Глава 8 ТИПОЛОГИЯ И ГЕНЕТИКА ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 8
ТИПОЛОГИЯ И ГЕНЕТИКА ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Нервная деятельность, как мышечная сила, рост, цвет глаз и прочие признаки, может быть индивидуально различной. Эти различия во многом зависят от наследственности. В свою очередь, наследственность формируется свойствами, приобретаемыми в истории вида. Отбор и наследственное закрепление новых типологических признаков и форм поведения является механизмом эволюции высшей нервной деятельности.
Общие типы нервной системы
Повседневная жизнь дает немало примеров тому, как по-разному ведут себя отдельные животные и поступают разные люди в одной и той же обстановке. Несмотря на чрезвычайное разнообразие, эти индивидуальные особенности имеют некоторые общие черты. По общим чертам поведения намечаются общие типы нервной системы, которые определяют разные темпераменты животных и характеры людей.
Темперамент и характер. Знаменитый врач древности Гиппократ объяснял природу индивидуальных особенностей поведения различием пропорций «жизненных соков» тела: крови, слизи и желчи. До наших дней сохранилось понятие — темперамент (от лат. temperamentum — надлежащее соотношение, соразмерность). Большое количество внимательных наблюдений позволяло Гиппократу подметить и описать четыре действительно наиболее часто встречающихся в жизни вида темперамента: сангвинический, флегматический, холерический и меланхолический.
Гиппократ считал, что когда у человека из всех его соков преобладает пылкая кровь (sangvis), то его поведение дает нам черты сангвинического темперамента — энергию, настойчивость, решительность. Если же пылкую кровь охлаждает находящаяся в избытке слизь (phlegma), то получается флегматик — хладнокровный и медлительный. Едкая желчь (chole) способствует образованию раздражительного, вспыльчивого, не знающего меры холерического темперамента. Но когда накапливается много испорченной черной желчи (melan chole), то такой вялый меланхолик постоянно будет пребывать в унынии.
Выяснение истинных функций крови, желчи и других «соков» тела привело к появлению других предположений о природе разных темпераментов. Так, в начале XX в. широкое распространение среди врачей получили взгляды на темперамент как следствие определенного анатомического склада тела. Например, утверждали, что полнота располагает к добродушию, а худые люди бывают большей частью желчными и злыми. В таких утверждениях явно перепутаны причина и следствие. Действительно, веселые, жизнерадостные люди часто бывают полными, а хмурые, раздражительные — худыми. Но это далеко не правило, более того, нередко бывает наоборот. Поэтому нет оснований считать анатомический склад тела (конституцию) причиной того или иного темперамента. Как правило, черты темперамента и внешний облик человека в равной мере зависят от условий его жизни. Конечно, жизненные невзгоды и лишения не только вызывают физическое истощение, но и делают человека озлобленным и раздражительным. Понятно также, что спокойная жизнь в достатке, часто с излишествами в еде, располагает к полноте и к благодушному настроению. Не от телесного сложения зависит настроение, а от условий жизни, которые определяют то или иное развитие наследственных задатков темперамента, и конституции. Поэтому лишены всякого научного основания попытки судить о темпераменте, а тем более о характере человека по каким-либо анатомическим показателям строения его тела.
Настоящая природа темперамента и характеров была раскрыта И.П. Павловым (1927) с помощью учения о высшей нервной деятельности. Многочисленные наблюдения и специальные опыты на лабораторных животных убедительно показали, что разные темпераменты есть не что иное, как проявление индивидуальных различий в свойствах протекания возбудительных и тормозных процессов в высших отделах мозга. Различия темпераментов отражают различия общих типов нервной системы, свойственных и человеку, и животным. Характер человека складывается из его прирожденного темперамента плюс изменения темперамента, обусловленные воспитанием, плюс сумма сложнейших отношений с окружающими людьми и событиями. (Более подробно о физиологических предпосылках формирования характера человека речь будет идти в гл. 17.)
Индивидуальные различия в протекании условных рефлексов. Проводя эксперименты на большом числе лабораторных собак, И.П. Павлов обратил внимание на то, что все они «работают» по-разному. Одни собаки легко и быстро, после нескольких сочетаний, образуют прочные условные рефлексы. Другие вырабатывают условный рефлекс с большим трудом, медленно, лишь после многих десятков сочетаний. Но даже когда они выработают рефлекс, то последний оказывается непрочным и слабым. Он то появляется, то исчезает. Несмотря на частые подкрепления, он остается неустойчивым, небольшим по величине и легко разрушается. Такая разница в образовании условных рефлексов обусловливается разной способностью к образованию крепких временных связей, т.е. разной силой условного возбуждения. Так выявился первый показатель типологических различий высшей нервной деятельности — сила возбудительных процессов. Показатель силы возбудительных процессов позволяет объективно оценить способность корковых клеток к развитию более или менее сильного условного возбуждения.
Однако если отобрать для дальнейших опытов собак с примерно одинаковой силой возбудительных процессов, то среди них почти обязательно проявятся другие индивидуальные различия условно-рефлекторной деятельности. Эти различия обнаружатся, как только у собак начнут вырабатывать дифференцировки условных раздражителей. Здесь может случиться, что две собаки, одинаково быстро и прочно образовавшие положительный условный рефлекс, например, на метроном с частотой 60 ударов/мин, резко разойдутся в своих способностях отдифференцировать от него неподкрепляемый сигнал — метроном с частотой 120 ударов/мин. Одна собака уже после нескольких применений метронома 120 ударов/мин без подкармливания перестает на него реагировать. Другая — не может угасить рефлекс на этот дифференцировочный раздражитель. Также плохо идет у нее выработка запаздывания, условного тормоза и угасания, т.е. разновидностей условного торможения. Такое различие в выработке тормозов говорит о разной способности к концентрированию возбудительного процесса, т.е. о разной силе условного торможения.
