Глава 10 ПРИМИТИВНЫЕ ФОРМЫ ВРЕМЕННЫХ СВЯЗЕЙ
Глава 10
ПРИМИТИВНЫЕ ФОРМЫ ВРЕМЕННЫХ СВЯЗЕЙ
Приспособительная изменчивость поведения свойственна всем живым существам. Уже простейшие могут образовывать своеобразные временные внутриклеточные связи. С появлением у кишечнополостных нервной системы (органа раздражимости многоклеточных организмов) эту функцию вначале берут на себя временные связи диффузной нервной системы. Затем нервная система централизуется, и, начиная с низших червей, индивидуальные приспособительные реакции животных осуществляются с помощью временных рефлекторных связей. Их развитие и дает начало истории условного рефлекса как исходного механизма формирования все более сложных форм высшей нервной деятельности.
Временные связи простейших
Как справедливо замечено, «в термине „простейшие“ больше иронии, чем правды». Действительно, микроскопическая амеба своей протоплазмой осуществляет сложнейшие функции раздражимости, передвижения, пищеварения и т.п., для обеспечения которых у высших животных работают целые органы и системы. Среди этих функций обращает на себя внимание отчетливо выраженная у некоторых из простейших способность к образованию внутриклеточных временных связей.
Амеба. Медленными тягучими движениями перемещается амеба, переливаясь протоплазмой тела в выпячивающиеся ложноножки. Однако, несмотря на крайнюю простоту своей моторики, она способна к приспособительной изменчивости поведения.
Возможность выработки новых форм реакций у амеб изучали в опытах с повторным действием сильного светового раздражения. Если на пути ползущей амебы поставить узкий луч от мощного источника света, то, как только ее ложноножка попадает под яркое освещение, амеба останавливается. Затем она начинает выпускать ложноножки, более или менее отклоняющиеся от прежнего направления, вновь втягивая их каждый раз, как только они входят в соприкосновение со световым лучом. Наконец, после ряда «проб» амеба резко изменяет направление своего движения и, выпустив ложноножку назад, уходит от световой преграды. Оказалось, что многократное повторение таких опытов на одной и той же особи приводит к значительному уменьшению числа «пробных» вытягиваний ложноножек до ухода назад. Например, в одном из опытов при первых трех испытаниях амебы совершали «пробные» вытягивания ложноножек 20 раз, при вторых испытаниях — 7, а при шестых — только 4 раза.
Стилонихия. На стеблях и листьях водных растений на дне пресноводных водоемов обитает брюхоресничная инфузория стилонихия. Органами передвижения ей служат склеенные между собой реснички — цирры. На них стилонихия поднимает свое тело, и переступая циррами, как ножками, ходит и даже подпрыгивает в поисках добычи.
Оказывается, у стилонихий легко вырабатывается временная связь между освещением и неблагоприятными условиями передвижения. Для этого следует пустить стилонихию в сосуд с наполовину шероховатым, а наполовину гладким дном и осветить шероховатую половину, а гладкую затемнить. Через некоторое время свет перестает быть для стилонихий безразличным и приобретает сигнальное значение. Свет становится сигналом «плохой дороги». Теперь он вызывает реакцию ухода в другое место. Это легко обнаружить, пересадив стилонихий в другой сосуд с наполовину освещенным, но на всем протяжении одинаково гладким дном. Как только стилонихия, находящаяся на темной половине, доходит до границы со светлой, она немедленно поворачивает обратно.
Физиологический механизм этих связей, видимо, состоит в повышении или понижении раздражимости, возможно, суммационной природы. Однако эти связи чрезвычайно слабые и рыхлые. Они разрушаются немедленно после того, как перестали действовать создавшие их раздражители. Достаточно нескольких минут, чтобы раздражимость начала возвращаться к исходной величине, и приобретенный новый способ реагирования был уже частично утрачен.
Туфельки. Почти в каждой капле стоячей воды можно увидеть под микроскопом быстро плавающие в разных направлениях или остановившиеся у комочка водорослей равноресничные инфузории из рода туфелек. Наиболее крупная из них — хвостатая туфелька, ведет себя очень активно, находится почти все время в движении и является удобным объектом для изучения приспособительной изменчивости поведения.
Туфелька обычно заглатывает все встречающиеся ей мелкие частицы. Однако она может выработать различение съедобных частиц от несъедобных. Это видно из следующего опыта.
В воду культуры туфелек вносят взвесь тонко растертого порошка алюминия, и время от времени берут туфелек из этой культуры для просмотра пищеварительных вакуолей. Вначале туфельки заглатывают алюминиевые частички. Со временем заглотанных частичек становится все меньше, и примерно через 20 ч туфельки перестают заглатывать алюминий, хотя органические частицы, например белка, по-прежнему энергично поедаются ими.
