклеточное пищеварение характеризуется тем, что ферменты, находящиеся в цитоплазме, гидролизуют проникающие в клетку небольшие молекулы субстрата, главным образом димеры и олигомеры, причем проникают такие молекулы пассивно или активно. Например, с помощью специальных транспортных систем, активно, переносятся через клеточную мембрану дисахариды и дипептиды у бактерий. Допускается, что и у высших организмов, в частности у млекопитающих, некоторые дипептиды могут активно транспортироваться внутрь кишечных клеток — энтероцитов. Если внутриклеточное пищеварение происходит в специальных вакуолях, или везикулах, которые образуются в результате эндоцитоза (пиноцитоза или фагоцитоза), то оно определяется как везикулярное, или эндоцитозное. При везикулярном внутриклеточном пищеварении эндоцитозного типа происходит впячивание определенного участка (участков) мембраны вместе с поглощаемым веществом. Далее этот участок постепенно отделяется от мембраны, и образуется внутриклеточная везикулярная структура. Как правило, такая везикула сливается с лизосомой, содержащей широкий спектр гидролитических ферментов, действующих на все основные компоненты пищи. В образовавшейся новой структуре — фагосоме и происходят гидролиз поступивших субстратов и последующее всасывание образующихся продуктов. Непереваренные остатки фагосомы обычно выбрасываются за пределы клетки путем экзоцитоза. Таким образом, внутриклеточное пищеварение — это механизм, за счет которого реализуется не только переваривание, но и поглощение клеткой пищевых веществ, в том числе крупных молекул и надмолекулярных структур. Внутриклеточное пищеварение лимитировано проницаемостью мембраны и процессами эндоцитоза. Последние характеризуются низкой скоростью и, по-видимому, не могут играть существенной роли в обеспечении нутритивных потребностей высших организмов. Как мы обратили внимание еще в 1967 г. (Уголев, 1967), с точки зрения энзимологии внутриклеточное пищеварение везикулярного типа представляет собой сочетание микрополостного и мембранного пищеварения. Везикулярное внутриклеточное пищеварение выявлено у всех типов животных — от простейших до млекопитающих (особенно большую роль оно играет у низших животных), а молекулярное — у всех групп организмов.
1.3.3. Мембранное пищеварение
Мембранное пищеварение происходит на границе внеклеточной и внутриклеточной сред и обладает некоторыми особенностями как внеклеточного, так и внутриклеточного пищеварения. Этот тип пищеварения обнаружен у всех организмов. У человека и высших животных мембранное пищеварение реализуется преимущественно в тонкой кишке ферментами, связанными со структурами мембраны кишечных клеток. К этим ферментам относятся: 1) ферменты, секретируемые клетками поджелудочной железы и адсорбированные на поверхности кишечных клеток, главным образом в гликокаликсе; 2) собственно кишечные ферменты, которые синтезируются в самих кишечных клетках и затем встраиваются в их апикальную мембрану. Активные центры ферментов, осуществляющих мембранное пищеварение, обращены в полость тонкой кишки, т. е. ориентированы определенным образом по отношению к мембране и водной фазе. Этим мембранное пищеварение существенно отличается от полостного и внутриклеточного типов переваривания. Мембранное пищеварение малоэффективно по отношению к крупным молекулам и тем более надмолекулярным агрегатам. Панкреатические ферменты, адсорбированные на структурах кишечной слизистой оболочки, реализуют преимущественно промежуточные стадии гидролиза пищевых веществ (углеводов, белков, жиров, витаминов и т. д.), мембранные — преимущественно заключительные стадии их расщепления. Мембранное пищеварение также интегрировано с процессами полостного переваривания и всасывания. Более того, собственно кишечные ферменты и транспортные системы мембраны могут образовывать ферментно-транспортные комплексы, благодаря которым продукты гидролиза получают преимущества при всасывании (рис. 1.2).