Так выявился второй показатель типологических различий высшей нервной деятельности — сила тормозных процессов. Показатель силы тормозных процессов позволяет объективно оценить способность корковых клеток к развитию более или менее сильного условного торможения.
Однако даже среди собак, проявивших одинаковые способности к развитию условного возбуждения и торможения, опять обнаруживаются индивидуальные различия условно-рефлекторной деятельности. Эти различия особенно ярко выявляются при переделках сигнального значения условных раздражителей, что свидетельствует о разной способности к замене возбудительного процесса тормозным и наоборот, т.е. о разной подвижности возбуждения и торможения. Так выявился третий показатель типологических различий высшей нервной деятельности — подвижность нервных процессов. Показатель подвижности нервных процессов позволяет объективно оценить способность корковых клеток к быстрому и полному переходу из возбужденного в тормозное состояние и обратно.
Итак, различия в свойствах условно-рефлекторной деятельности разных животных оказываются результатом индивидуальных вариаций силы возбуждения, силы торможения и их подвижности. Однако для проявления конечных результатов в поведении важна не столько абсолютная сила тормозного процесса, сколько его отношение к возбудительному. Например, резкий перевес возбуждения помешает даже сильному торможению в выработке дифференцировки. А без такой помехи даже более слабое торможение успешно справится с этой задачей. Поэтому вместо показателя абсолютной силы торможения в типологии обычно используют сравнительный показатель силы торможения относительно возбуждения, или показатель их уравновешенности. Тогда индивидуальные различия протекания процессов высшей нервной деятельности можно оценивать по трем показателям: силы, уравновешенности и подвижности. Следовательно, корковые процессы могут быть сильными или слабыми, уравновешенными или неуравновешенными, подвижными или инертными.
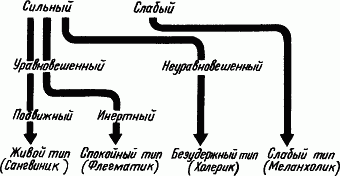
Рис. 57. Основные типы высшей нервной деятельности
Основные общие типы нервной системы. Из набора разных свойств — силы или слабости, уравновешенности или неуравновешенности, подвижности или инертности нервных процессов — в природе складывается множество комбинаций. При этом из всех возможных комбинаций наиболее четко вырисовываются четыре. Это и есть главные общие типы нервной системы, подмеченные еще Гиппократом в четырех темпераментах людей.
Какие свойства протекания процессов высшей нервной деятельности характеризуют четыре главных общих типа нервной системы (рис. 57)?
Первое, наиболее часто встречающееся в жизни сочетание свойств нервных процессов заключается в их удовлетворительной силе, хорошей уравновешенности и достаточной подвижности. Хорошие сила, уравновешенность и подвижность нервных процессов характеризуют живой тип по Павлову, соответствующий сангвинику Гиппократа. В лабораторных опытах такой темперамент проявляется в скором и прочном образовании условных рефлексов (сила), легкости выработки дифференцировок (уравновешенность) и осуществлении переделок (подвижность).
В природном поведении животных черты этого нервного типа выступают как активность и стойкость в борьбе (сила), как большая выдержка, например, у подстерегающих добычу (уравновешенность) , как способность быстро менять свои жизненные навыки при изменениях обстановки (подвижность). В характере человека черты живого типа проявляются энергией и настойчивостью в преодолении трудностей (сила), большим самообладанием, умением «держать себя в руках» (уравновешенность) и вместе с тем, в случае необходимости, быстрой перестройкой своих привычек и привязанностей (подвижность). Такие люди очень работоспособны, особенно когда есть интересующее их дело, и упорны в достижении цели. Они спокойно принимают перемены в жизни и умеют осваиваться в новой обстановке.
Другое часто встречающееся в жизни сочетание свойств нервных процессов — это их хорошая сила и уравновешенность, но малая подвижность. Хорошие сила и уравновешенность, но инертность нервных процессов характерны для спокойного типа, по Павлову, соответствующего флегматику Гиппократа. В лабораторных опытах такой темперамент проявляется в скором и прочном образовании условных рефлексов (сила) и легкости выработки дифференцировок (уравновешенность), однако переделки сигнального значения условных раздражителей идут с большим трудом (малая подвижность). В природе у животных черты спокойного типа выступают в том, что большая настойчивость (сила) и выдержка (уравновешенность) могут сочетаться с некоторой косностью поведения, неспособностью к быстрой переделке, например охотничьих навыков при изменении повадок добычи (малая подвижность).
В характере человека черты флегматического темперамента проявляются прежде всего неторопливостью. Наряду с энергией и работоспособностью (сила), самообладанием и хорошим «чувством меры» (уравновешенность) обнаруживается большая сила привычек, некоторая медлительность в принятии решений, особенно при неожиданных событиях (малая подвижность). Такие люди не склонны к поспешности, им чужды суетливость и готовность во всем следовать велениям момента. Флегматик трудно сходится с новыми людьми, но если сойдется, то будет верным другом. Это ровный и спокойный, упорный труженик жизни.
Следующее, нередко встречающееся сочетание свойств нервных процессов — это их удовлетворительная сила при неуравновешенности. В этом случае о подвижности вообще ничего сказать нельзя. Неуравновешенность обычно выражается в преобладании возбудительного процесса над тормозным. Сила, но неуравновешенность с преобладанием возбуждения характерны для безудержного типа, по Павлову, соответствующего холерику Гиппократа. В лабораторных опытах такой темперамент проявляется также скорым и прочным образованием условных рефлексов (сила), но дифференцировки у этих собак вырабатываются очень плохо. В природном поведении животных безудержный темперамент проявляется в сочетании смелости (сила) с отсутствием выдержки: например, нападает на крупного зверя и при подстерегании добычи раньше времени выскакивает из засады.