Приобретенные формы поведения сохраняются лишь до первого деления инфузорий. При перестройке клеточных структур во время акта размножения временные связи бесследно разрушаются. Дочерние туфельки должны заново приобретать свой жизненный опыт.
Подвижные инфузории легко вырабатывают временные связи на ориентиры своего обычного пути следования. Так, плавая в треугольном сосуде, вдоль его стенок, и многократно поворачивая в конце каждой грани на 120°, туфелька «привыкает» к такому повороту. Теперь ее можно выпустить в круглый сосуд и тем не менее она будет выписывать в нем треугольники. В то же время туфелька, выдержанная в четырехугольном сосуде, будет и в круглом сосуде при каждом столкновении со стенкой поворачивать на 90° и ее путь будет иметь форму квадрата (рис. 71).

Рис. 71. Путь туфельки после содержания в четырехугольном (А) и треугольном (Б) сосудах (по Брамштедту)
Временные связи простейших помогают им осваивать новые способы передвижения.
Если туфельку пустить в тонкий запаянный на конце капилляр, просвет которого меньше длины ее тела, она не сможет повернуться и будет вынуждена плыть все время вперед. Достигнув конца капилляра, туфелька окажется в тупике. Отступая и вновь устремляясь вперед, изгибаясь то в одну, то в другую сторону, она будет долго биться, пока, наконец, не найдет способа повернуться и выйти из узкого места. В первый раз для этого потребуется много времени, потом все меньше и меньше. После 15–20 таких «упражнений» туфелька, дойдя до конца капилляра, привычным движением сразу разворачивается и плывет обратно.
Новый прием передвижения в узких ходах осваивается туфелькой за несколько минут. Это естественный приспособительный механизм. Образование временных связей подобного рода несомненно происходит в природных условиях, позволяя инфузориям приспосабливаться к разнообразным условиям добывания пищи, например в узких складках и щелях гниющих растений.
У инфузорий удалось также выработать положительную реакцию на свет после того, как он многократно сочетался с тепловым воздействием. Однако в аналогичных опытах с сочетанием света и электрического раздражения показано, что здесь, возможно, играет существенную роль не свет, а место сосуда (капилляр), в котором производилось электрическое раздражение. Оказалось, что инфузории, дойдя до этого места, поворачивали, даже если свет не включался. Высказано предположение, что речь идет о реакции инфузории на какие-то вещества, выделявшиеся ею в ответ на действие электрического тока и накопившиеся в том месте, где инфузорию подвергали раздражению. Однако в аналогичных опытах, поставленных в широкой камере, туфельки также вырабатывали подобную реакцию, хотя трудно допустить, что они создавали зону выделений на столь большом протяжении. Вместе с тем даже в отсутствие света туфелька, достигнув этой условной линии, поворачивалась и уходила обратно. Возможно, выработка такой реакции, как и описанной выше реакции движения по определенному пути, имеет в своей основе сигналы от аппарата передвижения.
Если «засасывать» туфельку в капилляр, конец которого погрузить в сосуд с питательной средой, то с каждым разом она выплывает из капилляра все скорее. Однако эти опыты получили также иное объяснение, чем образование временных связей. После 10 сочетаний подпороговой вибрации (400–500 Гц) с ударом электрического тока, останавливающего движения туфелек, вибрация сама стала останавливать их движения. Дифференцировка к вибрациям 300 и 30 Гц вырабатывалась за несколько применений.
Сувойки. Весьма интересна возможность выработки своеобразных временных связей у колониальных форм круглоресничных инфузорий. Сочетая освещение красным или синим светом с прикосновением к сувойкам, получали после 140–160 сочетаний сокращение всех особей колонии на одно только применение света. Пересадка таких сувоек на другую колонию сообщает последней свойства защитной реакции на свет, сохраняющиеся и после удаления трансплантата. Не исключена возможность, что временные связи здесь имеют еще неисследованную гуморальную природу.
Временные нервные связи кишечнополостных, иглокожих, червей и моллюсков
Возникнув как орган раздражимости многоклеточных животных, нервная система берет на себя руководство всеми адаптивными реакциями организма. В частности, образование временных связей становится ее специальной функцией.
Кишечнополостные. Нервная система кишечнополостных имеет сетевидное строение. Прикрепленные гидры и полипы, ограниченные в своей моторике и рецепции, обладают сравнительно малыми возможностями для образования временных связей в своей диффузной нервной системе. Такие связи вырабатываются главным образом между сигналами, которые получают щупальца, и поисковыми движениями захватывающих органов. Это можно наблюдать в следующем опыте (рис. 72).