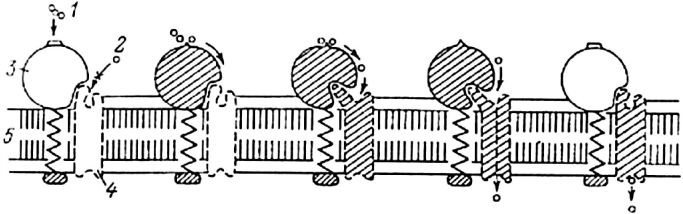
Рис. 1.2. Схема последовательных конформационных взаимодействий ферментной и транспортной частей функционирующего ферментно-транспортного комплекса (по: Уголев, 1985).
1 — субстрат; 2- продукт; 3 — трансмембранный фермент; 4 — транспортная система; 5 — мамбрана.
1.3.4. Схема реального усвоения пищевых веществ
В конце 50-х годов после открытия мембранного пищеварения классическая двухзвенная схема ассимиляции пищевых веществ (внеклеточное пищеварение-всасывание) была заменена трехзвенной (внеклеточное пищеварение-мембранное пищеварение-всасывание). Внутриклеточное везикулярное пищеварение, присущее преимущественно низшим организмам, у человека и высших животных играет важную роль, в частности на самых ранних этапах постнатального развития млекопитающих (Уголев и др., 1979; обзор: Henning, 1987). Внутриклеточное молекулярное пищеварение у человека и высших животных служит, по-видимому, дополнительным механизмом гидролиза ди- и трипептидов (обзоры: Gardner, 1984, 1988).
Для высших организмов наиболее характерна комбинация полостного и мембранного пищеварения, между которыми существует сложная функциональная зависимость, так как конечные продукты первого служат субстратом для второго (рис. 1.3). Крупные молекулы субстратов остаются в полости тонкой кишки и там гидролизуются главным образом ферментами, секретируемыми клетками поджелудочной железы. Относительная роль полостного пищеварения тем больше, чем крупнее молекулы пищевых веществ, а относительная роль мембранного гидролиза увеличивается по мере уменьшения размеров молекул. При определенных условиях мембранное пищеварение становится доминирующим механизмом, что, в частности, наблюдается в тонкой кишке млекопитающих в период их молочного питания, а также у некоторых кишечных паразитов.
Благодаря анализу особенностей питания разнообразных организмов стало ясно, что все три перечисленные типа пищеварения существуют на всех уровнях эволюционной лестницы — от бактерий до млекопитающих. Однако в зависимости от ряда причин, в том числе от типов питания, может доминировать один из них. Все многообразие пищеварительных механизмов обусловлено сочетанием трех основных типов пищеварения. Предполагается, что они возникли на ранних стадиях формирования жизни на основе механизмов, обеспечивающих эндогенное питание. Из двух разновидностей внутриклеточного пищеварения (молекулярного и везикулярного) последний появился лишь у эукариотов и представляет собой, как упомянуто выше, комбинацию микрополостного и мембранного пищеварения.
Следует заметить, что у некоторых насекомых первая стадия пищеварения протекает не только вне пищеварительной полости, но даже за пределами организма. Однако это только кажущееся противоречие. Хотя такие насекомые начинают переваривать свою жертву еще до поглощения, вводя свои ферменты в ее тело, ясно, что это — определенная модификация внеклеточного полостного пищеварения. Существуют и еще кажущиеся исключения. Так, многие кишечные паразиты потеряли способность секретировать пищеварительные ферменты. Однако внеклеточное пищеварение у них происходит за счет ферментов хозяина. Более того, у ряда паразитов часто адсорбируются ферменты хозяина для реализации промежуточных этапов мембранного пищеварения (обзоры: Уголев, 1985; Уголев и др., 1985).
В большинстве случаев усвоение пищевых веществ требует участия симбионтов — бактерий и простейших (см. 1.3.5). Таким путем ряду организмов удалось приспособиться к потреблению целлюлозы, шерсти, хитина и других материалов, не гидролизуемых собственными ферментами. Однако и здесь взаимодействуют известные типы пищеварения у макроорганизма и его симбионтов.