В характерах людей черты безудержного типа дают себя знать в самозабвенных увлечениях, когда человек, например, может делать громадную работу «запоем» (сила), а потом с таким же азартом ухватится за другое дело и здесь тоже «не знает меры». Они проявляются и в отношениях с людьми, когда какой-нибудь пустяк может вызвать взрыв гнева, в котором безудержный человек сам потом будет раскаиваться (неуравновешенность с преобладанием возбуждения).
Наконец, последний, сравнительно редко встречающийся темперамент, определяется слабостью и возбудительных и тормозных процессов. Слабость нервных процессов и быстрый переход корковых клеток в состояние запредельного торможения характерны для слабого типа, по Павлову, соответствующего меланхолику Гиппократа. В лабораторных опытах этот нервный тип проявляется в крайней нестойкости условных рефлексов, невозможности их образования на сигналы большой силы, так как всякое сильное раздражение вызывает общее угнетение ввиду запредельного торможения. Наблюдая поведение таких животных, их называют трусливыми. Эти животные избегают сильных раздражений, при встрече с врагом спасаются бегством, не проявляют настойчивости, например, быстро отказываются от затянувшегося преследования, решительным сопротивлением и даже просто громким окриком их можно вынудить к отступлению (слабость нервных процессов).
Черты меланхолического темперамента проявляются у человека прежде всего в том, что называют «слабость характера», т.е. в легком подчинении чужой воле, неспособности настоять на своем. Сильные жизненные раздражители оказываются для таких людей сверхсильными, в чрезвычайных обстоятельствах эти люди впадают в панику, жизнь представляется им состоящей из непреодолимых трудностей, полной тоски и уныния (слабость нервных процессов). Такие люди стремятся оградить себя от жизни с ее волнениями, избегают общества, боятся всякой ответственности. Яркий художественный образ меланхолика — «человека в футляре» — нарисовал А.П. Чехов в рассказе о робком учителе греческого языка, который всю жизнь боялся, «как бы чего не вышло».
Таковы четыре главных, встречающихся в жизни общих для человека и животных типа нервной системы. Однако в столь крайнем выражении, как они сейчас описаны, эти типы встречаются редко. Большей частью в жизни существуют промежуточные формы с преобладанием свойств того или другого типа.
Существует довольно распространенное мнение, что при определении типа нервной системы как бы оценивают ее «сорт»: например, сангвиник — обладатель первосортной нервной системы, а меланхолик — представитель последнего сорта. Такое мнение является ошибочным, далеким от действительности. Нет оснований мерить социальную ценность людей по их темпераментам. В разных областях деятельности могут оказаться нужными разные характеры. Если, например, отобрать у Чапаева некоторые черты его безудержного темперамента, то вряд ли он мог бы увлечь своих бойцов в лихом ударе по врагам. Но, чтобы терпеливо распутывать интриги врагов в дипломатических салонах, полезно обладать хорошей уравновешенностью и большой выдержкой.
Главная причина того, что темперамент не определяет социальной ценности человека, заключается в руководящей роли человеческого сознания. Образуя специальную надстройку высшей нервной деятельности в виде второй сигнальной системы, сознание человека подчиняет себе все поведение. Чувство долга удерживает слабохарактерного от соблазна, воспитание помогает неуравновешенному держать себя в руках, порывы темперамента сдерживаются голосом человеческого разума.
Генотип и фенотип. Наблюдая в течение длительного времени за животным, например за щенком, вырастающим во взрослую собаку, нетрудно заметить, как меняется его поведение. Но что тогда считать свойствами, характеризующими тип нервной системы?
Чтобы ответить на этот вопрос, понятие о типе нервной системы приходится усложнить, разделив его на понятие генотипа и фенотипа. Генотип характеризуется врожденными, переданными по наследству свойствами процессов высшей нервной деятельности. Фенотип характеризуется теми свойствами, которые сложились из взаимодействия врожденных и приобретенных под влиянием условий жизни.
Строго говоря, настоящий генотип животного обычно ускользает от определения, так как условия жизни начинают действовать на него, начиная с момента появления организма на свет. Однако исходя из стойкости генотипических признаков, считают, что их можно обнаружить и во взрослом состоянии, если с животным не произошло за это время ничего чрезвычайного, что могло бы вызвать резкие изменения в его нервной системе. Например, тяжелые нервные потрясения могут ослабить корковые клетки и привести к появлению черт меланхолического темперамента у животного сангвинического генотипа. Но и без всяких чрезвычайных происшествий жизнь непрерывно вносит в поведение дополнения и изменения. Жизненная обстановка, благоприятствующая тренированию внутреннего торможения, может несколько сгладить его холерическую неуравновешенность с возбуждением. Опыты показали, что если многократно переделывать сигнальные значения условных раздражений, то каждая последующая переделка наступает легче и быстрее. Следовательно, есть основания ожидать, что жизнь, богатая переменами, может помочь преодолению черт флегматичности. Все эти изменения свойств протекания нервных процессов, вносимые жизнью, накапливаются в фенотипе. Условия воспитания могут замаскировать истинные свойства нервного типа.
Многочисленные эксперименты, проведенные на животных разных видов, показали влияние условий жизни на формирование фенотипа. Например, подсчитывали число спариваний за 30 минут особей видов-двойников Drosophila melanogaster и Drosophila simulans, выращенных при разной температуре среды (от 12 до 30 °С). Наблюдения вели на 10 линиях, каждая из которых брала начало от одной оплодотворенной самки, т.е. имела один генотип. Тем не менее половое поведение мух оказалось различным в зависимости от температурных условий среды их развития. Из наблюдений за поведением позвоночных животных весьма демонстративны опыты с мышами, которые должны были отыскивать пищу. Оказалось, что особи всех шести инбредных линий при воспитании в условиях разнообразия обстановки, обозначаемого как «обогащенная среда», быстрее находили пищу, чем воспитанные в стандартных, более однообразных условиях.