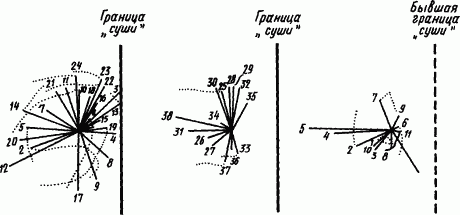
Рис. 72. Выработка временных нервных связей, изменяющих направление поисковых движений щупалец пресноводной гидры (по А.А. Зубкову, Г.Г. Поликарпову), А — вытягивание щупалец в течение первых 67 мин после прикрепления на расстоянии 5 мм от края водоема; Б — то же, в течение следующих 93 мин после отдаления границы между водой и «сушей» до 20 мм; В — то же, спустя 120 мин после удаления границы:
цифры отражают длину и продолжительность вытягивания в условных единицах, пунктиром отмечены отклонения от первоначального направления вытягивания
Если в часовое стекло налить воду и поместить туда пресноводную коричневую гидру, постоянную обитательницу почти каждого водоема, то, оправившись от сотрясений и прикрепившись подошвой ко дну, гидра вскоре выпустит щупальца и начнет свои поисковые движения, простирая щупальца во все стороны. Наклоняя часовое стекло, можно добиться, чтобы гидра оказалась почти на границе воды с воздухом и имела возможность вытягивать свои щупальца за добычей только в одну сторону. После того как она пробудет в таком положении 2–3 ч, можно вернуть стекло в прежнее положение, и, хотя гидру вновь окружает со всех сторон вода, поисковые движения она будет совершать только в одну сторону, а именно в ту, куда она вытягивала свои щупальца, находясь в условиях ограничения движений. С возвращением прежних условий такая приобретенная реакция постепенно утрачивается.
У гидр оказалось возможным также выработать после 100–300 сочетаний реакцию втягивания щупалец на легкое вибрационное раздражение, которое ранее не вызывало такого защитного движения. Для этого колебания воды язычком электромагнитного вибратора сочетали с сильным механическим раздражением тела гидры пузырьками воздуха из подведенной к месту ее прикрепления трубки. Однако достаточно было сделать небольшой перерыв в опытах и выработанная реакция исчезала. Эти и другие примеры показывают свойства суммационных реакций, которые требуют непрерывного возобновления и, с трудом образуясь, быстро разрушаются.
Отрицательное научение — реакция привыкания у гидры оставляет некоторый памятный след, который проявляется в ускорении повторного привыкания, вызванного через 1 ч, однако через 24 ч этот след исчезает.
Актинии питаются, щупальцами захватывая рыб и других небольших водных животных. Если поднести к щупальцу актинии кусочек мяса, то она его схватит и с помощью других щупалец втолкнет через ротовое отверстие в гастральную полость, где мясо будет переварено. Если же дать ей что-либо несъедобное (например, комок бумаги), то актиния сначала его тоже схватит, но потом выбросит прочь. Повторяя подачу несъедобной бумаги, можно добиться угнетения пищевой реакции схватывания. После того как эта реакция полностью исчезнет в результате многократных раздражений одного из щупалец, ее можно вызвать, поднося бумажный комок к другому щупальцу.
Этот факт дает основание предполагать, что временные связи нервной системы, еще не достигшей централизации, имеют местный ограниченный характер. Что касается физиологического механизма этих связей, то в его основе, видимо, также лежат временные изменения возбудимости, создаваемые взаимодействием раздражителей. Значение суммационных явлений в образовании таких связей наглядно показывают опыты по изменению интервалов между очередными сочетаниями света с электрическим раздражением, вызывающим сокращения тела гидры. Когда интервал между сочетаниями составлял 15–30 с, то один свет при своем изолированном применении мог вызвать двигательную реакцию, но если интервалы между раздражениями увеличивались до 3–5 мин, то свет терял свое действие.
Иглокожие. Начальная централизация нервной системы у иглокожих позволяет ожидать возникновения у них зачаточных условных рефлексов. Однако образ жизни этих медлительных обитателей морского дна, по-видимому, не благоприятствовал развитию такой возможности. В их приспособительном поведении преобладают реакции типа «привыкания».
Морской еж очень чувствителен к световым сигналам. Падающая на ежа тень проплывающей рыбы служит сигналом опасности и вызывает у него оборонительную реакцию — иглы поднимаются и поворачиваются в сторону рыбы. Если многократное затенение не сопровождается вредящими раздражениями, то оборонительная реакция начинает задерживаться. Однако как только сигнальное значение тени будет подкреплено, например за тенью последует прикосновение к коже животного, оборонительная реакция восстановится.