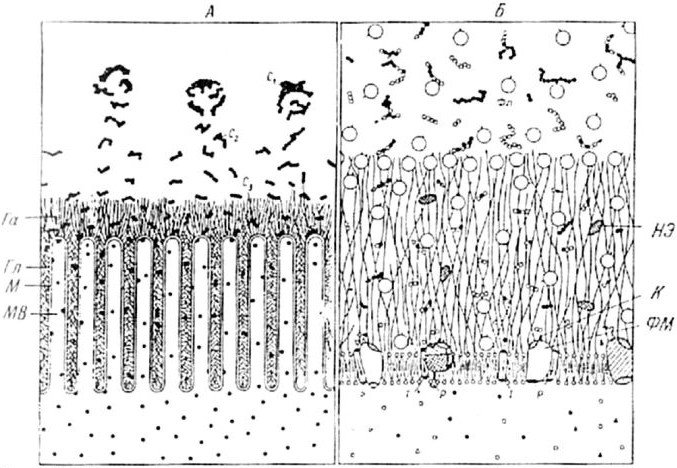
Рис. 1.3. Схема взаимодействий между полостным и мембранным пищеварение (по: Уголев и др., 1983).
А — последовательная деполимеризация пищевых субстратов в полости и на поверхности тонкой кишки; Б — фрагмент липопротеиновой мембраны с адсорбированными и собственно кишечными трансмембранными фарментами. М — мембрана; МВ — микроворсинки; Га — апикальный глококаликс; Гл — латеральный гликокаликс; С1-С3 — субстраты; Фп — панкреатические ферменты; Фм — мембранные ферменты; Т — транспортная система мембраны; Р — регуляторные центры ферментов; К — каталитические центры ферментов; НЭ — неэнезиматические факторы.
1.3.5. Симбионтное пищеварение и питание
Особое место среди других механизмов переваривания пищи занимает симбионтное пищеварение, т. е. пищеварение за счет микроорганизмов желудочно-кишечного тракта. Оно присуще почти всем многоклеточным организмам и отчетливо проявляется у растительноядных жвачных. Симбионтное пищеварение характерно и для других животных, подобных истинным жвачным, например для верблюда, ламы и др. Этот тип пищеварения широко распространен также у беспозвоночных, например у насекомых. Симбионтное пищеварение подробно охарактеризовано в ряде капитальных обзоров (Prosser, Brown, 1967; Сравнительная физиология…, 1977; Schmidt-Nielsen, 1982; Martin, 1989; Williams, 1989, и др.).
Как правило, пищеварительный тракт макроорганизма заселен бактериями и простейшими, которые частично или полностью снабжают организм хозяина необходимыми органическими веществами, в том числе витаминами, незаменимыми аминокислотами и др. Симбионтами некоторых животных, например моллюсков, актиний, кораллов, могут быть также водоросли — зооксантеллы и зоохлореллы, поставляющие пищевые вещества хозяину. Интересно, что у хищных насекомых симбионты отсутствуют, тогда как у тех, кто потребляет сок растений, симбионтное пищеварение играет весьма важную роль.
Анализ различных форм и модификаций симбионтного пищеварения позволил выделить в нем два фундаментальных биологических механизма: 1) первичные пищевые вещества разрушаются ферментами бактерий и простейших, а образующиеся продукты гидролиза используются преимущественно организмом хозяина; 2) бактерии и простейшие не только разрушают первичные пищевые продукты, но и утилизируют их, тогда как хозяин поглощает вторичную пищу, состоящую из структур симбионтов. Первый механизм точнее было бы назвать симбионтным пищеварением, а второй — симбионтным питанием в сочетании с симбионтным пищеварением. Животные со специализированными симбионтами, способными использовать метан и серу, оказались в начале трофической цепи в биологических изолятах на дне океанов, где наблюдалась высокая вулканическая активность. В этом случае существуют выходы метана или сероводорода, которые служат внешними источниками энергии подобно свету.
Симбионтные взаимодействия широки и разнообразны. Например, приспособление жвачных к использованию грубых растительных кормов во всех случаях связано с действием на эти субстраты симбионтов. Можно не считать преувеличением, что существование таких организмов в основном базируется на использовании симбионтных процессов.