Описаны наглядные примеры влияния воспитания на поведение птиц. Так, если воробьиных овсянок, предпочитающих ветви дуба, вырастить в лаборатории на ветвях сосны, то они будут предпочитать сосну. Птенцы воробьев, воспитанные в среде канареек, вместо врожденного чириканья начинают издавать звуки, похожие на пение канареек. Общеизвестны многочисленные случаи, когда дикие животные, даже хищники, воспитанные с раннего возраста человеком, коренным образом изменяли свой генотип поведения и становились ручными.
Рассмотренные выше примеры показывают влияние условий жизни, которые, не соответствуя требованиям генотипа, формируют фенотип, отклоняющийся от генетической программы поведения. Однако могут складываться такие условия жизни, которые не приходят в противоречие, а, наоборот, способствуют осуществлению программы генотипа. Эти условия создаются, например, для особей, нашедших для себя экологически оптимальные регионы обитания, или сельскохозяйственных животных, получающих тем больше пищи, чем выше их продуктивность.
О значении воспитания свидетельствует следующий чрезвычайно наглядный опыт.
Щенков одного помета разделили на две группы и стали воспитывать по-разному. Одни росли на свободе, подвергались действию природных раздражителей, накапливая жизненный опыт. Другие — были изолированы от окружения и росли в уединении, лишенные естественного разнообразия раздражителей, не имея повода для активной деятельности. Когда те и другие щенки выросли, их объединили. Разница в поведении животных была очевидна. Собаки, выросшие на свободе, вели себя гораздо более активно и непринужденно, чем воспитанные в изоляции.
Результаты этого опыта поучительны. Они убедительно показывают важность жизненной закалки характера и пагубность так называемого «оранжерейного» воспитания. Об этом всегда должны помнить родители и педагоги.
Определение типов нервной деятельности у животных
Необходимость в определении типа нервной деятельности животного возникает во многих случаях: и при изучении свойств условных рефлексов с учетом индивидуальных вариаций, и при решении практических вопросов рационального использования животных и увеличения их хозяйственной продуктивности.
Полная оценка типа нервной деятельности по классической методике слюнных рефлексов. В лабораториях И.П. Павлова на основании слюнных условных рефлексов были разработаны специальные пробы для определения типа нервной деятельности животных. Из этих проб со временем в Институте физиологии им. И.П. Павлова АН СССР составили так называемый «Большой стандарт» испытаний для диагностики функционального типа нервной системы.
Однако испытания условных слюноотделительных рефлексов по программе «Большого стандарта» занимают около двух лет, и хотя эта программа позволяет получить точное и полное представление о типе нервной системы исследуемой собаки, длительность эксперимента делает ее малоприемлемой. Чтобы получить сведения о типологических различиях собак в более короткие сроки, необходимо несколько сократить всесторонность типологического обследования, тем более что многие из перечисленных в «Большом стандарте» испытаний имеют ценность лишь при сопоставлении с результатами других испытаний. Поэтому выявление сравнительных данных в показателях силы, уравновешенности и подвижности процессов высшей нервной деятельности способствовало подбору минимума наиболее надежных проб, из которых составили так называемый «Малый стандарт» испытаний.
Подготовка собаки к испытаниям строго регламентирована. Вначале ее приучают есть из кормушки, затем вырабатывают условные рефлексы в определенном порядке, наконец, закрепляют стереотип из положительных и отрицательных сигналов. Стереотип состоит из положительных раздражителей — звонка, света, тона, шума, затем дифференцировочного тона и, в заключение, опять положительного раздражителя — звонка. Затем приступают к испытаниям по специальной программе.
Программа «Малого стандарта» для определения типа высшей нервной деятельности собаки состоит из следующих испытаний.
1. Силу возбуждения оценивают по скорости выработки рефлекса и испытывают кофеином.
2. Силу торможения прежде всего оценивают по скорости выработки дифференцировки.
3. Силу торможения также испытывают продлением времени действия дифференцировки до 5 мин. Результаты этого испытания учитывают лишь тогда, когда дифференцировка выдерживается первые 20 с. Учитывают влияние такого испытания на последующие опыты.
4. Подвижность нервных процессов испытывают одновременной переделкой положительного сигнала в отрицательный, а отрицательного в положительный. В новом сигнальном значении раздражители применяют 30 раз, и лишь в исключительных случаях больше.
5. Подвижность нервных процессов оценивают также выработкой рефлекса с запаздыванием. Для этого новый сигнал средней силы (зуммер) вводят в стереотип на второе место взамен света.
Испытания свойств условно-рефлекторной деятельности по такой короткой программе «Малого стандарта» дают возможность в течение 6–7 месяцев с большой надежностью определить тип нервной системы собаки.
Оценка функционального типа нервной системы по двигательным искусственным условным рефлексам. «Малый стандарт» испытания условных слюноотделительных рефлексов, к сожалению, не всегда можно применить для определения типа нервной системы животного. Во-первых, его сложно использовать при типологическом исследовании животного в раннем возрасте (это и техническая сложность выведения слюнного потока у очень молодых животных, и изменение нервных процессов у животного в течение достаточно длительного срока испытания, так как животное вырастет за этот период). Во-вторых, изучение слюнных рефлексов, т.е. необходимость операции наложения фистулы протока, исключает возможность массовых определений типа нервной системы у животных для практических целей в различных областях.
Поэтому при типологических исследованиях в раннем возрасте, а также при изучении сельскохозяйственных животных широкое применение находят показатели не слюноотделительных, а двигательных условных рефлексов. В специальных опытах на собаках установлено, что испытания двигательных условных рефлексов дают результаты, несколько отличающиеся от результатов слюноотделительных рефлексов. Однако сила, уравновешенность и подвижность нервных процессов отражаются в динамике двигательных проявлений условных рефлексов столь же полно, как и в динамике их секреторных (слюноотделительных) проявлений. Сохраняют свое значение и те испытания, посредством которых выявляются свойства протекания возбудительных и тормозных процессов.