Существуют противоречивые наблюдения о способности морских звезд к выработке условных рефлексов на кожно-мышечные сигналы. Так, при повторных одеваниях резиновой трубки на луч змеехвостки из семейства Ophiomixidae последняя вырабатывает реакции быстрого освобождения от трубки новыми все более удачными приемами. Однако проверка этих наблюдений показала, что время, необходимое для освобождения луча от трубки, в многократных повторных опытах практически не уменьшается.
Черви. У червей возникает хорошо централизованная узловая нервная система. Они способны образовывать временные связи, которые уже можно обозначить как примитивные условные рефлексы, хотя физиологический механизм последних весьма далек от такового у высших животных.
Если периодически направлять на планарию пучок света и каждый такой 3-секундный засвет сопровождать на последней секунде ударом электрического тока, то после ряда сочетаний один только световой сигнал вызовет условную оборонительную реакцию продольного сокращения тела.
Сообщения о том, что другие планарии, поедая «обученных», приобретают их навыки, было началом исследований так называемого переноса памяти, о котором шла речь в гл. 2 (Дж, Мак-Коннелл, 1971).
Выработанная у морских планарий условная оборонительная реакция сохранялась 10–15 ч. Она образовалась и у пресноводных планарий, но лишь после многих сочетаний и была непрочной. Переделка реакции ухода в темноту в лабиринте, где темный проход находился под током, привела к тому, что через 5 дней почти 3/4 планарий ползли в светлый проход.
Если планарий, у которых выработан условный оборонительный рефлекс, разрезать пополам, то не только головная, но и хвостовая половина способны к воспроизведению условной реакции. По регенерации из каждой половины планарий целого организма исследовали свойства сохранения памятного следа. Исследование проводили в лабиринте с темным выходом, находящимся под током, в котором у планарий до разрезания вырабатывали условный рефлекс избегания темноты. Он сохранился у планарий, регенерировавших из головной половины, и в меньшей степени — из хвостовой.
Так как опыты не только с поеданием обученных планарий, но и введением их гомогенатов были связаны с представлением о химическом носителе памяти (Дж. МакКоннелл, Дж. Шелби, 1970), то изучали действие таких гомогенатов на планарий, регенерировавших из обеих половин тела. Оказалось, что условная реакция лучше сохранилась у планарий, регенерировавших из головной части. Отмечена также высокая чувствительность к гомогенату и нервных клеток вновь образующейся головной части при сохранении хвостовой. Условные реакции планарий по их свойствам могут быть отнесены к суммационным рефлексам.
В природной обстановке очень многие черви чутко реагируют на изменение освещенности. Например, один из многощетинковых морских кольчецов живет в трубке, которую строит из песчинок и других мелких частиц, склеивая их слизью кожных желез. Обычно передняя часть его тела находится снаружи. Достаточно, чтобы на червя упала легкая тень, сигнализирующая об опасности, и он немедленно прячется в трубку. Многократное появление тени без каких-либо существенных последствий для червя приводит к торможению защитной реакции, которая постепенно угасает. Торможение, вырабатываемое в узлах нервной цепочки, оказывается еще слабым и недолгим, но рассеивается без всяких вмешательств в течение нескольких минут и сразу исчезает даже при однократном подкреплении теневого сигнала механическим раздражением.
При соответствующих условиях тень может сыграть и роль сигнала пищи. Так, после многократных сочетаний кормления червей нереид с появлением тени они стали высовывать головы из трубок при одном только затенении их убежища. Другие опыты показали, что если этих червей кормить только при освещении, то они могут с равным успехом связать с пищей свет.
Часто можно увидеть торчащие в сырой земле травинки, соломинки или обрывки листьев. Если вытащить такой листок, то станет видно, что он застрял у входа в норку дождевого червя. Если лист, который дождевой червь тянет в свою норку, оказывается слишком большим и застревает у входа, червь после нескольких безуспешных попыток оставляет его на месте, делает другой выход и, отправляясь за новыми листьями, проползает мимо этого листа, как будто его не существует. Такое выработанное торможение пищевой реакции дождевого червя оказывается более стойким, чем описанное выше торможение оборонительной реакции у морских кольчецов.
Свойства условных рефлексов червей удобно изучать в специальном аппарате Р. Иеркса для их «дрессировки».