При рассмотрении симбионтных питания и пищеварения взаимодействия бактериальной флоры и организма-хозяина анализируются обычно на примере человека и высших животных, преимущественно жвачных. Однако для понимания механизмов питания и пищеварения в естественных условиях, а также для формирования новых оптимальных искусственных условий целесообразно анализировать весь круг явлений, известных среди организмов. Нутритивные симбиозы известны уже у простейших, в которые вселяются бактерии и некоторые водоросли, обеспечивающие их продуктами фотосинтеза. Среди многоклеточных организмов уже у кишечнополостных обнаруживаются зоохлореллы и зооксантеллы, синтезирующие углеводы и снабжающие организм хозяина дополнительным кислородом.
Организмы, содержащие симбионты, более устойчивы к пищевому голоданию и недостатку кислорода. Можно думать, что во многих случаях быстрота адаптации и, следовательно, возможность изменения трофической ниши связаны не только, а иногда и не столько с изменением пищеварительных характеристик самого хозяина, сколько с изменением ферментных характеристик микрофлоры, населяющей желудочно-кишечный тракт (см. гл. 3). Много ярких примеров этому мы видим у насекомых. Так, питание шерстью или шелком зависит от формирования специфической кишечной флоры и по своему механизму является адаптацией на уровне бактериальной флоры. Следовательно, необходима разгадка того, каким образом организм хозяина культивирует нужный тип микрофлоры и затем передает потомству иногда чистую или почти чистую и хорошо стабилизированную культуру бактерий. Более того, образование экзотических трофических ниш и заполнение трофических пустот также в ряде случаев связаны с приобретением и культивированием соответствующих симбионтов.
Нельзя исключить, что в составе нормальной флоры желудочно-кишечного тракта всегда присутствуют «случайные» загрязняющие бактерии, способные утилизировать и расщеплять нехарактерные для данной популяции пищевые субстраты. Таким путем, вероятно, происходят эволюция питания и приспособление организмов к новым трофическим нишам. Кроме того, у многоклеточных организмов в пределах пищеварительного аппарата могут существовать своеобразные трофические цепи, что обеспечивает многие важнейшие эволюционные и экологические перестройки. Например, термиты, питающиеся преимущественно древесиной, обеспечивают свои потребности в белке за счет включения в кишечную флору азотфиксирующих бактерий. По-видимому, благодаря присутствию в желудочно-кишечном тракте азотфиксирующих бактерий действительно возможны подлинная фиксация атмосферного азота и включение его в биологический цикл, минуя промежуточные звенья растительной трофической цепи. Этот пример является многообещающим и демонстрирует недостаточно изученную, но исключительно важную сторону эволюции процессов питания и пищеварения в животном мире.
1.3.6. Индуцированный аутолиз
Ранее нами было высказано предположение о важной роли так называемого индуцированного аутолиза в ассимиляции пищи. Согласно нашей гипотезе, этот тип пищеварения, при котором объект питания в естественных условиях обеспечивает собственное переваривание и усвоение, является наиболее древним механизмом начальной переработки пищевых веществ у многоклеточных организмов. Индуцированный аутолиз реализуется собственными ферментами пищевого объекта, а пищеварительные ферменты хозяина создают благоприятные условия для их действия. Однако до последнего времени считалось, что аутолиз играет незначительную роль в общем пищеварительном процессе, так как начальные этапы пищеварения осуществляются в основном в полости желудочно-кишечного тракта высших организмов секретируемыми ферментами хозяина, а промежуточные и заключительные — в результате мембранного пищеварения.
Таким образом, было важно установить относительную роль ферментов хозяина и пищевого объекта в начальных этапах гидролиза пищевых веществ при питании в естественных условиях. Особенно трудно было интерпретировать питание хищников, у которых происходит переваривание больших порций мяса, из-за чего отношение поверхность/масса мало и невыгодно для ферментов желудочного сока, действующих на поверхности пищевого объекта.