Типологические различия высшей нервной деятельности у щенков можно обнаружить в опытах с выработкой условного рефлекса подбегания к кормушке по сигналу. Для этого стенка ящика, в котором находятся щенки, откидывается и образует мостик к кормушке.
У одних щенков условное подбегание вырабатывалось быстро, происходило регулярно, с коротким латентным периодом, совершалось энергично. Это признаки хорошей силы, возбуждения. У других щенков выработка условного рефлекса задерживалась, подбегания наблюдались нерегулярно, с большими латентными периодами; щенки бежали к кормушке нерешительно, часто не добегали и поворачивали обратно. Это признаки слабости возбудительных процессов.
Продолжая такой опыт со щенками, можно вырабатывать у них дифференцирование условных раздражителей, а затем и переделку их сигнального значения. При этом четко выявляются индивидуальные различия силы торможения и подвижности нервных процессов.
Изучение типов нервной системы сельскохозяйственных животных производят главным образом при помощи двигательных пищевых рефлексов. Для различных животных разработана разная техника исследования этих рефлексов соответственно биологическим особенностям их питания.
Оценка типа нервной системы по двигательным натуральным условным рефлексам. Во всех описанных выше примерах определения типа нервной системы прежде чем приступить к испытаниям условных рефлексов (слюнных или двигательных) их нужно было вырабатывать. Именно из-за предварительной выработки искусственных условных рефлексов много времени уходит на повторные длительные опыты. При максимальном сокращении программы испытаний выработка условных рефлексов может занимать больше времени, чем сами испытания. К натуральным рефлексам применимы все приемы типологических испытаний, разработанные для искусственных. Результаты этих испытаний быстро, хотя и приблизительно, характеризуют состояние процессов высшей нервной деятельности.
Испытания натуральных условных рефлексов были применены, например, для быстрой оценки типологических различий служебных собак. Схема установки для проведения таких испытаний в условиях питомника служебного собаководства изображена на рис. 58. Если для оценки типа нервной системы испытывают пищевые условные рефлексы, то условным раздражителем служит привычный для служебной собаки бачок с едой. Сила возбудительных процессов характеризуется тем, насколько нарастает двигательная реакция при усилении условного пищевого раздражителя (приближение бачка с едой). Сила тормозных процессов характеризуется скоростью и полнотой угашения двигательной реакции на стоящий перед собакой бачок с едой. Подвижность нервных процессов приблизительно характеризуется ходом восстановления этой угашенной двигательной реакции. Все исследование длится несколько минут.
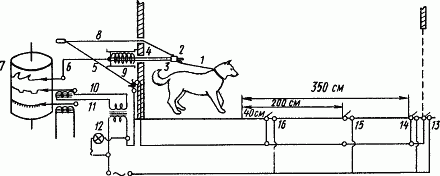
Рис. 58. Установка для быстрой оценки типологических различий служебных собак (по Э.А. Варухе):
1 — поводок от ошейника собаки, 2 — замок прикрепления поводка к стержню, 3 — муфта освобождения поводка, чтобы собака могла побежать за подкреплением, 4 — стержень, сжимающий пружину при рывках собаки, 5 — пружина в обойме, 6 — угловой рычаг, записывающий движение натурального условного рефлекса, 7 — кимограф, 8 — шнур для освобождения поводка с отметкой этого момента размыкателем (9), 11 — отметка времени, 13–16 — выключатели на полу для отметки раздражений в записи (10) и сигнальной лампочки (12) при испытании натурального условного рефлекса с разных расстояний
Типологическое значение получаемых показателей устанавливают однажды в данных условиях опытов по «эталонным» собакам, типы нервной системы которых были обнаружены по стандарту классической слюноотделительной методикой.
Если испытывать у собаки на той же установке не пищевой, а агрессивный натуральный условный рефлекс, то результаты такого испытания могут обнаружить иные типологические черты животного, в зависимости от его доминирующей мотивации. Следовательно, в разных системах рефлексов (в данном случае — пищевых и агрессивных) могут проявляться несколько различные свойства протекания нервных процессов, что необходимо учитывать, например, при отборе служебных собак с требуемыми типологическими качествами.
Генетика высшей нервной деятельности животных
Механизмы и свойства процессов высшей нервной деятельности формировались и совершенствовались на протяжении многовековой эволюции мозга. В бесчисленном ряду поколений природа отбирала и формировала генотипы прогрессивных форм поведения, обеспечивающих наибольшее приспособление к условиям жизни.
Генетика поведения. По своей роли в повседневном решении задачи выжить в изменчивой среде обитания поведение животного, являясь весьма динамичным, имеет первостепенное значение. Человек на протяжении многих поколений путем настойчивого отбора коренным образом преобразовывал поведение животных в соответствии со своими потребностями.
Закономерности наследования особенностей поведения в естественных условиях и при отборе изучали у животных разных классов и видов, стоящих на разных ступенях филогенеза (у насекомых, рыб, птиц, млекопитающих). Важные результаты получены при генетическом исследовании поведения насекомых. Например, при скрещивании двух рас пчел по показателям ритма сигнальных движений так называемого танца пчел (см. гл. II) выявилось доминирование материнских генов. При селекции мух по медленности спаривания доминирующими также оказались гены самок (рис. 59).