Червя выпускают в коридор, который заканчивается развилкой. Если червь повернет направо, он попадет в ящик с влажной землей — свое жилище, богатое кормом. Если же он повернет налево, то натолкнется на оголенные провода, находящиеся под напряжением и, замкнув своим телом цепь тока, получит сильный электрический удар. Вначале червь поворачивал вправо или влево без специального выбора, случайно, и поэтому число поворотов в обе стороны оказывалось в среднем примерно одинаковым. Однако после 30–50 упражнений он стал заметно избегать поворота, приводящего к току, а после 150–200 проб поворачивал только на безопасный путь, в ящик с землей. Так, образовался условный рефлекс на осязательно-мышечные сигналы с двойным подкреплением: оборонительной реакцией, вызванной электрическим ударом после поворота в левое ответвление коридора, и пищевой при попадании через правое ответвление в ящик с землей. Рефлекс оказался довольно прочным. Он не угасал даже после нескольких испытаний без подкреплений.
Выработанный условный рефлекс можно переделать, если перенести провода в правую ветвь коридора, а из левого сделать выход в ящик с землей. Тогда червь начинает поворачивать не вправо, а влево.
Большой интерес имеют опыты, проведенные с червями, которым была сделана операция удаления головных узлов нервной цепочки. Оказалось, что такая операция не препятствовала выработке условного рефлекса поворота в заданном направлении, а если рефлекс был уже выработан, то не уничтожала его. Оказалось также, что если «обученного» червя разрезать пополам, то не только передняя, но и задняя половина тела сохраняла выработанный навык поворота на развилке. Отсюда следует, что временные связи у него образовывались не только в головных, но и во всех других узлах нервной цепочки. Это очень вероятно, так как осязательно-мышечные сигналы, на которые вырабатывался данный рефлекс, приходили последовательно от каждого сегмента тела в соответствующий узел. Здесь производился их анализ, и осуществлялось замыкание на нервный механизм соответствующего этапа движения.
Моллюски. Почти у всех моллюсков центральная нервная система сосредоточивается в виде крупных надглоточных и подглоточных узлов. Эти узлы координируют движения и организуют целостное поведение по сигналам дистантных рецепторов. В них, видимо, происходит замыкание временных связей условных рефлексов. Большинство улиток ведет малоподвижный образ жизни. Одни из них отфильтровывают из протекающей воды свой корм, другие — кормятся на растениях, третьи — плотоядные, прикрепившись к своей жертве, постепенно ее выедают. В связи с этим условные рефлексы на прием пищи у них развиты слабо. Чтобы выработать у пресноводной улитки физы реакцию открывания рта на сигнальное прикосновение к ноге, за которым следовало кормление, потребовалось около 250 упражнений. Более развиты у этих моллюсков оборонительные рефлексы. Так, для многих наземных улиток приближение опасности, например птицы, которая может их схватить, сигнализируется тенью, внезапно падающей на глазки щупалец. Улитка немедленно останавливается и втягивает щупальца. Этот натуральный защитный рефлекс можно полностью угасить, повторяя теневой сигнал без подкрепления. Разные виды улиток проявляют различные свойства выработки такого торможения.
При выработке у виноградной улитки оборонительной условной реакции наступали характерные изменения импульсации командных нейронов, защитные реакции дыхания. В других опытах у улитки вырабатывали условную оборонительную реакцию на время путем периодической микростимуляции отдельного командного нейрона. У некоторых морских моллюсков можно было выработать оборонительную реакцию на определенный вид пищи, если он сопровождался болевым раздражением. При этом предъявление пищи угнетало импульсацию командного нейрона пищевой реакции.
У приморского гребешка быстро (хотя и непрочно) вырабатывался условный оборонительный рефлекс на свет, вслед за которым в аквариум опускали хищную морскую звезду.
Среди вырабатываемых моллюсками реакций большое практическое значение имеет «приучение» устриц к длительному плотному закрыванию створок раковины, что делает этих съедобных моллюсков пригодными для дальней транспортировки в живом виде. Для этого прибрежные бассейны (откормочные парки) с устрицами начинают временно лишать воды. В первые дни они осушаются на полчаса в сутки, затем на более длительный срок и, наконец, на всю ночь. Каждый раз, когда устрица, оставаясь на воздухе, пробует открыть створки, она теряет свою воду. Вскоре вырабатывается прочный защитный рефлекс. Обсушивание поверхности становится сигналом удерживания раковины в закрытом положении. Таких устриц можно перевозить и сохранять живыми в течение нескольких дней.
Выдающееся положение среди других моллюсков занимают головоногие, быстро вырабатывающие новые поведенческие навыки: небольшие спруты охотятся за раками-отшельниками, но когда этим спрутам подбросили раков, несущих морские анемоны, то ужаленные ими спруты перестали нападать на раков-отшельников.