Индуцированный аутолиз был обнаружен нами около 20 лет назад, но описан лишь в 1984 г. (Уголев, Цветкова, 1984). (Детально этот механизм освещен также в обзорах: Уголев, 1985, 1987а.) При индуцированном аутолизе жертва, или, шире говоря, объект питания, обеспечивает свое собственное переваривание. Так, например, происходит, если удав заглатывает кролика. До последнего времени было неясно, каким образом проглоченная целиком жертва переваривается удавом. Действительно, поверхность контакта жертвы с ферментами желудочного сока хищника сравнительно мала, так как пищевой объект не раздроблен. Оказалось, однако, что раньше, чем ферменты хищника гидролизуют структуры жертвы, начиная с ее поверхности, организм этой жертвы будет аутолизироваться за счет индуцированного аутолиза.
Индуцированный аутолиз был исследован нами в модельных экспериментах, названных «маленький искусственный удав». В прозрачную камеру, заполненную желудочным соком человека, лошади или собаки, помещались «сырая» лягушка и лягушка после недолгой термической обработки. В первые несколько часов гидролиз сухожилий термически обработанной лягушки шел быстрее, чем «сырой», что служило подтверждением общепринятых взглядов. Однако в последующие 2–3 дня «сырая» лягушка полностью растворялась, тогда как структуры термически обработанной лягушки в значительной мере сохранялись. Таким образом, в этих экспериментах наряду с доказательством существования индуцированного аутолиза было продемонстрировано, что нативные белки гидролизуются быстрее денатурированных.
В дальнейшем мы детально исследовали механизм переваривания нативных тканей желудочным соком.
Выяснилось, что суть этого механизма состоит в следующем. Кислый желудочный сок хищника индуцирует самопереваривание жертвы ее же ферментами. Под действием этого сока происходят разрушение лизосом и выход в клетку многочисленных лизосомальных гидролаз, разрушающих все структуры клетки при pH 3.5–5.5.
Итак, организм-ассимилятор индуцирует расщепление структур пищевого объекта ферментами последнего, активируя их и создавая оптимальные условия среды, в том числе pH. В соответствии с классическими представлениями о механизмах переваривания пищевого объекта, ферменты пищеварительных соков осуществляют свой эффект только поверхностно. Скорость диффузии гидролаз внутрь пищевого объекта даже при низком диффузионном сопротивлении мембраны лимитирована их сравнительно большой молекулярной массой, тогда как скорость диффузии протона примерно на 3–4 порядка больше. При выходе гидролаз из лизосом под влиянием сдвига pH возникает множество центров гидролиза в каждой клетке (рис. 1.4), что создает практически тотальное расщепление ткани. Следует добавить, что в кислых секретах организма-ассимилятора содержатся главным образом протеазы, тогда как ферментный спектр лизосом практически универсален. Однако в ассимилируемых объектах имеются также структуры (например, белки соединительной ткани, жировые депо, в тканях растений — полисахаридные депо), лишенные лизосом и не подвергающиеся индуцированному аутолизу. Следовательно, можно предположить, что ферменты пищеварительных соков (гидролазы, расщепляющие белки, жиры и углеводы) особенно важны для утилизации указанных структур с высокой скоростью.
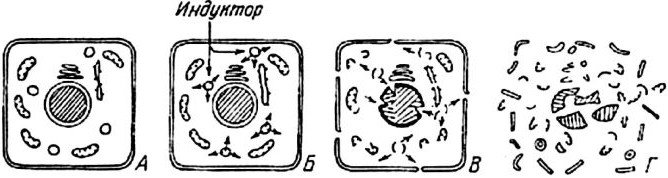
Рис. 1.4. Упрощенная концептуальная схема внутриклеточный процессов при индуцированном аутолизе (по: Уголев, 1985).
А — клетка с субклеточными органеллами (светлые кружки — лизосомы); Б — проникновение индукторов (Н+, детергентов и т. д.) в клетку; В, Г — выход лизосомальных ферментов в цитоплазму при действии индукторов и лизис клеточных структур (при подходящих условиях).