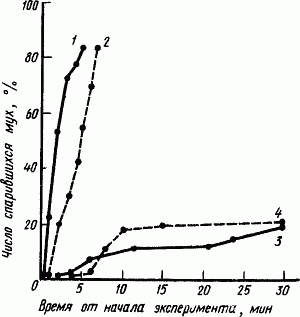
Рис. 59. Скорость спаривания мух (по А. Мэннингу):
1 — отобранные самцы и контрольные самки, 2 — контрольные самцы и контрольные самки, 3 — отобранные самцы и отобранные самки, 4 —контрольные самцы и отобранные самки
В других случаях (исследования на курах) могут доминировать отцовские гены. Так, например, в результате длительного отбора по признаку яйценоскости куры породы леггорн перестали насиживать яйца. Однако когда этих кур спаривали с петухами породы корниш, куры которой в подавляющем большинстве являются наседками, то 88 % кур первого поколения от такого скрещивания проявили инстинкт насиживания. Здесь доминирующими оказались гены петухов, а материнские — рецессивными.
Исследования генетики поведения кур показали значение наследственности также в пищевом и оборонительном поведении. При этом отбор по одному признаку (например, на высокую агрессивность бойцовых петухов) вызывает приспособительную перестройку и других физиологических функций (например, ускорение свертывания крови, что должно способствовать выживанию в «битве»).
Своеобразная передача признаков родителей гибридам была описана и при скрещивании двух видов попугаев: Agapornis rogeicollis и Agapornis fischeri. Так, самки первого вида при постройке гнезда приносят для него веточки, листья и другие материалы, засовывая их между перьями, а самки второго вида — в клюве. Гибриды пытаются совместить оба эти способа, но засовывают веточки очень неловко или не выпускают их из клюва и не могут построить гнездо несмотря на все усилия. В результате у большинства птиц верх берет генетическая программа доставки гнездостроительного материала в клюве, но это достигается обучением в течение 2–3 лет. Вместе с тем гибриды, как и их родители, легко и быстро обучаются всевозможным навыкам поведения.
Наблюдения на мышах показали, что ген, определяющий окраску шерсти, влияет и на поведение. Кроме того, описана связь одних поведенческих признаков с другими. Так, отбор мышей по «эмоциональности», которую оценивали частотой дефекации, привел к выведению двух линий, обозначаемых как реактивные и нереактивные. Оказалось, что в обеих линиях происходило однонаправленное изменение признака двигательной активности, по которому не велось отбора.
В некоторых случаях выявилась связь поведенческих признаков с морфологическими. Так, мыши линии C57BL, отличавшиеся высокой двигательной активностью, выраженностью ориентировочно-исследовательских реакций и малой эмоциональностью, были крупными, с густой шерстью на животе и коротким хвостом, а у мышей линии BALB/c все эти признаки имели противоположную направленность.
Фенотипические программы поведения гибридов отличаются от родительских и тем, что они оказываются более устойчивыми к внешним влияниям. Вместе с тем, когда эти программы складываются с участием влияния раннего опыта, они несут на себе отпечаток условий среды. Для мышей, например, особое значение имеют присутствие человека, шум, освещение, разнообразие окружения и действующих раздражителей, обозначаемые как обогащенная среда. Влияние условий развития мышей в обогащенной среде проявляется в большей степени на гибридах, чем на их родителях из инбредных линий (рис. 60).
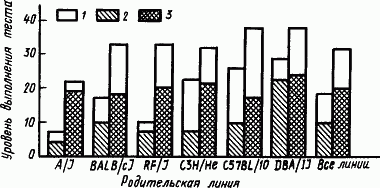
Рис. 60. Средние показатели успешного нахождения пищи мышами инбредных линий и гибридами от их скрещивания, выращенными в условиях стандартной и «обогащенной» среды (по Н. Гендерсону):
1 — эффект обогащенной среды, 2 — инбредная линия, 3 — гибрид
Исследование характеристик памяти на примере сохранения зрительного оборонительного условного рефлекса у крыс показало, что потомки наследуют свойства долговременной, но не краткосрочной памяти родителей.
Генетические программы пищевого поведения рогатого скота формировались в связи с условиями его содержания и кормления. Поэтому, например, у коров обнаруживаются характерные наследуемые породные особенности процесса жвачки. Различия программ пищевого поведения проявляются также в конкурентных отношениях с другими особями при подходе к кормушке. По этим показателям наблюдается прямая зависимость между такой активностью и размерами коров данной породы. Однако сравнительное исследование совместного поведения коров разных пород выявило, что наиболее активны в этом отношении коровы ангусской породы, им уступают шортгорны и на последнем месте оказались герефорды. Внутрипородные отношения перекрываются межпородными. Поэтому крупный герефорд уступает место у кормушки меньшему по размерам ангусу.
Значение наследственности в приручении диких животных наглядно показали наблюдения в питомнике серебристо-черных лисиц, распределение которых по классам прирученности (1 — высший, 2 — средний, 3 — низший) оказалось зависящим от числа ручных предков в их родословной (табл. 13).
Таблица 13. Распределение по классам прирученности, % (по Л.Н. Трут)

К наследуемым признакам относятся и некоторые патологические формы поведения. Так, отбор по чувствительности к звуковому раздражителю позволил вывести линию крыс, в которой 98–99 % особей реагировали на звук судорожными припадками. Характерной особенностью их поведения было двигательное возбуждение после прекращения звукового раздражения. Гибридологический анализ этой линии КМ (Л.В. Крушинский, Л.Н. Молодкина) показал, что такая наследуемая особенность поведения контролируется двумя генами.
Примером другой патологической формы поведения может служить подергивание лапок при выходе из эфирного наркоза у мух Drosophila melanogaster. Особи селекционированной по этому признаку линии были подвергнуты генетическому и нейрофизиологическому исследованию. Оказалось, что такой гиперкинез связан с геном, локализованным в X-хромосоме, и осуществляется нервными клетками грудного ганглия. Продолжение патологических разрядов нейрона даже при перерезке периферических нервов означает, что генерируемая им импульсация зависит от состояния его генетического аппарата.
Наблюдения за пищевым поведением инфузорий показали, что оно определяется их хеморецепцией, на которую оказывает регулирующее влияние аппарат ядра.
