Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 5
ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Вся сложная и разнообразная деятельность высших отделов нервной системы построена на работе двух основных нервных процессов — возбуждения и торможения. Протекая в подвижных пространственных и временных отношениях друг с другом, эти процессы то разливаются (иррадиируют), то сосредоточиваются (концентрируются), то возбуждение порождает торможение (отрицательная индукция), то торможение дает начало возбуждению (положительная индукция). Непрерывное взаимодействие движущихся и вызывающих друг друга возбудительных и тормозных процессов создает в высших отделах мозга чрезвычайно тонкую мозаику, колеблющийся узор из переплетения возбужденных и заторможенных нейронов. Такие мозаики лежат в основе как различных актов поведения, так и их торможения в явлениях сна.
Иррадиация и концентрация нервных процессов
Возбуждение или торможение, возникшие в какой-либо клетке или группе клеток мозга, не остаются неподвижными, а всегда склонны к распространению. Распространение нервного процесса из очага его возникновения на окружающие нервные клетки называется иррадиацией.
Иррадиация торможения. Иррадиацию условного торможения удобно наблюдать в кожном анализаторе. Широко развернутая кожная поверхность тела является как бы увеличивающим зеркалом, в котором можно ясно видеть, как по последовательно расположенным проекционным полям будет иррадиировать тормозное состояние, например дифференцировочное торможение.
Иррадиацию дифференцировочного торможения обнаружили в следующем опыте.
Вдоль задней ноги собаки от стопы до бедра наклеивали пять «касалок» — приборов для механического раздражения кожи (рис. 35). Четыре верхние касалки использовали как сигналы условных пищевых слюноотделительных рефлексов. Эти рефлексы укрепляли, пока их слюноотделительный эффект не стал приблизительно одинаковым. Нижняя касалка служила дифференцировочным раздражителем и применялась без подкрепления едой, пока не перестала вызывать даже малейшего слюноотделения. Если теперь, вслед за применением дифференцировочной касалки, попробовать положительные раздражители, то оказывается, что слюногонное действие последних претерпевает закономерные изменения.
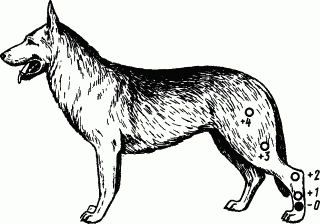
Рис. 35. Опыт с иррадиацией дифференцировочного торможения по корковым клеткам кожного анализатора:
0 — дифференцировочный раздражитель, 1, 2, 3, 4 — положительные условные раздражители (прикладываются к пунктам кожи ноги на расстоянии от дифференцировочного раздражителя соответственно на 3, 9, 15 и 22 см)
В классических опытах Н.Н. Красногорского и Б.А. Когана условный рефлекс на каждую из положительных касалок составлял 5 капель слюны за 30 с. Но каждый раз, когда дифференцировочная касалка создавала очаг торможения, начинали изменяться и соседние положительные рефлексы. Следовательно, торможение выходит за пределы своего очага и захватывает соседние клетки анализатора, в данном случае те, на которые проецируются кожные пункты положительных касалок. Далее видно, что в одинаковых условиях минутного интервала после троекратного действия дифференцировочной касалки условные рефлексы с положительных касалок изменяются по-разному, в зависимости от расположения последних относительно дифференцировочной. Так, рефлекс с ближайшего пункта (касалка 1) оказался полностью заторможенным. Рефлекс с пункта, расположенного несколько дальше (касалка № 2), был только уменьшен. Рефлексы с остальных пунктов, расположенных еще дальше, не только не испытали торможения, но даже усилились. Следовательно, иррадиирующее торможение оказывает тем более сильное воздействие на клетки анализатора, чем ближе они находятся к тормозному очагу. По своей природе иррадиация торможения — это развитие ТПСП на все большей территории коры, но не истинное движение процесса.