По-видимому, индуцированный аутолиз существует не только у плотоядных, но и у растительноядных животных. Например, при использовании травоядными свежих кормов расщепление пищи отчасти обусловлено ферментами, находящимися в клетках растений. Некоторое значение в гидролизе пищевых веществ в желудочно-кишечном тракте новорожденных детей могут иметь гидролитические ферменты материнского молока. Наконец, «созревание» мяса, по-видимому, есть не что иное, как частичный аутолиз, который можно рассматривать как некоторый элемент пищеварительного процесса.
Примеры индуцированного аутолиза обнаружены также среди низших животных, в частности у простейших и плоских червей. После захвата пищевого объекта происходит образование новых вакуолей, или везикул, обладающих кислой реакцией. Хотя кислая фаза переваривания в этих случаях была интерпретирована как выполняющая лишь денатурацию структур пищевого материала, представленные данные свидетельствуют о возможности значительного расщепления пищи в результате индуцированного аутолиза. В соответствии с этой гипотезой кислая среда пищеварительных везикул приводит к активации лизосомальных ферментов двумя путями: 1) увеличивая проницаемость мембран лизосом; 2) создавая адекватные условия для действия лизосомальных ферментов, которые активны в слабокислой среде. Следовательно, возможен не только протеолиз, но и расщепление множества других пищевых веществ.
Можно думать, что представления об индуцированном аутолизе позволяют глубже понять механизмы естественного переваривания пищи, ее легкое усвоение без термической обработки и, наконец, то обстоятельство, что индуцированный аутолиз — это важное и древнее приспособление последовательных партнеров в трофической цепи.
1.3.7. Транспорт
Основные концепции транспорта пищевых веществ через клеточные мембраны, клеточные слои и межклеточные пути охарактеризованы в ряде фундаментальных обзоров (Никольский, 1977; Елецкий, Цибулевский, 1979; Levin, 1979; Crane et al., 1980; Hoshi, Himukai, 1982; Pharmacology…, 1984; Уголев, 1985, 1987a; Мембранный гидролиз…, 1986; Semenza, Corcelli, 1986; Alpers, 1987; Hopfer, 1987; Shiau, 1987; Molecular basis…, 1988; Evans, Graham, 1989, и др.).
Долгое время доминировало представление об исключительном разнообразии механизмов транспорта у различных организмов. Тем более удивительно, что, как оказалось, у организмов, стоящих на разных уровнях эволюционной лестницы, механизмы трансмембранного и трансцеллюлярного транспорта пищевых веществ идентичны или сходны. Это сходство выражается прежде всего в общих принципах построения и функционирования транспортных систем клеточной мембраны. Среди них общий источник энергии для активного транспорта — АТФ; наличие высокоспецифических связывающих и транспортирующих белков; сопряженность избирательного трансмембранного переноса глюкозы и аминокислот с переносом ионов натрия или протонов; идентичность механизмов действия некоторых активаторов и ингибиторов на транспортные системы далеких друг от друга видов организмов, и т. д. Важно, что у всех групп многоклеточных организмов, и даже у Protozoa и, наконец, в ряде случаев у прокариотов, обнаружена идентичность систем переноса пищевых веществ через мембраны клеток или через клеточные слои (рис. 1.5).
Согласно современным представлениям, существует два типа транспорта — макромолекулярный и микромолекулярный. Под первым подразумевается перенос крупных молекул и надмолекулярных комплексов. Этот тип транспорта в большинстве случаев осуществляется с помощью эндоцитоза и обеспечивает, например, проникновение небольших, но значимых для организма количеств белка через кишечный барьер млекопитающих. Однако основным типом транспорта пищевых веществ у всех организмов, по-видимому, является микромолекулярный, в результате которого через клеточную мембрану переносятся преимущественно мономеры (различные ионы, глюкоза, аминокислоты, жирные кислоты и другие молекулы), а также олигомеры с небольшой молекулярной массой. Например, у высших организмов углеводы транспортируются через плазматическую мембрану кишечных клеток в основном в виде моносахаридов (глюкозы, галактозы, фруктозы и т. д.), белки — в виде аминокислот, жиры — в виде жирных кислот и глицерина.