Дальнейшее более глубокое изучение механизмов генетического контроля поведения на уровне клетки проводили путем сопоставления мутантных признаков с состоянием физиологических, биохимических и электрофизических характеристик. Так, в опытах со скрещиванием мутантов инфузории Paramecium aurelia установлено, что изменение поведения парамеций путем перестройки движения ресничек при деполяризации или гиперполяризации мембраны контролируется генами определенной локализации. Генетический анализ с помощью спонтанных и индуцированных мутаций и испытания последних на аллельность и рекомбинации позволяют определить локализацию соответствующих генов и выяснить их связь с особенностями поведения.
Из поведения особей, связанного с половым отбором, складывается генотипический состав популяции. В результате миграции индивидов с различным набором генов и предпочтительного выбора самками гетерогенных самцов поддерживается генетический полиморфизм, открывающий широкие возможности возникновения новых признаков, которые закрепляются наследственностью. Таким образом, поведение и его изменчивость становятся одним из механизмов приспособительной эволюции вида.
Наследование типологических особенностей. История приручения диких предков домашних животных дает много поразительных примеров коренного изменения их темперамента. Например, такой контраст наблюдался между поведением коровы, спокойно пережевывающей жвачку, и ее предка — дикого тура, безудержного во взрывах дикой ярости. Подобные метаморфозы поведения достигались настойчивым отбором на протяжении многих веков. Животных отбирали с полезными для их использования типологическими особенностями. Скрещивание особей с наиболее выраженными полезными свойствами усиливало эти признаки. Так, были отобраны спокойные коровы, которые давали себя доить, смелые собаки, которые бросались на опасного хищного зверя, быстрые лошади, чутко реагирующие на малейшее движение всадника. Естественный отбор в природе приспосабливает темперамент животного к условиям его жизни.
Достигнутые в результате отбора признаки типа нервной системы передаются по наследству и проявляются в потомстве как врожденные. Это было показано в результате типологического изучения ряда поколений собак основного генетического стада в лаборатории экспериментальной генетики Института физиологии им. И.П. Павлова АН СССР.
Точные опыты определения типа нервной системы по стандарту, испытаний слюноотделительных рефлексов у родительских пар и потомства показали интересные закономерности наследования материнских и отцовских черт темперамента. Примером может служить следующее наблюдение.
У собаки-самки Пены наблюдали относительную слабость торможения. Она с большим трудом, после длительных применений, выработала дифференцировку, да и то неполную. Пену случили с кобелем Томбушем, имевшим отличную силу как возбудительного, так и тормозного процесса. О силе торможения у Томбуша можно было судить по тому, что ему достаточно было только трех применений дифференцировочного раздражителя, чтобы выработать абсолютную дифференцировку, которая оставалась полной все дальнейшее время работы. У Пены от Томбуша родилось три щенка: Прибой, Познай и Принц. Когда они подросли, типы их нервной системы стали изучать по стандарту типологических испытаний. Это исследование, продолжавшееся более двух лет, показало у всех потомков Пены и Томбуша унаследованную от матери четко выраженную относительную слабость торможения. Все они, как и Пена, с большим трудом вырабатывали дифференцировки, которые так и оставались нестойкими и неполными. Таким образом, типологические признаки одного из родителей оказались доминирующими, а признаки другого — рецессивными.
В другой серии опытов были получены следующие результаты: если оба родителя имели сильные возбудительные процессы, то из 48 потомков 44 были сильного типа и только 4 слабого, но когда один из родителей был сильного типа, а другой — слабого, то потомков сильного типа оказалось меньше (15), а слабого больше (29).
В опытах по изучению степени наследования свойств подвижности оказалось, что у группы мышей с высокой подвижностью нервных процессов 50 % потомков были подвижными и 11 % инертными, а у инертных родителей 50 % инертных и только 6 % подвижных.
Наряду с фактами сохранения типологических признаков родителей в потомстве изучали изменение этих признаков под влиянием условий воспитания. Например, наблюдали, как влияет тренировка нервных процессов мышей на свойства условных рефлексов их потомства. Для этого от пары мышей было получено три помета (24 мышонка). Затем нервная система родительской пары была подвергнута функциональной тренировке. Эта тренировка заключалась в выработке положительных и отрицательных условных рефлексов с последующей неоднократной полной обратной переделкой сигнального значения положительных и отрицательных сигналов. После такой тренировки от этой пары мышей было опять получено три помета (18 мышат).
Все потомство — родившиеся до и после тренировки родителей — прошло типологическое исследование с определением силы, уравновешенности и подвижности их нервных процессов. Оказалось, что мышата, родившиеся после тренировки родителей, имеют несколько большую силу возбудительных процессов и более высокую подвижность. Однако эти эксперименты и выводы из них встретили ряд возражений (Л.В. Крушинский, 1981).
Наследование способности к обучению. Способность обучаться, приспосабливать свое поведение к меняющимся условиям среды обитания на основании индивидуального опыта играет ведущую роль в жизни животных. Поэтому можно ожидать, что эта способность будет совершенствоваться в результате естественного отбора.
Установлено, что различия в скорости образования условных рефлексов выхода из лабиринта у крыс наследуются их потомством. Скрещивание крыс, быстро и медленно обучающихся решению лабиринтной задачи, показало, что в потомстве второго поколения по этому признаку происходило четкое расщепление 3:1. Однако дальнейшие исследования выраженности защитных реакций и эмоциональности у родителей и потомства дали основание предположить, что на скорость образования условного рефлекса влияет степень приуроченности, пугливость животных и выраженность ориентировочно-исследовательского инстинкта.
Во многих работах проводили селекцию по признаку способности к обучению. Так, отбор по лучшим и худшим результатам обучения выходу из 17-тупикового лабиринта, уже начиная с 8-го поколения, дал достоверные различия, а доведенный до 22-го поколения привел к формированию двух стабильных линий — «способных» и «неспособных» крыс. Однако «способные», выведенные экспериментаторами на пищевом подкреплении, в условиях, требующих обучения оборонительным реакциям, оказались хуже успевающими, чем «неспособные». При отборе крыс по скорости выработки условного пищедобывательного рефлекса рефлекс вырабатывался уже в 4-м поколении у крыс «способной» линии за 15 сочетаний, в то время как «неспособные» требовали для этого 23 сочетания.