Концентрация торможения. После широкой иррадиации наступает сосредоточение, концентрация торможения в месте своего возникновения. Этот процесс тоже удобно проследить на примере дифференцировочного торможения в кожном анализаторе. Опыты проводили так же, как и при наблюдении иррадиации, но положительные рефлексы с каждого пункта кожи испытывали в различные сроки после окончания действия тормозного раздражителя. При помощи такого приема можно увидеть, как сначала далеко распространившееся тормозное состояние начинает сосредоточиваться, возвращаясь к исходному пункту.
При своем концентрировании торможение проходит в обратной последовательности все те пункты проекционных полей анализаторов, которые оно захватывало в своем поступательном движении.
Что собой представляет процесс концентрации торможения? Здесь не исключены две возможности. Первая заключается в том, что распространившееся торможение рассеивается, затухает на периферии и занимаемая им территория постепенно уменьшается. Вторая возможность — это подъем обратной волны торможения к тому месту, откуда оно распространилось. Последнее более вероятно, как, например, упрочение дифференцировки сопровождается усилением тормозного процесса. Следовательно, концентрация торможения связана не с рассеиванием и ослаблением, а с его сосредоточением и усилением.
Скорость иррадиации и концентрации торможения в коре головного мозга собаки. Скорость иррадиирования тормозного состояния удалось измерить в кожном анализаторе. Для этого определяли время развития вторичного торможения условного рефлекса на раздражение пункта кожи, находящегося в известном удалении от первично угашаемого. Опыты проводили следующим образом. В разных местах тела собаки приклеивали к коже касалки. На все касалки вырабатывали условные пищевые слюнные рефлексы и укрепляли их до одинаковой величины. Затем наносили повторные раздражения какой-либо одной касалкой без подкреплений до тех пор, пока не достигали исчезновения слюноотделения. Как только эта касалка проявляла свое тормозное действие, испытывали состояние рефлекса с другого пункта. На основании ряда таких испытаний в разные сроки после вызова угасательного торможения и при разных расстояниях испытуемых пунктов от его очага определяли время иррадиирования (Б.А. Коган).
В этом опыте вторично затормаживаемые пункты испытывали медленно после раздражения, давшего нулевой результат, т.е. без всякого промежутка времени между ними. В этот момент, как видно, торможение из пункта, соответствующего правой стороне груди еще не успело распространиться дальше ближайших участков. До области, например, правой предплюсны оно еще не дошло. Однако постепенно увеличивая промежутки времени от тормозного раздражения до момента испытания рефлекса, можно уловить момент, когда торможение дойдет до пункта, соответствующего правой предплюсне. Оказалось, что процесс иррадиирования торможения по нервным клеткам коры протекает очень медленно. Для прохождения области одного только кожного анализатора торможению требуются минуты.
Продолжая увеличивать промежуток времени между моментом приложения тормозного раздражителя к одному пункту и моментом испытания рефлекса с другого пункта, можно определить и скорость концентрирования угасательного торможения в кожном анализаторе.
Абсолютные величины времени концентрации тормозного процесса, как и времени его иррадиации, сильно зависят от индивидуальных особенностей подопытных животных, однако их соотношение оказалось довольно постоянным у всех испытуемых собак. Как правило, иррадиирование происходит в 4–5 раз быстрее, чем последующее концентрирование.
Иррадиация возбуждения. Опыт, показывающий иррадиацию возбудительного процесса, в некоторых отношениях напоминает описанные выше опыты с иррадиацией торможения.
У собаки вдоль задней ноги от плюсны до таза на примерно одинаковом расстоянии друг от друга приклеивали пять касалок (рис. 36). На действие самой нижней касалки (№ 1) вырабатывали условный рефлекс слюноотделения, подкрепляемый вливанием раствора кислоты в рот собаки. При первом испытании и другие касалки (№ 2, 3, 4 и 5) как сходные раздражители тоже вызывали слюноотделение. Поэтому необходимо было выработать дифференцирование этих касалок, применяя многократно касалку № 1 с подкреплением, а остальные касалки — без подкрепления. Теперь только касалка № 1 вызывала слюноотделение, а остальные превратились в тормозные сигналы.