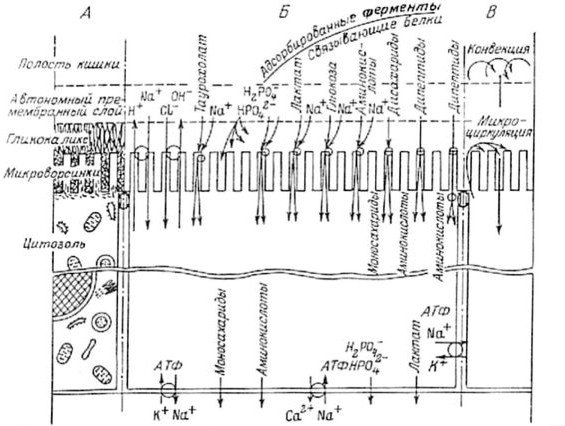
Рис. 1.5. Схема структурных и функциональных компартментов кишечной слизистой (по: Смирнов, Уголев, 1981).
А — Структура компартментов; Б — потоки веществ; В — потоки воды.
Через мембрану клеток вещества проникают за счет пассивного транспорта, облегченной диффузии и активного транспорта. Пассивный транспорт объединяет в себе процессы диффузии и осмоса. Облегченная диффузия осуществляется особыми переносчиками, или транспортерами, — специфическими белковыми молекулами, которые облегчают проникновение субстратов через липопротеиновую мембрану клетки без затраты энергии. Активный транспорт — это процесс проникновения веществ через плазматическую мембрану клетки против электрохимического градиента. В этом процессе, требующем затраты энергии, участвуют специальные транспортные системы, которые функционируют по типу мобильных переносчиков, конформационных переносчиков или, что встречается наиболее часто, каналов (обзор: Hopfer, 1987, и др.).
Имеются серьезные основания полагать, что существует особый транспортный механизм, названный нами «подвижная адсорбция», который был рассмотрен в нескольких наших работах, начиная с 1967 г. (Уголев, 1967). Такая адсорбция осуществляется благодаря движению молекул по активным поверхностям и центрам по градиенту концентраций, который может создаваться различными путями: за счет транспортных систем, локализованных в определенных точках и активно переносящих вещество из одного компартмента в другой, или с помощью ферментных систем, трансформирующих это вещество. Мы полагаем, что такой механизм можно было бы назвать транссорбцией.
Все перечисленные механизмы справедливы для организмов, стоящих на разных уровнях развития: от примитивных прокариотов до высших эукариотов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
10.1. Характеристика обменных процессов
10.1. Характеристика обменных процессов Обмен веществ и энергии – основа процессов жизнедеятельности организма. В организме человека, в его органах, тканях, клетках идет непрерывный процесс синтеза, т. е. образования сложных веществ из более простых. Одновременно с этим
8.7. Регуляция процессов размножения и полового поведения
8.7. Регуляция процессов размножения и полового поведения Большой сложностью отличается нейрогуморальная регуляция процессов размножения и полового поведения. Роль гипоталамо-гипофизарной системы в этой регуляции особенно наглядна. Гипоталамус, посредством секреции
11.6. В дебрях когнитивных процессов
11.6. В дебрях когнитивных процессов Таким образом, мы можем видеть, что на протяжении всей истории психологии ее раздробленность не позволяла создать единый научный язык, без которого нельзя говорить о целостной науке. Внутри каждого направления и у каждого автора
Количество пищи
Количество пищи Прежде всего следует решить вопрос о необходимом количестве пищи. Нормальный человек руководствуется чувством голода. Как правило, количество пищи зависит от вида и объема выполняемой работы, хотя у каждого из нас существуют свои особенности, связанные
Глава 4 ПСИХОФИЗИОЛОГИЯ СЕНСОРНЫХ ПРОЦЕССОВ