Обучение является сложным процессом; в него входят восприятие сигнальных раздражений, их анализ, установление связей в механизмах краткосрочной памяти, их консолидация в долгосрочной. Оказалось, что эти компоненты процесса обучения могут наследоваться независимо друг от друга. Особенно значительные различия выявлены при сравнительном изучении краткосрочной и долгосрочной памяти. Так, крысы, быстро обучающиеся, дольше закрепляют памятный след, чем медленно обучающиеся.
Скрещивания кур австралоры и плимутрок показало, что признак величины положительных условных рефлексов наследуется по материнской линии, а величина отрицательных условных рефлексов и подвижность нервных процессов проявляют промежуточный тип наследования. Вопросы наследования способности к обучению у людей рассмотрены в гл. 17.
Генетическая связь условных и безусловных рефлексов. В физиологии высшей нервной деятельности проблема наследования способности к обучению получила своеобразное развитие в изучении взаимоотношений условных и безусловных рефлексов. Условные рефлексы обеспечивают индивидуальную приспособленность, а безусловные воспроизводят в масштабах вида одни и те же акты приспособительного поведения, необходимые для данных условий жизни. Так, у некоторых насекомых можно выработать условные рефлексы питания новыми растительными объектами, а у целых видов насекомых, питавшихся дикими растениями, после посева на целине культурных злаковых был обнаружен на последние безусловный пищевой рефлекс, так что эти насекомые стали злостными вредителями. Таким образом, возникает вопрос о возможности некоторой генетической связи между безусловными и условными рефлексами.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.6. Типы высшей нервной деятельности
6.6. Типы высшей нервной деятельности Условно-рефлекторая деятельность зависит от индивидуальных свойств нервной системы. Индивидуальные свойства нервной системы обусловлены наследственными особенностями индивидуума и его жизненным опытом. Совокупность этих свойств
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ С незапамятных времен внимание человека привлекали проявления психической жизни. Откуда берутся мысли? Как понимать сознание, вмещающее весь мир и управляющее нашими поступками? Что
Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Вся сложная и разнообразная деятельность высших отделов нервной системы построена на работе двух основных нервных процессов — возбуждения и торможения. Протекая в подвижных пространственных и временных
Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА Гибкий и подвижный механизм высшей нервной деятельности обнаруживает громадные приспособительные возможности нормального функционирования при самых разных состояниях организма.Знание
ГЛАВА 3. УПРАВЛЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТЬЮ (ПОВЕДЕНИЕМ)
ГЛАВА 3. УПРАВЛЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТЬЮ (ПОВЕДЕНИЕМ) Поведение собаки, т.е. сложная совокупность ее взаимодействий с окружающей средой, привычно рассматривается человеком с позиций собственного душевного мира. Такой подход к психике животных хорошо объясним и,
3.10. НАРУШЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
3.10. НАРУШЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Сочетание нейрохирургических вмешательств в определенные отделы мозга и условно-рефлекторного метода привело к пониманию многих сторон высшей нервной деятельности. Анализ ее нарушений расширил представления о работе мозга в
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 20 ]
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 20 ] Второй раз я имею высокую честь и глубокое удовлетворение принимать участие в торжестве открытия деятельности чрезвычайных русских обществ, народившиеся здесь, в Москве, и свидетельствующих о том, как
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 25 ]
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 25 ] Прежде всего я считаю своим долгом благодарить Философское общество, что оно в лице своего председателя изъявило готовность выслушать мое сообщение. Мне трудно было сообразить, насколько это
ЭКСПЕРИМЕНТАЛЬНАЯ ПАТОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ [ 51 ]
ЭКСПЕРИМЕНТАЛЬНАЯ ПАТОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ[ 51 ] Несколько вступительных слов о сложной судьбе нашей работы по физиологии и патологии высшей нервной деятельности, принимая, что прилагательные «высшая нервная» отвечают прилагательному
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1 Образы, картины поведения как нас самих, так и близких к нам высших животных, с которыми мы находимся в постоянных жизненных отношениях (как, например, собак), представляют огромное разнообразие, прямо
ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ В СВЯЗИ С НЕВРОЗАМИ И ПСИХОЗАМИ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ НЕВРОТИЧЕСКИХ И ПСИХОТИЧЕСКИХ СИМПТОМОВ [ 52 ]
ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ В СВЯЗИ С НЕВРОЗАМИ И ПСИХОЗАМИ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ НЕВРОТИЧЕСКИХ И ПСИХОТИЧЕСКИХ СИМПТОМОВ[ 52 ] Из огромного материала, относящегося к изучению высшей нервной деятельности у собак по методу условных рефлексов, я остановлюсь
6. Типы высшей нервной деятельности
6. Типы высшей нервной деятельности На основании изучения условно-рефлекторной деятельности огромного числа собак академик И. П. Павлов создал свое учение о типах высшей нервной деятельности. Все многообразные особенности условно-рефлекторной деятельности собак
7. Патология высшей нервной деятельности
7. Патология высшей нервной деятельности Одной из сторон работ академика И. П. Павлова, имеющей большое значение для служебного собаководства, является развитие им учения о патологии нервной деятельности.Высшая нервная деятельность собаки может быть сорвана
Некоторые актуальные вопросы генетики поведения и высшей нервной деятельности[50]
Некоторые актуальные вопросы генетики поведения и высшей нервной деятельности[50] Исследования по генетике поведения начались почти с самого начала нынешнего столетия, когда законы Менделя стали достоянием широких кругов биологов.При этом выяснилось, что признаки