После такой подготовки приступали к главной части опыта. Включали положительную касалку № 1 на 15 с и сразу после ее выключения действовали отдифференцированной касалкой № 2. Однако ее действие тоже вызывало слюноотделение. Это означало, что пункт кожного анализатора, соответствующий касалке № 2, обычно находящийся в тормозном состоянии, сразу после возникновения очага возбуждения в пункте, соответствующем касалке № 1, тоже оказался в возбужденном состоянии. Иначе говоря, возбуждение из пункта касалки № 1 в это время распространилось на пункт касалки № 2. Если испытать таким образом и какой-либо другой, более удаленный пункт кожного анализатора, то можно судить о районе такой иррадиации. Эти опыты показали, что иррадиирующее возбуждение по мере удаления от очага своего развития постепенно ослабевает.

Рис. 36. Опыт с иррадиацией возбуждения по корковым клеткам кожного анализатора:
1 — положительный условный раздражитель, 2, 3, 4, 5 — дифференцировочные раздражители
В другом варианте опытов под наблюдение была взята более обширная область кожного анализатора. Касалки располагали от конца передней лапы через все туловище до конца задней лапы. Первая касалка на конце передней лапы была положительным сигналом, а все остальные — дифференцировочными. Испытания дифференцировочными касалками показали, что уже через 0,5–1 с по прекращении положительного сигнала возбудительный процесс обнаруживается на соседних пунктах.
Таким образом, иррадиация возбуждения в коре мозга происходит гораздо быстрее, чем иррадиация торможения, и требует менее секунды для распространения по области кожного анализатора.
Через некоторое время после положительного сигнала соседние пункты анализатора вновь оказываются в прежнем тормозном состоянии. Это означает, что волна возбуждения успела уже разлиться по коре и вновь сосредоточиться в исходном пункте.
Индукция нервных процессов
Движение основных процессов высшей нервной деятельности определяется не только свойствами иррадиации и концентрации, но и свойствами их взаимной индукции. Индукцией называется свойство каждого из основных нервных процессов вызывать вокруг себя и после себя противоположный процесс.
Положительная индукция из очагов угасательного и дифференцировочного торможения. В опытах с иррадиацией угасательного торможения по кожному анализатору часто наблюдают такое явление: повторение раздражения какой-либо одной касалкой без подкрепления способствует полному угасанию условного рефлекса с раздражаемого пункта кожи. Однако сразу после этого раздражение касалкой с другого отдаленного пункта выявляет этот рефлекс в резко усиленном виде. Возникший в первом пункте очаг сильного торможения индуцировал в отдаленном пункте возбуждение, привел этот пункт в состояние повышенной возбудимости.
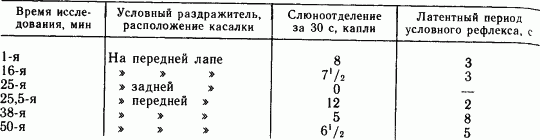
Еще более четко явление положительной индукции было выявлено в специальных опытах на примере индукции, развиваемой дифференцировочным торможением. Так, у собаки вырабатывали условный пищевой рефлекс слюноотделения, в котором сигналом служило раздражение касалкой кожи передней лапы. Другая касалка была установлена на задней ноге. Ее применяли без подкреплений, так что скоро она приобрела тормозное значение дифференцировочного раздражителя. На включение дифференцировочной касалки слюноотделения не наступало, однако испробованный сразу после нее положительный раздражитель давал резко усиленный рефлекс (табл. 7).
Измерение силы условного рефлекса количеством слюны обнаруживает, что торможение в пункте задней лапы усилило условное возбуждение в пункте передней лапы почти на 50 %. Следовательно, в данном случае положительная индукция произошла из очага торможения в дальнем пункте анализатора.