Глава 4 ПСИХОФИЗИОЛОГИЯ СЕНСОРНЫХ ПРОЦЕССОВ 1. ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ Сенсорной системой называют часть нервной системы, воспринимающую внешнюю для мозга информацию, передающую её в мозг и анализирующую её. Сенсорная система состоит из воспринимающих
6. ДИСКРЕТНОСТЬ МНЕМИЧЕСКИХ ПРОЦЕССОВ
6. ДИСКРЕТНОСТЬ МНЕМИЧЕСКИХ ПРОЦЕССОВ Буквы алфавита, атомы и молекулы – всё это кодовые обозначения важных сущностей, значение открытий которых нельзя переоценить.Первым было открытие иероглифов и азбуки. Азбука состоит из малого количества букв. Например, кириллица
6.2. Научение как реактивация процессов развития
6.2. Научение как реактивация процессов развития В настоящее время становится общепризнанным, что многие закономерности модификации функциональных и морфологических свойств нейронов, а также регуляции экспрессии генов, лежащие в основе научения у взрослых, сходны с
Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Вся сложная и разнообразная деятельность высших отделов нервной системы построена на работе двух основных нервных процессов — возбуждения и торможения. Протекая в подвижных пространственных и временных
1.4. НЕЙРОЭНДОКРИННАЯ РЕГУЛЯЦИЯ ПОЛОВЫХ ПРОЦЕССОВ
1.4. НЕЙРОЭНДОКРИННАЯ РЕГУЛЯЦИЯ ПОЛОВЫХ ПРОЦЕССОВ Важнейшая функция нервной системы — управление деятельностью целостного организма при помощи нервных и гуморальных сигналов на основании сбора, анализа и интеграции информации, исходящей от отдельных частей организма
ГЛАВА 6 НЕПОСТОЯНСТВО «ФУНДАМЕНТАЛЬНЫХ КОНСТАНТ»
ГЛАВА 6 НЕПОСТОЯНСТВО «ФУНДАМЕНТАЛЬНЫХ КОНСТАНТ» ФУНДАМЕНТАЛЬНЫЕ ФИЗИЧЕСКИЕ КОНСТАНТЫ И ИЗМЕРЕНИЯ ИХ ЗНАЧЕНИЙ «Физические константы» представляют собой числа, которые ученые используют в своих вычислениях. В отличие от математических констант вроде числа ?, значения
ТЕОРИИ ИЗМЕНЯЮЩИХСЯ «ФУНДАМЕНТАЛЬНЫХ КОНСТАНТ»
ТЕОРИИ ИЗМЕНЯЮЩИХСЯ «ФУНДАМЕНТАЛЬНЫХ КОНСТАНТ» Несколько физиков — к примеру, Артур Эддингтон и Поль Дирак — после долгих размышлений пришли к выводу, что по крайней мере некоторые из «фундаментальных констант» могут со временем изменять свои значения. В частности,
ГЛАВА 6 НЕПОСТОЯНСТВО «ФУНДАМЕНТАЛЬНЫХ КОНСТАНТ»
ГЛАВА 6 НЕПОСТОЯНСТВО «ФУНДАМЕНТАЛЬНЫХ КОНСТАНТ» За последние несколько лет появилось множество новых данных по определению числового значения гравитационной постоянной G.[334]Как уже сообщалось в шестой главе, в период с 1970 по 1989 гг. «лучшие» значения этой константы
4. Иррадиация и концентрация процессов возбуждения и торможения
4. Иррадиация и концентрация процессов возбуждения и торможения Как было указано выше, возбуждение и торможение являются теми процессами, которые лежат в основе высшей нервной деятельности, в основе поведения. Взаимодействие этих процессов, их движение по коре
1.3. Общность фундаментальных процессов ассимиляции пищи
1.3. Общность фундаментальных процессов ассимиляции пищи Итак, формирование трофологии стало возможным благодаря достижениям ряда современных наук, после того как было продемонстрировано, что все разнообразие известных в биологии трофических процессов на микро-, макро-
Общность цели
Общность цели А со стороны реверса нужно обратить внимание на то, что природа у нас в одном лице. Это доказывается тем, что в любом ее закутке, который человек сумел обследовать, присутствует одно и то же вещество, одна и та же материя, и везде действуют единые законы