Очень активный образ жизни ведет осьминог — хищник морей. Его головной мозг обнаруживает четкую локализацию центров выполнения различных рефлекторных актов (рис. 73). Охота за рыбами и крабами вырабатывает у осьминога разнообразные натуральные рефлексы нападения на движущиеся объекты. Однако достаточно ему 10–15 раз подряд схватить протаскиваемый перед ним несъедобный предмет, как осьминог перестает на него реагировать. Это торможение обнаруживает все свойства условного и должно быть отнесено к категории угасательного. Оно оказывается довольно стойким и может пройти более 1 ч, пока осьминог вновь схватит движущуюся приманку.
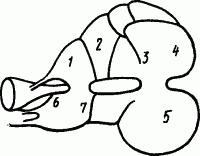
Рис. 73. Расположение центров рефлекторных движений в нервных узлах головоногого (по И. Икскюлю): 1 — смыкание челюстей, 2 — еда, 3 — плавание, 4 — ползание, 5 — дыхание, 6 — прикрепление присосок, 7 — освобождение присосок
Оборонительный условный рефлекс на световой сигнал, сопровождаемый уколом, осьминог сохраняет без подкрепления более 3 месяцев. По своей условно-рефлекторной деятельности осьминог превосходит других головоногих, кальмаров и каракатиц, что можно объяснить развитием его мозга в результате использования щупалец как своеобразных «рук», выполняющих разные функции.
Осьминоги легко вырабатывают условные зрительные пищевые рефлексы, иногда весьма сложные. Например, обнаружив краба в стеклянной банке, осьминог вначале бьет щупальцами о стекло, долбит его клювом, пока не найдет отверстие в банке. На следующий день осьминог, увидев краба в банке, не предпринимает бесплодных попыток пробиться к нему через стекло, а сразу вытаскивает краба через отверстие.
У осьминога можно выработать различение показываемых фигур по форме, подкрепляя одни из них и не подкрепляя другие кормлением, а также более сложные формы высшей нервной деятельности, как, например, условный тормоз. Однако об их способности различать цвета имеются противоречивые данные. Осьминоги успешно вырабатывали на пищевом подкреплении условную реакцию вытягивания одного из щупалец в трубу, опускаемую в воду. Очень интересна способность осьминогов строить себе из камня убежища, материал для которых они тщательно выбирают и в случае необходимости перетаскивают иногда на значительные расстояния. Конечно, в сложном поведении головоногих проявляются не какие-то «идеи» или «представления», как предполагали некоторые исследователи, а способность их мозга к сравнительно тонким зрительным дифференцировкам, выработке условных тормозов и специфической манипуляционной деятельности.
Условные рефлексы ракообразных и паукообразных
Уже у низших членистоногих проявляются некоторые общие тенденции эволюции этого типа животных, в частности, чрезвычайная видовая специализация форм приспособительного поведения.
Дафнии. Дафнии составляют основную массу планктона пресноводных водоемов. При недостатке корма или других неблагоприятных обстоятельствах они устремляются к свету, который сигнализирует о богатых пищей и более теплых поверхностных слоях воды. Однако при изменении условий жизни может измениться и сигнальное значение света.
Такое изменение, например, наблюдали в эксперименте при сочетании света с холодом, а тепла с темнотой.
Для этого сосуд с одной группой дафний при ярком освещении охлаждали до 2–4 °С и оставляли при низкой температуре в течение 4 ч. Затем его затемняли и нагревали до температуры 15–20 °С, оптимальной для дафний. У другой группы дафний вырабатывали обратные отношения, т.е. свет сочетался с теплом, а темнота с холодом. Соответствующие условные рефлексы быстро образовывались: дафнии первой группы шли в темноту, дафнии второй группы — к свету.
Изменить сигнальное значение света для дафний можно также, создавая иные условия питания.
Для этого помещали дафний в хорошо освещенный сосуд с чистой водой без корма. К вечеру их переносили в сосуд с водой, богатой инфузориями, закрывали светонепроницаемым колпаком и оставляли на всю ночь кормиться в темноте. Утром после ополаскивания дафний опять переносили в первый сосуд голодать весь день на свету и т.д. В результате у дафний выработался прочный условный пищевой рефлекс на темноту.
Ниже приведены данные по выработке условного пищевого рефлекса на темноту у дафний (по А.Б. Когану и А.П. Семеновых).