Таблица 7. Положительная индукция из очага дифференцировочного торможения в кожном анализаторе (по Д.С. Фурсикову, 1922)
Однако мозг часто дифференцирует раздражители, связанные с одним и тем же пунктом анализатора, но отличающиеся друг от друга по силе или характеру воздействия. Будет ли в таких случаях проявляться положительная индукция? Ответ на этот вопрос дает следующий опыт. У собаки был выработан условный пищевой рефлекс на сильный свет, от него отдифференцировали слабый свет. Затем сильный свет был испробован сразу после слабого. И здесь условный слюнной рефлекс, вызванный сразу после дифференцировочного, увеличился почти на 50 %. Следовательно, в данном случае положительная индукция произошла из очага торможения в том же пункте анализатора.
Таким образом, положительная индукция может проявляться в различных анализаторах и при разных отношениях тормозного очага и положительно индуцируемого рефлекса.
Отрицательная индукция из очага возбуждения в очаг торможения. Явления отрицательной индукции можно продемонстрировать в следующем опыте. У собаки образован условный пищевой рефлекс на метроном 120 ударов/мин. К этому положительному раздражителю выработана дифференцировка метронома 60 ударов/мин. Как известно, дифференцировку очень легко разрушить, если начать сопровождать дифференцировочный раздражитель подкреплением. И действительно, после того как несколько раз метроном 60 ударов/мин применили с подкармливанием, он сам начал вызывать слюноотделение. Это простой и безотказный способ уничтожения тормозного очага.
Однако при помощи некоторых средств можно задержать разрушение дифференцировки, т.е. продлить существование очагов условного торможения. В частности, таким средством оказалось применение положительных сигналов, т.е. создание очагов условного возбуждения. Это видно из следующих опытов.
Например, собаку подкармливают после каждого применения метронома с частотой 60 ударов/мин до тех пор, пока у нее не начнет сильно выделяться слюна (разрушение дифференцировки). Тогда применяют с подкреплением один раз метроном с частотой 120 ударов/мин. В результате используемый вслед за ним метроном <...часть числа не видна…>0 ударов/мин, который только что вызывал слюноотделение, сразу теряет свое действие. Дифференцировка при этом восстанавливается, что связано с возникновением рядом очага возбуждения. Этот очаг отрицательно индуцировал, т.е. затормозил клетки пункта метронома с частотой 60 ударов/мин, и индукционное торможение усилило остатки дифференцировочного.
Таким образом, положительный сигнал благодаря отрицательной индукции укрепляет свое дифференцирование от близких отрицательных сигналов.
Мозаика возбуждения и торможения в высших отделах нервной системы. Взаимодействие иррадиирующих и индуцированных нервных процессов создает необычно сложное и меняющееся от момента к моменту их уравновешивание и территориальное разграничение. В результате возбуждение и торможение образуют дробный рисунок подвижной мозаики, непрерывно меняющей свои очертания.
В свое время И.П. Павлов говорил о том, какую замечательную картину вспыхивающих и затухающих, непрерывно перемежающихся мерцаний мы увидели бы на поверхности мозга, если бы его возбужденные пункты светились.
Эта мысль Павлова получила осуществление при изучении движения нервных процессов по коре больших полушарий с помощью методики электроэнцефалоскопии (М.Н. Ливанов, В.М. Ананьев, 1960). Электроэнцефалоскоп позволяет наблюдать мозаику электрической активности коры мозга при одновременном отведении из <…часть числа не видна…>00 ее пунктов и воспроизводит на экране телевизионной трубки непрерывно возникающие и меняющиеся подвижные картины, которые фиксируются киносъемкой (рис. 37). Такой «телевизор» мозга значительно расширяет возможности объективного изучения пространственной динамики активности коры при условно-рефлекторной деятельности.

Рис. 37. Перераспределение очагов активности в коре мозга кролика при выработке условного двигательного рефлекса на зрительное раздражение (по М.Н. Ливанову):
кадры киносъемки из опыта с сочетаниями вспышек света и изоритмических электрокожных раздражений лапы, яркость каждой точки на экране топоскопа отражает величину электрической активности этого пункта в данный момент (обратить внимание на постепенную концентрацию активности в районе двигательного и зрительного анализаторов)
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.6. Типы высшей нервной деятельности
6.6. Типы высшей нервной деятельности Условно-рефлекторая деятельность зависит от индивидуальных свойств нервной системы. Индивидуальные свойства нервной системы обусловлены наследственными особенностями индивидуума и его жизненным опытом. Совокупность этих свойств
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ С незапамятных времен внимание человека привлекали проявления психической жизни. Откуда берутся мысли? Как понимать сознание, вмещающее весь мир и управляющее нашими поступками? Что
Глава 8 ТИПОЛОГИЯ И ГЕНЕТИКА ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 8 ТИПОЛОГИЯ И ГЕНЕТИКА ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Нервная деятельность, как мышечная сила, рост, цвет глаз и прочие признаки, может быть индивидуально различной. Эти различия во многом зависят от наследственности. В свою очередь, наследственность формируется
Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА Гибкий и подвижный механизм высшей нервной деятельности обнаруживает громадные приспособительные возможности нормального функционирования при самых разных состояниях организма.Знание
ГЛАВА 3. УПРАВЛЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТЬЮ (ПОВЕДЕНИЕМ)
ГЛАВА 3. УПРАВЛЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТЬЮ (ПОВЕДЕНИЕМ) Поведение собаки, т.е. сложная совокупность ее взаимодействий с окружающей средой, привычно рассматривается человеком с позиций собственного душевного мира. Такой подход к психике животных хорошо объясним и,
3.10. НАРУШЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
3.10. НАРУШЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Сочетание нейрохирургических вмешательств в определенные отделы мозга и условно-рефлекторного метода привело к пониманию многих сторон высшей нервной деятельности. Анализ ее нарушений расширил представления о работе мозга в
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 20 ]
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 20 ] Второй раз я имею высокую честь и глубокое удовлетворение принимать участие в торжестве открытия деятельности чрезвычайных русских обществ, народившиеся здесь, в Москве, и свидетельствующих о том, как
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 25 ]
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 25 ] Прежде всего я считаю своим долгом благодарить Философское общество, что оно в лице своего председателя изъявило готовность выслушать мое сообщение. Мне трудно было сообразить, насколько это
ЭКСПЕРИМЕНТАЛЬНАЯ ПАТОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ [ 51 ]
ЭКСПЕРИМЕНТАЛЬНАЯ ПАТОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ[ 51 ] Несколько вступительных слов о сложной судьбе нашей работы по физиологии и патологии высшей нервной деятельности, принимая, что прилагательные «высшая нервная» отвечают прилагательному
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1 Образы, картины поведения как нас самих, так и близких к нам высших животных, с которыми мы находимся в постоянных жизненных отношениях (как, например, собак), представляют огромное разнообразие, прямо
ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ В СВЯЗИ С НЕВРОЗАМИ И ПСИХОЗАМИ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ НЕВРОТИЧЕСКИХ И ПСИХОТИЧЕСКИХ СИМПТОМОВ [ 52 ]
ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ В СВЯЗИ С НЕВРОЗАМИ И ПСИХОЗАМИ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ НЕВРОТИЧЕСКИХ И ПСИХОТИЧЕСКИХ СИМПТОМОВ[ 52 ] Из огромного материала, относящегося к изучению высшей нервной деятельности у собак по методу условных рефлексов, я остановлюсь
6. Типы высшей нервной деятельности
6. Типы высшей нервной деятельности На основании изучения условно-рефлекторной деятельности огромного числа собак академик И. П. Павлов создал свое учение о типах высшей нервной деятельности. Все многообразные особенности условно-рефлекторной деятельности собак
7. Патология высшей нервной деятельности
7. Патология высшей нервной деятельности Одной из сторон работ академика И. П. Павлова, имеющей большое значение для служебного собаководства, является развитие им учения о патологии нервной деятельности.Высшая нервная деятельность собаки может быть сорвана
Некоторые актуальные вопросы генетики поведения и высшей нервной деятельности[50]
Некоторые актуальные вопросы генетики поведения и высшей нервной деятельности[50] Исследования по генетике поведения начались почти с самого начала нынешнего столетия, когда законы Менделя стали достоянием широких кругов биологов.При этом выяснилось, что признаки