Число дней кормления в темноте и голодания на свету | Процент положительных реакций на темноту
0 | 0
7 | 32
14 | 57
21 | 89
Раки. Речные раки собираются к месту, где они несколько раз находили пищу. Этой способностью раков пользуются при их ловле, известна защитная реакция раков-отшельников, скрывающих свое мягкое брюшко в пустые раковины. Если ракам-отшельникам дать несколько раковин, отверстия которых забиты пробками, то после безуспешных попыток ввести брюшко в отверстия раковин раки прекращают это занятие. Такое торможение защитной реакции вырабатывается медленно. Пытаясь открыть раковины, раки упорно действуют клешнями в течение нескольких суток. Но когда торможение выработалось, оно оказывается весьма прочным. Даже спустя нескольких дней ракам предлагали те же раковины, но заклеенные тонкой бумагой вместо пробок, и бумага оставалась целой, отшельники даже не прикасались к объектам своей недавней энергичной деятельности. Однако если предложить им другие раковины, то раки немедленно их схватывают и начинают обследовать. Видимо, здесь происходит дифференцирование формы предмета.
Десятиногих раков испытывали в «лабиринте» (рис. 74, А). Вначале число выходов в аквариум (1) и заходов в тупик (2) было примерно одинаковым, лишь постепенно раки стали правильно выбирать дорогу. Рис. 74, Б показывает «кривую обучения» раков. Даже после двух недель перерыва в работе условный рефлекс хорошо сохранился. Этот факт в соответствии с приведенными выше наблюдениями за поведением раков свидетельствует о значительной стойкости вырабатываемых временных связей.
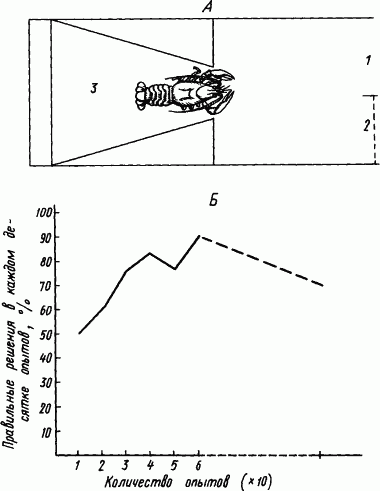
Рис. 74. Опыты «обучения» раков в простом лабиринте (по В.М. Боровскому). А — схема лабиринта; Б — кривая «обучения» рака в этом лабиринте:
1 — свободный вход в аквариум, 2 — стеклянная перегородка, закрывающая выход, 3 — выпускное отделение, прерывистая часть абсциссы означает 2-недельный перерыв
Когда речных раков стали кормить только в освещенной части аквариума, то у них за 4–12 сочетаний образовался и за 10–20 сочетаний укрепился условный пищевой рефлекс на освещение. Такой рефлекс удалось угасить за 6–17 применений светового сигнала без подкармливания, но на следующий день рефлекс восстанавливался.
Получены сведения о способности краба, где бы он ни находился, при испуге бежать по прямой линии к своей норе. Показано, что этот краб может использовать для ориентации сигналы поляризованного света. Однако скальный омар, покидая свое убежище для ночной кормежки, возвращается в него, используя, по-видимому, местные ориентиры. Вместе с тем некоторые наблюдения указывают на врожденную способность членистоногих к ориентации по неизвестным пока признакам. Так, когда особям береговой блохи, рожденным и выросшим в лаборатории, была предоставлена свобода передвижения, то они устремились в направлении берега, где обитали их родители.
Пауки. Условия охоты пауков сделали жужжание насекомых натуральными сигналами их охотничьей пищевой реакции. При этом паук живо реагирует на сотрясение паутины именно с той частотой, с которой муха бьет крыльями, пытаясь освободиться из плена. По-видимому, рассказы о «музыкальности» домового паука имеют известное основание. Некоторые тоны скрипки, близкие к мушиному жужжанию, привлекают паука так сильно, что он спускается как можно ближе к инструменту. Если подобрать камертон, имеющий сходную тональность, то, услышав его звучание, паук немедленно выглянет из своего убежища. Однако после нескольких таких применений камертона вырабатывается торможение реакции, не получающей пищевое подкрепление, звук теряет свое сигнальное значение.
Живущие в норах тарантулы подстерегают добычу у входа в нору. Они выскакивают и схватывают проходящих жуков, бабочек, стрекоз и других насекомых, но не реагируют на неподвижных.
Пищевой рефлекс паука очень силен и может быть использован для подавления защитного. Описывают, что даже свирепого калаб<...>ийского тарантула можно приучить к жизни в неволе в бумажной трубке, откуда он выходит и берет живых мух из рук человека.
Если предложить пауку-прыгуну муху, смазанную терпентином, то он ее не возьмет. Пищевой рефлекс будет заторможен. После того как пауку трижды давали таких мух, он переставал брать и обыкновенных мух, годных ему для еды. Это наблюдение, так же как и предыдущее, считают проявлением малой способности нервной системы паука к выработке дифференцировок. Однако когда паукам предлагали мух, пропитанных сахаром или хинином при звуках метронома 262 или 523 колебаний/с соответственно, то уже после 15 проб они переставали хватать мух при дифференцировочном звуке.
В пространственной ориентации пауков большую роль играет зрение. Так, паук при вибрации паутины от попавшего в нее насекомого бросается на него и, схватив, возвращается в свое убежище. При этом он ориентируется главным образом по направлению освещения. Если после того, как он схватил добычу, переместить источник света, то паук окажется дезориентированным и не сумеет сразу вернуться в убежище (рис. 75). По-видимому, обучение ориентации по направлению освещения происходит быстро, так как если источник света переместить даже незадолго до того, как паук бросится на добычу, то он уверенно возвращается в убежище.

Рис. 75. Дезориентация паука Agelena изменением положения источника света (по М. Бартельсу). А — путь паука в гнездо при обычном положении источника света; Б — путь паука при перемещении источника света на другую сторону
Действие некоторых фармакологических веществ на паука исследовали, пропитывая ими предлагаемых ему мух, оценивая результат по его поведению при выплетании паутины. Получив хлоралгидрат, паук засыпал, прервав выплетание. Кофеин вызвал возбужденную деятельность по беспорядочному сплетению паутинных нитей. Неожиданный эффект действия лизергиновой кислоты состоял в том, что паук выплетал наиболее точно рассчитанную и безукоризненно выполненную паутину (Н. Тинберген, 1978).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
11.2. Почему «процветают» примитивные паразиты
11.2. Почему «процветают» примитивные паразиты Одним из главных материалистических достижений дарвинизма явился отказ от признания ламарковского внутреннего стремления к совершенству как одной из причин эволюции. (В гл. 1 мы говорили об отношении Ч. Дарвина к прогрессу,
Эти «примитивные» губки…
Эти «примитивные» губки… Идея «клеточного государства», в общем, довольно спорная, если речь идет о принципах строения высших животных, не покажется, вероятно, столь уж абсурдной, если мы спустимся на несколько ступенек вниз по эволюционной лестнице. Давайте задержимся
Глава вторая Основные формы животных
Глава вторая Основные формы животных Мы видели, что в природе все беспрестанно изменяется, перестраивается, совершенствуется, но вместе с тем каждому известно, что основная форма тела[16] животных сохраняется неизменной, не только при самых разнообразных внешних
6.1. Роль обратных временных связей в нервном механизме «психологической защиты»
6.1. Роль обратных временных связей в нервном механизме «психологической защиты» Одна из форм психологической защиты выражается в повышении порога осознания эмоционально неприятных для субъекта явлений внешней среды, которое охраняет его сознание от психологически
Эволюция формы
Эволюция формы Морфология реальных организмов или их частей может достигать очень высокой сложности, а поэтому производить количественные определения скоростей изменения формы труднее, чем определения скоростей таксономического изменения или изменений размеров. Это
3.1. Проблема формы
3.1. Проблема формы Не очевидно, что форма вообще представляет какую-то проблему. Окружающий нас мир полон форм, мы узнаем их в каждом акте восприятия. Но легко забываем, что существует глубокая пропасть между этим аспектом нашего опыта, который мы воспринимаем просто как
5.2. Общие соображения о возможности трансвременных причинных связей
5.2. Общие соображения о возможности трансвременных причинных связей Хотя гипотеза формативной причинности предполагает существование трансвременной, или диахронической (то есть характеризующей последовательность развития явлений во времени. — Прим. пер.), причинной
Глава 7. Наследование формы
Глава 7. Наследование формы 7.1. Генетика и наследственность Наследственные различия между организмами, одинаковыми в других отношениях, зависят от генетических различий; эти последние зависят от различий в структуре ДНК или в ее расположении в хромосомах, а эти различия
Глава 7. Формы поведения, аналогичные моральным
Глава 7. Формы поведения, аналогичные моральным Не убий. Пятая заповедь В 5-й главе, где речь шла о процессе ритуализации, я попытался показать, каким образом это явление, причины которого все еще весьма загадочны, создает совершенно новые инстинкты, диктующие организму
ЖИЗНЕННЫЕ ФОРМЫ
ЖИЗНЕННЫЕ ФОРМЫ Пожалуй, любой человек, не обладающий специальными знаниями в ботанике, может дать приблизительное описание большинства мировых биомов: в тропических дождевых лесах много деревьев; в степях и саваннах мало или нет деревьев, но много травы; в пустынях
Половой диморфизм и ведущий тип сексуальных связей: современные приматы и ископаемые гоминины
Половой диморфизм и ведущий тип сексуальных связей: современные приматы и ископаемые гоминины Все же определенные связи между типом сексуальных отношений у обезьян и рядом морфофизиологических параметров мужских и женских особей существуют. В частности,